Effects of anthropogenic infrastructures on the spatial ecology of raptors and bustards
198
0
0
Texto
Imagem

+7
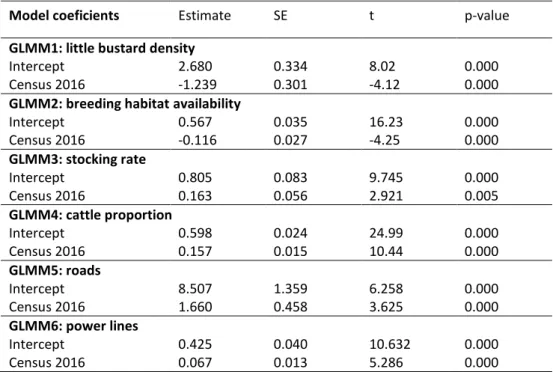
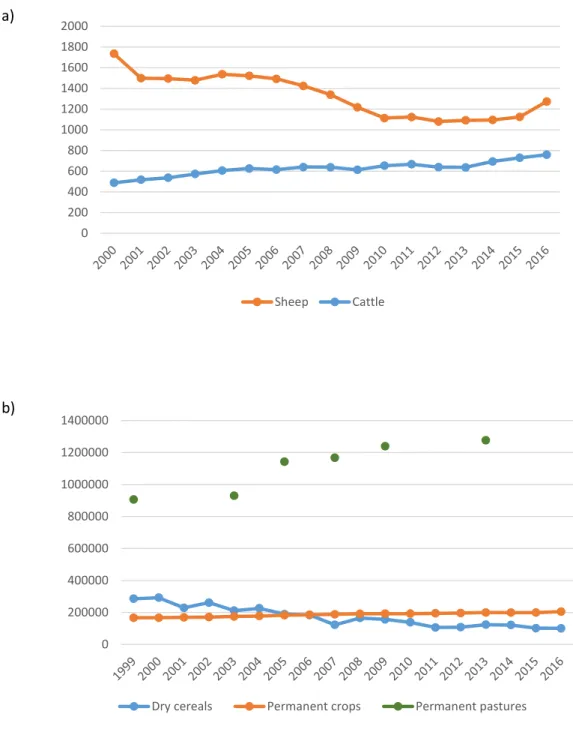
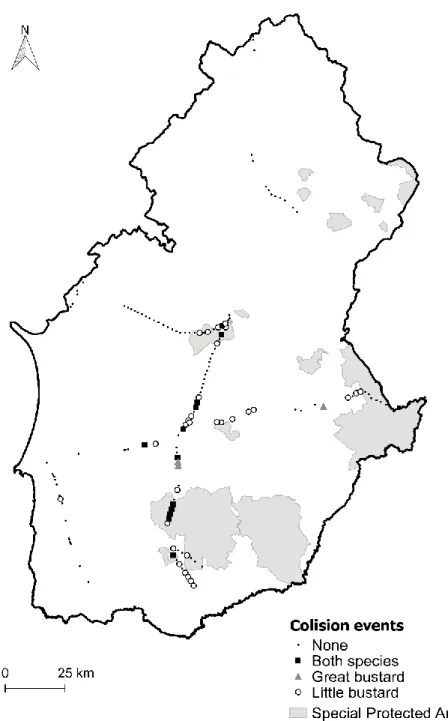
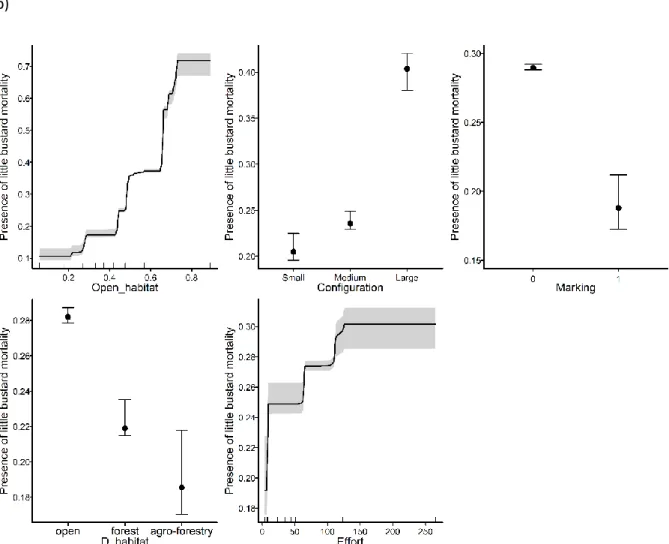
Documentos relacionados