AN EVOKED POTENTIAL MAPPING OF TRANSCALLOSAL
PROJECTIONS IN THE CAT
A. CUKIERT * — C. TIMO-IARIA **
S U M M A R Y — I n t e n a d u l t c a t s a n e s t h e t i z e d w i t h k e t a m i n e h y d r o c h l o r i d e t h e n e o c o r t e x w a s e x p o s e d a n d r e c t a n g u l a r p u l s e s ( 1 m s e c , 0.5 H z a n d v a r i a b l e i n t e n s i t y ) w e r e a p p l i e d t o d i s c r e t e p o i n t s o f o n e s i d e a n d t r a n s c a l l o s a l e v o k e d p o t e n t i a l s w e r e r e c o r d e d f r o m t h e o t h e r . T h e s t i m u l a t i o n a n d r e c o r d i n g p o s i t i o n s w e r e d e t e r m i n e d o n a c a r t e s i a n m a p o f m o s t o f t h e e x p o s a b l e n e o c o r t i c a l a r e a s a n d t h e p o t e n t i a l s w e r e a n a l y s e d a s t o t h e i r c o m p o n e n t s , v o l t a g e a n d l a t e n c y . P a s s i v e s p r e a d a n d e l e c t r o t o n i c p o t e n t i a l s a n d t h e e f f e c t s o f i n c r e a s i n g f r e q u e n c y w e r e a l s o a n a l y s e d . T h e r e s u l t s s h o w e d l a r g e t r a n s c a l l o s a l p o t e n t i a l s i n s o m e a r e a s a n d an i n c r e a s e o f p o t e n t i a l s i n t h e c a u d o r o s t r a l d i r e c t i o n , a t t a i n i n g t h e h i g h e s t v a l u e s i n a n t e r o m e d i a l a r e a s o f t h e s u p r a s y l v i a n g y r u s . C o n f i r m i n g a n a t o m i c a l s t u d i e s , a f e w s i l e n t s p o t s w e r e f o u n d in t h e m o t o r a n d s o m e s t h e t i c c o r t e x a n d i n r e s t r i c t e d p o s t e r i o r r e g i o n s o f t h e v i s u a l c o r t e x , w h e r e s m a l l o r z e r o v o l t a g e s o c c u r r e d . W h i l e c a u s i n g w e a k c o n t r a l a t e r a l p o t e n t i a l s , s t i m u l a t i o n o f s o m e p o s t e r i o r s i t e s p r o v o k e d h i g h v o l t a g e p o t e n t i a l s i n a n t e r i o r r e g i o n s o f t h e s i d e b e i n g s t i m u l a t e d a n d i n t h e c o r r e s p o n d i n g a r e a o f t h e o p p o s i t e s i t e . T h e s e p o s t e r i o r s i t e s are. p o o r l y i n t e r c o n n e c t e d b y t h e c o r p u s c a l l o s u m . T h e L - s h a p e d i n d i r e c t c o n n e c t i o n d e s c r i b e d i n t h i s w o r k m a y b e i n v o l v e d i n s o m e t y p e s o f e p i l e p s y a n d m a y e x p l a i n t h e e f f e c t i v e n e s s o f p a r t i a l c a l l o s o t o m y i n t h e i r t r e a t m e n t .
The corpus callosum, which is the largest brain commissure, plays a key role in interhemispheric integration and as a pathway mediating the spread of secondarily generalized seizures 11,17,19-21,25. Its relevance in pathology of the brain led to several experimental and clinical studies 1,9,16,27,28, Of great significance also have been those related to anatomical distribution and topography of callosal fibers, made in the r a t1 2
,
* L a b o r a t o r y o f F u n c t i o n a l N e u r o s u r g e r y , S c h o o l o f M e d i c i n e U n i v e r s i t y o f S ã o P a u l o , R e s i d e n t i n N e u r o s u r g e r y ; ** L a b o r a t o r y o f C l i n i c a l P h y s i o l o g y , S c h o o l o f M e d i c i n e U n i v e r s i t y o f S ã o P a u l o , F u l l P r o f e s s o r o f P h y s i o l o g y . T h i s r e s e a r c h w a s s u p p o r t e d b y t h e S t a t e o f S ã o P a u l o R e s e a r c h F o u n d a t i o n ( F A P E S P , g r a n t s 84/1078-9 a n d 85/3531-5), F I N E P ( g r a n t 43.86.0222.00) a n d I n s t i t u t e o f t h e L a b o r a t o r i e s o f M e d i c a l R e s e a r c h ( L I M - 7 4 - 4 5 ) .
Laboratório de Fisiologia Clínica, Faculdade de Medicina USP — Caixa Postal 2921 - 01051 São Paulo SP - Brasil
Mapeamento por potencial evocado das projeções transcalosas no gato.
cat 6,8,io,24> monkeys 13-15,23, and humans 26. Transcallosal evoked potentials were
first studied by Curtis 4
- 5 by the time oscillographic recording of E E G and evoked
potentials had been introduced in neurophysiology. He mapped most of the exposable convexity of the cortex in terms of amplitude of the symmetrical contralateral trans-callosal evoked potentials and their topography and time course in a few cortical sites. Morphology and origin of transcallosal evoked potentials were studied much later^.3.
In this paper a detailed analysis of the location of transcallosal evoked potentials in the cat is reported, including a novel type of projection.
M A T E R I A L , A N D M E T H O D S
T e n a d u l t c a t s o f e i t h e r s e x w e r e a n e s t h e t i z e d w i t h k e t a m i n e h y d r o c h l o r i d e a n d ,
a f t e r h a v i n g b e e n f i x e d t o a s t e r e o t a x i c i n s t r u m e n t w e r e s u b j e c t e d t o a n e x t e n s i v e
b i l a t e r a l c r a n i o t o m y t o e x p o s e m o s t o f t h e n e o c o r t e x . O n l y a n a r r o w s a g g i t a l s t r i p o f
b o n e a b o u t t w o m m w i d e w a s left i n t a c t a b o v e t h e l o n g i t u d i n a l s i n u s . F o r l o c a l i z a t i o n
p u r p o s e s a m a p w a s p r e p a r e d a s a C a r t e s i a n m a t r i x m a d e u p o f o n e m i l i m e t e r s q u a r e s ,
w h i c h w e r e d e f i n e d a s s i t e s f o r s t i m u l a t i o n a n d c o n t r a l a t e r a l r e c o r d i n g . W i t h t w o e l e c t r o d e
h o l d e r s t h e s y s t e m w a s s t e r e o t a x i c a l l y c a l i b r a t e d s o t h a t t h e r e a l c o r t i c a l s i t e s w o u l d
m a t c h t h e C a r t e s i a n m a t r i x . T h i s p r o c e d u r e a l l o w e d p l o t t i n g a d e t a i l e d d i s t r i b u t i o n o f
the s t i m u l a t i o n s i t e s a n d o f t h o s e f r o m w h i c h t h e p o t e n t i a l s w e r e r e c o r d e d ( F i g . 1 ) .
A B C O E F G M I K L M N O P O H S
S t i m u l a t i o n o f 138 s i t e s w i t h r e c t a n g u l a r p u l s e s o f v a r i a b l e f r e q u e n c y (0.5 t o 10 H z ) ,
d u r a t i o n (0.1 t o 3 m s ) a n d c u r r e n t (0.5 t o 6 m A ) w a s p e r f o r m e d t h r o u g h b i p o l a r s i l v e r
baU e l e c t r o d e s 0.8 m m in d i a m e t e r , 1 m m a p a r t . F o r m a p p i n g p u r p o s e s o n l y 1 H z , 0.5
m s a n d 4 m A p u l s e s w e r e u s e d . T h e r e c o r d i n g e l e c t r o d e w a s a l s o a s i l v e r b a l l 0.8 m m
in d i a m e t e r . T h e e l e c t r o d e s c o u l d b e p l a c e d , b y m e a n s o f an e l e c t r o d e c a r r i e r , o n w e l l
d e f i n e d s i t e s o f m o s t o f t h e n e o c o r t e x a c c o r d i n g t o t h e C a r t e s i a n m a p . F o r e a c h s t i m u l a t e d
site a n e v o k e d p o t e n t i a l w a s f i r s t r e c o r d e d f r o m t h e h o m o l o g o u s c o n t r a l a t e r a l a r e a . A f t w e r
-w a r d s , p o t e n t i a l s -w e r e r e c o r d e d f r o m s i t e s a l o n g t h e a n t e r o p o s t e r i o r p a r a l l e l s a n d t h e n
f r o m t h e d o r s o v e n t r a l m e r i d i a n s , h a v i n g t h e h o m o l o g o u s l o c u s as its c e n t e r . F o r each
site 20 p o t e n t i a l s w e r e e l e c t r o n i c a l l y a v e r a g e d .
R E S U L T S
M o r p h o l o g y o f t h e t r a n s c a l l o s a l e v o k e d p o t e n t i a l s w a s in g e n e r a l s i m i l a r t o t h o s e
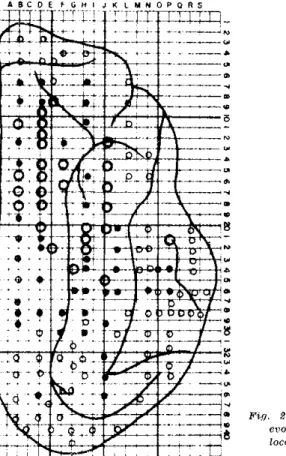
r e p o r t e d b y o t h e r a u t h o r s ( 2 - 5 ) . A s s e e n in F i g . 2 a h i g h a m p l i t u d e z o n e w a s f o u n d
in the m i d d l e a n d a n t e r i o r p a r t s o f t h e c o r t e x , c o m p r i s i n g t h e m a r g i n a l a n d s u p r a s y l v i a n
g y r i , in w h i c h the v o l t a g e d i m i n i s h e d in b o t h a n t e r i o r a n d p o s t e r i o r d i r e c t i o n s . T h e
l o w e s t a m p l i t u d e s w e r e r e c o r d e d f r o m t h e v i s u a l a n d m o r e r o s t r a l c o r t i c a l a r e a s . T h e
A B C D E F G H I J K L M N O P Q R S
Fig. 2 — Relative amplitude of transcallosal evoked potentials as a function of their location on the cortex. °, low voltage;
medium voltage; O , high voltage.
p o t e n t i a l s h a d a n e a r l y a n d a l a t e c o m p o n e n t . T h e f o r m e r , g e n e r a l l y b i p h a s i c o r t r i p h a s i c ,
s t a r t e d after a s h o r t l a t e n c y , r a n g i n g f r o m 2 t o 10 m s a n d s t o o d f r e q u e n c y i n c r e m e n t s
u p t o 50 H z o r m o r e w i t h o u t c h a n g i n g its m o r p h o l o g y . T h e l a t e c o m p o n e n t w a s m o r e
S l i g h t i n c r e a s e s i n f r e q u e n c y , j u s t a b o v e 1 H z , r e d u c e d its a m p l i t u d e ,
a b o l i s h e d b y s t i m u l i d e l i v e r e d at f r e q u e n c i e s b e y o n d 10 H z .
I t w a s v i r t u a l l y
I n w h a t s p a t i a l d i s t r i b u t i o n o f t h e e v o k e d p o t e n t i a l is c o n c e r n e d , t w o p a t t e r n s w e r e
f o u n d . I n o n e , t h e p o t e n t i a l s h a d t h e h i g h e s t v o l t a g e s o v e r o r n e a r t h e c o n t r a l a t e r a l s i t e
h o m o l o g o u s t o t h e s t i m u l a t i o n l o c u s , d e c l i n i n g i n t h e a n t e r i o r , p o s t e r i o r , m e d i a l a n d l a t e r a l
d i r e c t i o n s ( F i g . 3 ) . A s t o t h e o t h e r , s t i m u l a t i o n o f a f e w s i t e s l o c a t e d p o s t e r i o r l y a t t h e
m a r g i n a l a n d s u p r a s y l v i a n g y r i e v o k e d s m a l l r e s p o n s e s f r o m t h e h o m o l o g o u s a r e a s w h i l e
l a r g e p o t e n t i a l s w e r e e l i c i t e d o v e r a n t e r i o r a r e a s , m a i n l y in t h e m e d i o - a n t e r i o r r e g i o n
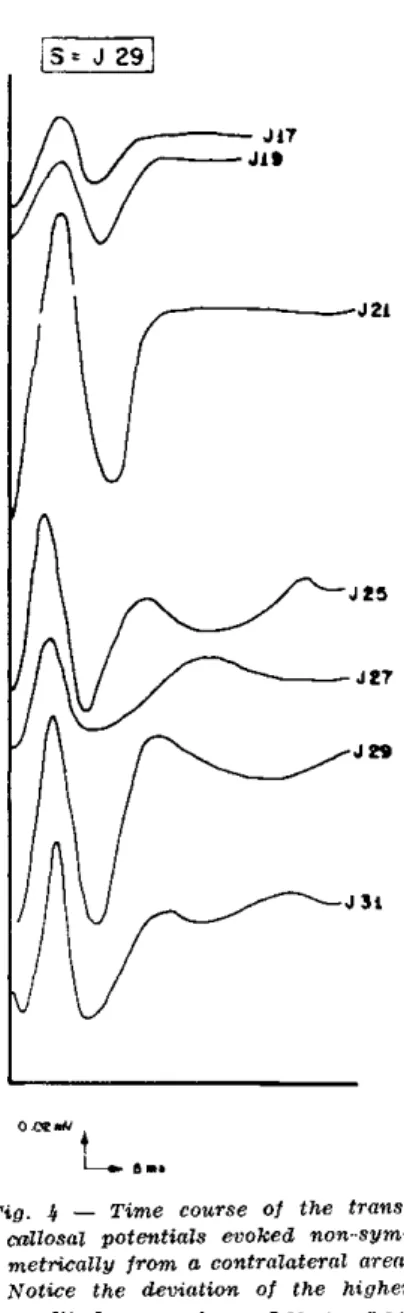
o f t h e m a r g i n a l a n d s u p r a s y l v i a n g y r i ( F i g . 4 ) . T h e s e r e s p o n s e s w e r e e v o k e d f r o m a r e a s
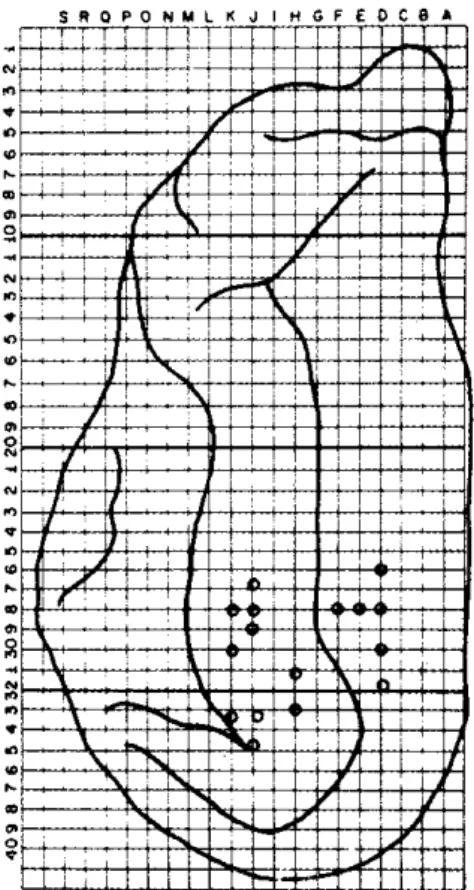
p o o r l y i n t e r c o n n e c t e d b y t h e c o r p u s c a l l o s u m , a c c o r d i n g t o E b n e r a n d M y e r s (<?) m a p ( F i g . 5 ) .
S = H 2 4
—J 21
J 2 5
J 27
J 31
Fig. 3 — Left: transcalloml evoked potentials as a function of position along one of the cortical meri-dians (H, see fig. 1) in response to stimulation of the square H.24- Right: potentials recorded from parallel 2kl notice the increase in voltage and decrease in latency from H.30 and H.19 toward H.26.
S R 0 P 0 N M L K J I H G F E D C B *
Fig. 5 — Areas whose stimulation pro-voked non-symmetrical transcallosal responses.
C O M M E N T S
The above described experiments showed that single pulse stimulation of dis-crete sites of one side of the cerebral cortex in the cat causes complex evoked potentials on the corresponding site of the contralateral hemisphere. The transcallosal evoked potentials are distributed across and along the cerebral cortex essentially in agreement with Curtis 4
. 5 . The high voltage potentials found in some areas are conceivably related to a high density of callosal fibers and/or special intracortical connectivity that recruits a larger number of neurons. The early and late components reflect most probably, the involvement of mono and polisynaptic pathways, respectively, inasmuch as the first showed a marked resistance to increasing frequencies and appeared after short latencies, being always present, in sharp contrast to the longer latency, frequency sensitivity and instability of the late component.
Studying the electrophysiological properties and mechanisms was not a main concern of the present research. Accordingly, the subject will not be discussed at length here. It is noteworthy, however, that the first component of the evoked transcallosal potentials displays some features compatible with the hypothesis that they reflect the activation of axons of the projecting contralateral neurons. This is in partial agreement with the accepted origin of the early sensory evoked potentials as presynaptic in nature. The late component is surely due to the activation of post-synaptic neurons, as seen by its susceptibility to increasing frequencies of stimu-lation and instability, in addition to its longer latency in restimu-lation to the early component.
areas through indirect connections. That might be explained by assuming that the thalamus, receiving information from both the ipsilateral and the contralateral cortex, activates the cortical area far from the symmetric locus. There could be a massive oblique callosal interhemispheric connection between anterior areas of one side and posterior areas of the other. A thalamic-mediated process, if involved, would be difficult to recognize i n our experiments. An oblique direct connection of the corpus callosum cannot either be invoked to explain the non-symmetric callosal response since recent anatomical data have shown it to have not this type of connection 17. The assumption of an intrahemispheric pathway linking the ipsilateral posterior and anterior areas has to be demonstrated by means of callosal and percortical sectioning experi-ments. It is noteworthy also that areas thus interconnected coincide with those poorly interconnected by the corpus callosum, revealing the possibility of non- symmetric callosal activation by means of transcallosal pathways (from one side to the other) in series with intrahemispheric pathways (mainly in the postero- anterior direction). Such a connection would provide the anatomical substrate for integration of several functions which require, for instance, visuomotor interplay.
Severing the corpus callosum has been used as a palliative surgery for polifocal and/or frontal epilepsies non-responding to drugs. Selective anterior and total callosotomy have been advocated 7,9. The present study suggests that, it connections similar to those hereby described do exist in man, some precisely located posterior foci could project anteriorly via this postero-anterior pathways and drive anterior epileptic bursts in polifocal epilepsies. Marcus i 8f when studying the interaction between foci created in both hemispheres at the same time, had already noted in monkeys that parietal foci trigger spike and wave discharges that predominate in more anterior rather than in parietal regions. This fact might be explained by the pathway hereby proposed. Put together all this would suggest that it is not necessary to perform a whole callosotomy to treat drug-intractable epilepsies. Also our data would suggest that seletive anterior callosotomy may interfere with the interhemispheric integration of areas posterior to the limit of the commissurotomy, which may result in disconnection syndromes not expected from the extent of the lesion made.
R E F E R E N C E S
1. A v i l a J O , R a d v a n y J, H u c k F R , F i r e s d e C a m a r g o C H , M a r i n o R J r , R a g a z z o P C , R i v a D — A n t e r i o r c a l l o s o t o m y a s a s u b s t i t u t e f o r h e m i s p h e r e c t o m y . A c t a N e u r o c h i r , s u p p l , 30:137, 1980.
2. C h a n g H T — C o r t i c a l r e s p o n s e t o a c t i v i t y o f c a l l o s a l n e u r o n s . J N e u r o p h y s i o l 16 :117, 1953.
3. C h a n g H T — I n t e r a c t i o n o f e v o k e d c o r t i c a l p o t e n t i a l s . J N e u r o p h y s i o l 16 :133, 1953.
4. C u r t i s H J — I n t e r c o r t i c a l c o n n e c t i o n s uf c o r p u s c a l l o s u m a s i n d i c a t e d b y e v o k e d p o t e n t i a l s . J N e u r o p h y s i o l 3:405, 1940.
5. C u r t i s H J — A n a n a l y s i s o f c o r t i c a l p o t e n t i a l s m e d i a t e d b y t h e c o r p u s c a l l o s u m . J N e u r o p h y s i o l 3:414, 1940.
6. E b n e r F F , M y e r s R E — D i s t r i b u t i o n o f c o r p u s c a l l o s u m a n d a n t e r i o r c o m m i s s u r e in c a t a n d r a c o o n . J C o m p N e u r o l 124:353, 1965.
7. G a t e s J R , M a x w e l l R , L e p p i k I E , G u m n i t R J — E l e c t r o e n c e p h a l o g r a p h i c a n d c l i n i c a l e f f e c t s o f t o t a l c o r p u s c a l l o s o t o m y . I n R e e v e s A G ( e d ) : E p i l e p s y a n d t h e C o r p u s C a l l o s u m . P l e n u m P r e s s , N e w Y o r k , 1965, p g 315.
8. H u b e l D H , W i e s e l T N — C o r t i c a l a n d c a l l o s a l c o n n e c t i o n s c o n c e r n e d w i t h t h e v e r t i c a l m e r i d i a n o f v i s u a l f i e l d s i n t h e c a t . J N e u r o p h y s i o l 30 : 1561, 1967.
9 H u c k F R , R a d v a n y J, A v i l a J O , P i r e s d e C a m a r g o C H , M a r i n o R J r , R a g a z z o P C , R i v a D — A n t e r i o r c a l l o s o t o m y in e p i l e p t i c s w i t h m u l t i f o r m s e i z u r e s a n d b i l a t e r a l s y n c h r o n o u s s p i k e a n d w a v e E E G p a t t e r n . A c t a N e u r o c h i r , s u p p l , 30:127, 1980.
10. I m i g T J , B r u g g e J F — S o u r c e s a n d t e r m i n a t i o n s o f c a l l o s a l a x o n s r e l a t e d t o b i n a u r a l a n d f r e q u e n c y m a p s i n p r i m a r y c o r t e x o f t h e c a t . J C o m p N e u r o l 182:637, 1978.
12. I v y G O K i l l a c k e y H P — T h e o n t o g e n y o f t h e d i s t r i b u t i o n o f c a l l o s a l p r o j e c t i o n n e u r o n s in t h e r a t p a r i e t a l c o r t e x . J C o m p N e u r o l 195:367, 1981.
13. J a c o b s o n S, T r o j a n o w s k y J Q — P r e f r o n t a l g r a n u l a r c o r t e x o f t h e R h e s u s m o n k e y : I I . I n t e r h e m i s p h e r i c c o r t i c a l a f f e r e n t s . B r a i n R e s 132:209, 1977.
14. J o n e s E G , B u r t o n H , P o r t e r R — C o m m i s s u r a l a n d c o r t i c o c o r t i c a l c o l u m n s i n t h e s o m a t i c s e n s o r y c o r t e x o f p r i m a t e s . S c i e n c e 190:572, 1975.
15. J o n e s E G , C o u l t e r J D , W i s e S P — C o m m i s s u r a l c o l u m n s in t h e s e n s o r y m o t o r c o r t e x
o f m o n k e y s . J C o m p N e u r o l 188:133, 1979.
16. L u e s s e n h o p A J — I n t e r h e m i s p h e r i c c o m m i s s u r o t o m y . A m S u r g 36:265, 1970.
17. M a r c u s E M , W a t s o n C W — B i l a t e r a l s y n c h r o n o u s s p i k e a n d w a v e e l e c t r o g r a p h i c p a t t e r n s in t h e c a t . A r c h N e u r o l 14:601, 1966.
18. M a r c u s E M — G e n e r a l i z e d s e i z u r e m o d e l s a n d t h e c o r p u s c a l l o s u m . I n R e e v e s A G ( e d ) : E p i l e p s y a n d t h e C o r p u s C a l l o s u m . P l e n u m P r e s s , N e w Y o r k , 1985, p g 131.
19. M a r e s P — S y m m e t r i c a l e p i l e p t o g e n i c f o c i in c e r e b r a l c o r t e x o f i m m a t u r e rat. E p i l e p s i a 14:422, 1973.
20. M u s g r a v e J, G l o o r P — T h e r o l e o f c o r p u s c a l l o s u m in b i l a t e r a l i n t e r h e m i s p h e r i c s y n c h r o -n y o f s p i k e a -n d w a v e d i s c h a r g e s i-n f e l i -n e g e -n e r a l i z e d p e -n i c i l i -n e p i l e p s y . E p i l e p s i a 21:369, 1980.
21. M u t a n i R , D u r e l l L — M e c h a n i s m s o f i n t e r a c t i o n o f a s y m m e t r i c a l b i l a t e r a l e p i l e p t o g e n i c
f o c i i n n e o c o r t e x . E p i l e p s i a 21:549, 1981.
22. O t t i n o C A , M e g l i o M , R o s s i G F — A n e x p e r i m e n t a l s t u d y o f t h e s t r u c t u r e s m e d i a t i n g b i l a t e r a l s y n c h r o n y o f e p i l e p t i c d i s c h a r g e s o f c o r t i c a l o r i g i n . E p i l e p s i a 12:299, 1971.
23 P a n d y a D N , H e i l b r o w n D — T h e t o p o g r a p h i c d i s t r i b u t i o n o f i n t e r h e m i s p h e r i c p r o j e c t i o n s i n t h e c o r p u s c a l l o s u m o f t h e R h e s u s m o n k e y . B r a i n R e s 32:31, 1971.
24. P o l j a k S — A n e x p e r i m e n t a l s t u d y o f t h e a s s o c i a t i o n c a l l o s a l a n d p r o j e c t i o n f i b e r s o f t h e c e r e b r a l c o r t e x o f t h e c a t . J C o m p N e u r o l 44:197, 1927.
25. S c h w a r t z k r o i n P A , F u t a m a c h i K J , N o e b e l s J L — T r a n s c a l l o s a l e f f e c t s o f a c o r t i c a l e p i l e p t i f o r m f o c u s . B r a i n R e s 55:59, 1975.
26. T o m a s c h J — Size, d i s t r i b u t i o n a n d n u m b e r o f f i b e r s i n t h e h u m a n c o r p u s c a l l o s u m . A n a t R e c 119:119, 1954.
27. V a n W a g e n e n W P , H a r r e n R Y — S u r g i c a l d i v i s i o n o f c o m m i s s u r a l p a t h w a y s i n t h e c o r p u s c a l l o s u m . A r c h N e u r o l P s y c h i a t 44:740, 1940.