Influence of Mother Plant and Seed Coat Colour on
Germination Success
of Retama sphaerocarpa L. (Boissier)
João Pedro Fonseca Ferreira
Thesis to obtain the Master Degree in
Forestry and Natural Resource Management
Supervisor: António Manuel Dorotêa Fabião
Co-supervisor: Marta Reis Carneiro Falcão Afonso
J
ury:President:
Doctor Manuela Rodrigues Branco Simões, Assistant Professor with Aggregation to Instituto Superior de Agronomia, Universidade de Lisboa
Members:
Doctor António Manuel Dorotêa Fabião, Associate Professor with Aggregation to Instituto Superior de Agronomia, Universidade de Lisboa.
Doctor Maria Helena Reis de Noronha Ribeiro de Almeida, Associate Professor, Instituto Superior de Agronomia, Universidade de Lisboa.
I
Acknowledgements
This study has been funded by Fundação para a Ciência e a Tecnologia (FCT) through the Forest Research Center (Pest-OE/AGR/UI0239/2011) and project “Enerwood – Potencial of native woody species for energy production from biomass” (PTDC/AGR-CFL/114826/2009). The author also acknowledges Vera Serrão for the development of the experimental work and Carla Faria for assistance on germination chamber preparation and management.
To my supervisor, Professor António Fabião, and co-supervisor, Dr. Marta Carneiro, I acknowledge the support, availability and comprehension.
To Ana Sofia, my close friend, I thank all the very good moments, good disposal and top tips, and also to Alícia, for all the support and friendship from the beginning of my college career.
To Jota for his senseless but funny stupidity, good disposal and friendship, and to all the other friends, my grateful acknowledgements.
To my family, in special to my Parents, thank you for all the long-lasting support and encouragements.
II
Abstract
The aim of this study was to determine the influence of mother-plant and seed external morphologic characteristics of Retama sphaerocarpa on germination success. Two experiments were established in a germination chamber to evaluate the influence of seed coat colour (black, brown or green) and mother-plant on germination success. A pre-germination treatment of seeds for breaking seed-coat dormancy was performed soaking the seeds in hot water (80ºC) with maintenance in the same water for 24 hours (long scald). For both studies the germination rate and the vigour index were determined. The results after 28 and 162 days suggest that the seed coat colour influenced both evaluated parameters. Germination rate was highest in green-coated seeds (55.1% at 162 days), and the vigour index in brown-coated seeds (26.2). The influence of mother plant on germination rate and vigour index was also confirmed: at the end of the experiment the I5 individual presented the highest values both in germination rate and vigour index with 57.0% and 35.3 respectively, also occurring significant statistical differences between some other individuals. Consequently for seed collecting purposes, the mother-plant selection based on known performances and the seed selection based on seed coat colour may be of utmost importance to optimise seedling production in nursery.
III
Resumo
Influência do Progenitor e da Cor do Tegumento das Sementes no Sucesso Germinativo das Sementes de Retama sphaerocarpa L. (Boissier)
O objectivo do presente estudo consistiu em determinar a influência do progenitor e de características morfológicas externas das sementes de Retama sphaerocarpa no seu sucesso germinativo. Para o efeito instalaram-se numa câmara de germinação dois ensaios para avaliar a influência do progenitor e da cor do tegumento das sementes (preta, castanha, ou verde) na germinação destas. As sementes utilizadas foram submetidas a um tratamento de pré-germinação para quebrar a dormência do tegumento, consistindo numa escarificação com água quente (80ºC) seguida de manutenção das sementes na mesma água previamente à sementeira (escaldão longo). Determinou-se em ambos os ensaios a taxa de germinação e o índice de vigor da mesma. Os resultados após 28 e 162 dias indicaram que a cor do tegumento influenciou ambos os parâmetros. A taxa de germinação foi mais elevada (55,1% após 162 dias) nas sementes de tegumento verde, enquanto o índice de vigor da germinação foi mais alto (26,2) nas de tegumento castanho. Confirmou-se também a influência do progenitor nestes dois parâmetros: no fim do ensaio o progenitor identificado como I5 apresentou os valores mais altos da taxa de germinação e do índice de vigor, respectivamente 57,0% e 35,3, tendo-se também determinado diferenças significativas entre outros progenitores. Consequentemente, para fins de recolha de sementes a selecção de progenitores com performances conhecidas e a selecção de sementes com base na cor do seu tegumento pode ser da maior importância para optimizar a produção em viveiro de plantas desta espécie.
Palavras-chave: Cor do tegumento da semente, Germinação; Progenitor; Retama
sphaerocarpa; Vigor
IV
Resumo Alargado
Influência do Progenitor e da Cor do Tegumento das Sementes no Sucesso Germinativo das Sementes de Retama sphaerocarpa L. (Boissier)
O abandono das terras tem sido uma das mais importantes alterações no uso do solo em áreas rurais da bacia do Mediterrâneo ao longo das últimas décadas. Entre 1961 e 2011, 24,5% da área cultivada com culturas anuais e permanentes no Sul da Europa foram abandonadas (Gabarón-Galeote et al., 2014), atribuindo-se o facto a várias causas sociais e económicas complexas, incluindo as políticas de financiamento da União Europeia para a agricultura e a florestação. Depois do abandono o processo de recuperação começa com a colonização pela vegetação natural e, na Península Ibérica, o piorno-amarelo, ou retama (Retama sphaerocarpa L. (Boissier)), é uma das principais espécies colonizadoras, especialmente em solos delgados de áreas semiáridas e áridas espalhadas, no território de Portugal, pelo vale do rio Douro e por grande parte do interior Centro e Sul do Continente. É um arbusto Mediterrâneo que se inclui taxonomicamente na tribo Genisteae da subfamília Faboideae, família Fabaceae (Boulila et al., 2009). Pode crescer até aos 4 metros de altura, praticamente sem folhas funcionais, substituídas na função por segmentos caulinares fotossintéticos (cladódios) (Guerrouj et al., 2013), que se desenvolvem verticalmente minimizando a interseção da luz e o sobreaquecimento por condução (Haase et al., 2000). Esta espécie de arbusto autóctone tem um elevado interesse para a recuperação de solos degradados, produção de biomassa para energia e utilização em jardins mediterrânicos de baixa manutenção. A retama tem sido também descrita como uma espécie capaz de melhorar as condições microclimáticas ao nível da vegetação de sub-bosque, aumentando a diversidade e produtividade de outras espécies, geralmente herbáceas. Neste contexto, o objetivo deste estudo consistiu em verificar a influência da planta mãe e da cor do tegumento da semente (após o seu humedecimento em tratamento de pré-germinação) no sucesso germinativo das sementes. As hipóteses de trabalho a testar foram (i) que plantas mãe já estudadas e assinaladas como suscetíveis de produzir sementes com elevado sucesso germinativo (Fabião, 2011; Fabião et al., 2014) mantêm essa característica em anos diferentes e (ii) que a cor do tegumento das sementes humedecidas é um bom indicador desse sucesso germinativo.
As sementes do piorno-amarelo foram colhidas manualmente no início de Setembro de 2012 em exemplares desta espécie existentes no campus do Instituto Superior de Agronomia (ISA) (Tapada da Ajuda, Lisboa, 100 ha, 38° 42′ 27.61″ lat. N, 9° 10′ 56.32″ long. O, cota máxima: 131 metros acima do nível do mar), de nove plantas mãe diferentes, já estudadas e localizadas cartograficamente por Fabião (2011) e Fabião et al. (2014). Depois de colhidas, as vagens das sementes foram armazenadas separadamente à temperatura ambiente do Laboratório de Ecologia Florestal do ISA, em envelopes de papel, para evitar contaminação
V externa, até ao começo da experiência. Foram então descascadas manualmente, selecionadas e pesadas, tendo sido excluídas aquelas que mostraram algum tipo de dano. O ensaio de germinação começou a 21 de Novembro de 2012 e durou 162 dias (até dia 2 de Maio de 2013), seguindo as regras recomendadas pela International Seed Testing
Association (ISTA, 2003) para o género Cytisus, por não haver neste normativo
recomendações específicas para o género Retama, numa câmara de germinação localizada no Viveiro Florestal do ISA, mantida durante o ensaio a uma temperatura de 21±3ºC, na ausência de luz.
Foi usado um tratamento de pré-germinação, para quebrar a dormência do tegumento, que consistiu na imersão das sementes em água quente (80ºC), mantendo-as na mesma água por 24 horas (escaldão longo). Seguidamente a este tratamento pré-germinativo, as sementes molhadas mostraram diferentes cores do tegumento – preto (P), castanho (C) e verde (V) – tendo sido separadas por cores e por cada uma das nove plantas mãe (i.e., 3 cores x 9 plantas mãe). Para análise estatística foram consideradas 3 repetições de cada cor e planta mãe, contendo 15 sementes cada (i.e., 3 cores x 9 plantas mãe x 3 repetições que consistiam em 15 sementes).
Os dados da taxa de germinação e do vigor foram organizados e analisados preliminarmente através das plantas progenitoras, por um lado, e das cores do tegumento das sementes, por outro, usando folhas de cálculo e utilitários de produção de gráficos do
software Microsoft Office Excel 2007 SP2 (®Microsoft Corporation, 2007). A análise
estatística foi feita usando o software SPSS 19 (PASW Statistics, Release 19.0.0, 2010; ®SPSS Inc.).
Para a análise estatística detalhada dos resultados, a normalidade da distribuição foi verificada usando o teste de Kolmogorov-Smirnov, e a homogeneidade das variâncias pelo teste de Levene. Quando ambos os pressupostos foram verificados, as comparações entre tratamentos foram realizadas por Análise de Variância (ANOVA) e pelo teste de Tukey. O teste T3 de Dunnett foi usado sempre que se confirmou a normalidade da distribuição, mas não a homogeneidade das variâncias, e testes não paramétricos – teste de Kruskal-Wallis seguido pelo teste de Games-Howell (Maroco, 2010) – para comparar as médias quando ambos os pressupostos da análise de variância não foram satisfeitos. As diferenças entre as cores do tegumento da semente em cada planta mãe não foram testadas, devido ao baixo número de repetições (n=3) disponíveis para tal teste estatístico.
A taxa de germinação teve os resultados mais elevados com as sementes verdes, tendo atingido 55,1% no final do ensaio (162 dias); por outro lado, as sementes pretas apresentaram os resultados mais baixos, tanto para a taxa de germinação, como para o índice de vigor da germinação. As sementes de tegumento castanho tiveram os valores mais elevados do índice de vigor. No que diz respeito à influência da planta mãe na taxa de germinação os melhores indivíduos foram os designados por I2, I5, I6, I7 e I8, com o I5 a
VI atingir o valor mais elevado. Relativamente à influência da planta mãe no índice de vigor da germinação destacaram-se I2, I4, I5, I6 e I7, sendo de novo o I5 o mais elevado.
Estes resultados confirmaram que a cor do tegumento da semente humedecida tem uma boa relação com a taxa de germinação e com o índice de vigor, sendo que as de tegumento verde mostraram os mais elevados valores de vigor médio. Pode constituir, por isso, um bom estimador do sucesso germinativo, segregando como menos interessantes para a propagação em viveiro as sementes que apresentem, após humedecimento, o tegumento de cor preta. Foi também observado que há forte influência do progenitor em ambos, taxa de germinação e índice de vigor, sendo este o fator que mais influenciou estes parâmetros. A taxa de germinação e o índice de vigor tiveram no geral valores superiores no estudo de Fabião et al. (2014) e de Fabião (2011), ao fim de 28 dias e de um período de estudo alargado de cerca de 160 dias, respetivamente, do que no estudo presente. Os resultados relativos à influência da planta mãe não foram totalmente concordantes com os descritos naquelas referências: no estudo presente o grupo dos melhores indivíduos incluiu I2, I4, I5, I6, I7 e I8, enquanto os valores de referência mais elevados foram obtidos das plantas mãe I2, I3 e I7, i.e., apenas 2 deles coincidiram em ambos os estudos, sugerindo todavia uma manutenção no tempo das boas características que já haviam sido assinaladas para estas duas plantas mãe. Estes resultados também sugerem a utilidade de desenvolver linhas de investigação futuras que aprofundem a constância da relação entre o progenitor e o sucesso germinativo das suas sementes, e que esclareçam mais detalhadamente a relação entre a cor do tegumento das sementes e a sua taxa de germinação, designadamente no referente a causas anatómicas e/ou fisiológicas que a fundamentem.
VII
Index
1. Introduction 1.1 General approach... 1
1.2. Rural abandonment in the Mediterranean ... 1
1.3. Establishment and growth of vegetation in Mediterranean disturbed ecosystems ... 2
1.4. Forest biomass utilization in Portugal ... 2
1.5. The Retama sphaerocarpa (L.) Boissier ... 3
2. Material and Methods ... 7
2.1 Germination trials ... 7
2.2. Statistical Treatment ... 10
3. Results
………..……….………….113.1. Seed Color Influence ... 11
3.2. Parental Influence ... 12
4. Discussion ... 18
5. Conclusions ... 20
6. Bibliographic References ... 21
VIII
Figure List
Figure 1 - Retama sphaerocarpa in Tapada da Ajuda, in Lisbon... 4 Figure 2 – Draft of the distribution of Retama sphaerocarpa (L.) Boissier in Portugal (Fabião,
2011). ... 5
Figure 3 - Retama sphaerocarpa pods in Tapada da Ajuda. ... 5
Figure 4 - Map with location of the 9 individuals (I1, I2, I3,…,I9) of Retama sphaerocarpa
evaluated in this trial (adapted from Fabião et al., 2014). ... 7
Figure 5 - Retama sphaerocarpa selected seeds separated by lots of different colours (P –
black, C – brown and V – green). ... 8
Figure 6 - Germination chamber with the 3 trays corresponding to the 3 repetitions (left) and
the tray corresponding to one repetition with 27 lots of seeds (right). ... 8
Figure 7 – Detail of a seed lot containing some germinated seeds and contaminated with
fungal mycelia (left), and of healthy germinated seeds (right). ... 9
Figure 8 - Percentage of germinated seeds after 28 days and at the end of the trial (162
days), grouped by colour. The black bars represent the standard error and the different characters indicate the significant differences (p<0.05). ...11
Figure 9 - Vigour index at 28 days and at the end of the experimental period (162 days),
grouped by seed coat colour. The black bars represent the standard error and the different characters indicate the significant differences (p<0.05). ...12
Figure 10 - Seed germination rates after 28 days and 162 days (at the end of the
experiment) per mother plant (I1, I2, I3, …, I9). The black bars represent the standard error and the different characters indicate the significant differences (p<0.05). ...13
Figure 11 - Vigour index per mother plant at 28 days and at the end of the experimental
period (162 days). The black bars represent the standard error and the different characters indicate the significant differences (p<0.05). ...14
Figure 12 - Percentage of germinated seeds grouped by color (black, brown and green) and
by mother plant (I1, I2, …,I9) in the end of the trial (162 days). The black bars represent the standard error. ...15
Figure 13 - Average Vigour Index by color (Black, Brown and Green) and by individual (I1,
IX
Figure 14 - Germination curves of Retama sphaerocarpa by colour seed lots over the
experiment time-span (162 days). The black bars represent the standard error. ...17
List of abbreviations
ISA – Instituto Superior de Agronomia
ISTA – International Seed Testing Association ANOVA – Analysis of Variance
DNFF – Direcção Nacional das Fileiras Florestais
1
1. Introduction
1.1 General approach
The yellow broom, Retama sphaerocarpa (L.) Boissier, is an autochthonous shrub species in the Iberian Peninsula, with high interest for restoration of degraded soils, biomass production for energy purposes and utilization in low maintenance Mediterranean gardens (Ruiz de la Torre et al., 1996; Barceló and Parera, 2005). This species has been reported as able to improve the microclimatic conditions at the understory vegetation level, increasing the diversity and productivity of herbaceous species (Pugnaire et al. 1996b; Moro et al. 1997a, 1997b). Moro et al. (1997a) refers that this “fertile islands” behave as high biological activity spots dispersed in a heterogeneous landscape, where the dominant interaction between plants is facilitation (Callaway, 1997). This facilitation effect is reciprocal, since the presence of herbaceous species improves the development of adult plants of R. sphaerocarpa, leading Pugnaire et al. (1996a) to describe this relationship as mutualism.
1.2. Rural abandonment in the Mediterranean
Land abandonment has been one of the most important changes on land use on rural areas of the Mediterranean basin over the last decades (Gabarón-Galeote et al., 2014). Between 1961 and 2011, 24.5% of the cultivated area with annual and permanent cultures in Southern Europe (127450 Km2) was abandoned (Gabarón-Galeote et al., 2014). This phenomenon
has been assigned to complex socioeconomic factors such as globalization and specific policy, and the European Union and local governments reduced the financial supports for extensive agriculture and increased those for reforestation of arable land. It has been reported that some ecological properties of abandoned lands are different from those observed in natural areas, as a result of previous crops (Comptonand and Boone, 2000), the colonization by natural vegetation gradually changing soil properties until further reaching of characteristics similar to those observed in undisturbed areas (Bonet, 2004). In Mediterranean environments, secondary successions usually begin by colonization by herbaceous plant communities that persist over a variable time span until the establishment of woody plant dominated communities (Gabarón-Galeote et al., 2014). Nevertheless, the recovery rate depends on the extension of the rain season and how it affects plant cover changes (Poeplau et al., 2011). In wet environments, the colonization of abandoned fields by herb communities causes fast recovery of the soil organic content (Poeplau et al., 2011). On the contrary, in Mediterranean arid and semiarid areas, the soil properties slowly recover after the abandonment due to the seasonality of the grassland cover (Ruiz-Sinoga and Martinez-Murillo, 2009) and to the delay on the colonization process by woody species (Rey Benayas, 2005). Such situation recommends a diversification of vegetation species used in
2 restoration projects, an adequate use of available autochthonous species, and an integrated approach considering the mutual effects on fauna (including microbial), soil and flora (Vallejo, 2009; Fabião et al., 2013).
1.3. Establishment and growth of vegetation in Mediterranean disturbed ecosystems
In the Mediterranean region, agricultural practices over centuries have limited the availability of propagules of some perennial species, influencing the course of secondary succession after land abandonment (Bonet, 2004; Pugnaire et al., 2006). Moreover, in arid ecosystems, plant communities are driven by summer drought and inter-annual and seasonal variations in rainfall (Fabião, 2011). Long and dry summer seasons give herbaceous vegetation a steppe character and explain the dominance of trees and shrubs with perennial leaves, also determining countless xerophytic adaptations (Fabião, 2011). Some Mediterranean species have adaptations to drought: leaf surface reduction, leathery and/or waxed leaves, covered with cuticle on the upper page and/or hair on the lower leaf surface (Fabião, 2011).
In recent decades, the cessation of agriculture in the southwest Spain semiarid region has led to an irregular environment where practically monospecific stands of colonizer species, like Anthyllis cytisoides L. or Retama sphaerocarpa (L.) Boiss. are still part of the landscape several decades after abandonment (Pugnaire et al., 2006). Both species are part of the adult communities and of late succession, but for a long time those communities stayed monospecific. This can be due to limitations in seed dispersal (Pugnaire et al., 2006), arising from distance, obstacles, or seed predation before establishment (Hensen 2002), allelopathy or other soil constraints to seed germination (Pugnaire et al., 2006), weather conditions adverse to plant establishment, inter-specific competition, or herbivory (Pugnaire et al., 2006).
1.4. Utilization of forest biomass for energy in Portugal
In Portugal there is a huge difference between the potential and the effective availability of residues in forests. In many circumstances, only a small part of the residues are economically interesting as raw material for energy production, given orographic conditions of part of the territory, insufficient forest road network, and high costs of extraction and transport of forest residual biomass (DNFF, 2010). Although in short/medium term the utilization of forest biomass will be directed preferably to the thermoelectric power plants (DNFF, 2010), biomass is a raw material susceptible of transformation in several different types of biofuel, both solid (briquettes and pellets), and liquid (ethanol and methanol) or
3 gaseous (methane). Shrub removal for energy can only make sense when associated to the creation or maintenance of corridors or other “open” spaces within a context of wildfire prevention, and in proximity to the utilization centres. Considering the needs of raw material for the supply of the future thermoelectric centrals and the current industrial units that already consume forest biomass, it is expectable that, at a medium term, the pressure over this energetic resource will increase.
The availability of biomass with energy production feasibility is lower than 3 million tons/year, facing an estimated need higher than 4,4 million tons/year (DNFF, 2010). Biomass removal from underexploited sources, through direct actions like pine and eucalypt stump removal, and the use of raw material from stand cleaning and/or eradication of invasive species, is an envisaged approach to minimize biomass deficit (DNFF, 2010). However, establishment of dedicated fast-growing woody species plantations for energy production may also be a possible alternative to guarantee a sustainable offer. Thus, it is of utmost importance to develop studies on biomass utilization from underexploited or unexploited cultures, and on the potential of unmanaged areas, which totalize in Portugal ca. 3 million hectares (ICNF, 2013).
1.5. The Retama sphaerocarpa (L.) Boissier
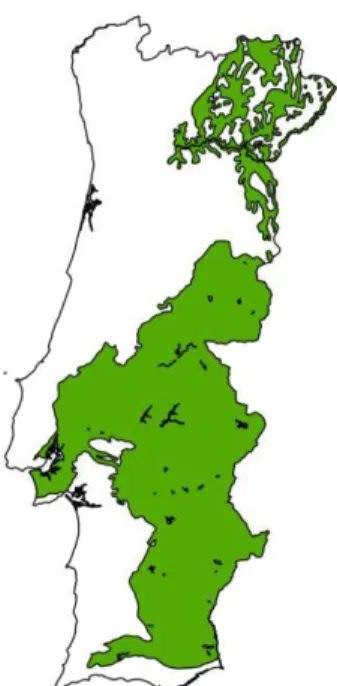
The yellow broom, Retama sphaerocarpa (L.) Boissier, is a Mediterranean shrub belonging to the Genisteae tribe, subfamiliy Faboideae, family Fabaceae (Boulila et al., 2009) (Figure 1). It may grow until 4 meters high, in shrub form, practically without functional leaves, replaced by photosynthetic stems (cladodes) (Guerrouj et al., 2013), which develop vertically minimizing light interception and overheating by conduction (Haase et al., 2000). Therefore, this species has excellent adaptations for water conservation in arid or semiarid ecosystems, allowing it to survive under high radiation and drought conditions. It is very well adapted to the Mediterranean region, growing well in shallow and dry soils until 1400 meters altitude (Ruiz de la Torre et al., 1996; Lopez González, 2004). It occurs naturally in Portugal in the Northeast, mainly within the river Douro basin, and in most of the interior Center and South, usually dominating in successional stages following holm oak stand degradation (Bingre et
al., 2007) (Figure 2).
Yellow broom is a drought tolerant species, forming a symbiosis of the root system with nitrogen fixing bacteria, and possessing a very deep and dense root mat that can reach 25 meters depth (Haase et al., 1996; Fabião, 2011). The root system of Retama sphaerocarpa forms a dense layer of fine roots on the top 20 cm soil layer, corresponding to the most frequent rainfall infiltration depth, and poorly ramified vertical roots that can penetrate deeply
4 Figure 1 - Retama sphaerocarpa in Tapada da Ajuda, in
Lisbon.
into the soil. This dual root system is typical of many phreatophyte species (Mooney & Dun, 1970), allowing water uptake from deep soil strata and opportunistic profit from rain infiltration at the top soil following rain events. R. sphaerocarpa has therefore an important ecological role in arid and semiarid regions where it occurs, contributing to plant species diversity and to ecosystem functions; it is also a valuable species for land rehabilitation and Mediterranean vegetation restoration projects (Caravaca et al., 2003).
The growth season of this species corresponds mainly to a relatively short period of spring, as it frequently happens on other woody species of the Mediterranean semiarid and arid zones (Haase et al., 2000). Nevertheless, blooming occurs between spring and early summer, showing yellow flowers with a green cup; fructification is abundant, usually occurring between July and August and producing brown-yellow pods with one (sometimes two) 3.5-5 mm long ellipsoidal seed (Castroviejo et al., 1999), green-chesty or black when mature (Figure 3). The wind is the main seed dispersal agent, followed by occasional water and animal involvement in the process (Haase et al., 2000; Pugnaire et al., 2006).
5 Figure 3 - Retama sphaerocarpa pods in Tapada da Ajuda.
Figure 2 – Draft of the distribution of Retama sphaerocarpa (L.) Boissier in Portugal (Fabião,
2011).
Previous studies have been contributing to the optimization of nursery production of Retama
sphaerocarpa (Fabião, 2011; Morón Agut, 2012; Fabião et al., 2013, 2014). In this context,
the aims of this study were to coherently verify the mother plant and seed coat colour influence in the germination success of the seeds, following some of the conclusions and recommendations of those studies. For seed coat colour three different seed coat colour
6 were analysed: black, brown and green. The hypothesis of the present study were: (i) the germination rate and vigour index are similar to those observed by Fabião (2011) and Fabião
et al. (2014) in the same mother plants in 2010; (ii) the mother plant influence is similar to
that reported in those studies; and (iii) the seed coat colour has a relationship with germination rate and/or vigour index, as suggested by Morón Agut (2012) and Fabião et al. (2013).
7
2. Material and Methods 2.1 Germination trials
Yellow broom seeds were harvested in the beginning of September 2012 in the Instituto Superior de Agronomia (ISA) campus (100 ha, Lisbon, Portugal, 38° 42′ 27.61″ lat. N, 9° 10′ 56.32″ long. W, max elevation: 131 m a.s.l.), from nine different mother plants (Figure 4). After collecting, seed pods were stored in separated paper envelopes, to avoid external contamination, at laboratory room temperature until the start of the experiment. The seeds were peeled manually, selected and weighted, being excluded those showing any type of damage.
The germination trial started in the 21st November 2012 and lasted for 162 days (until the 2nd
May 2013), following the rules recommended by the International Seed Testing Association (ISTA, 2003) for the Cytisus genus, (there are no recommendations for Retama spp.), in a germination chamber installed at the Forest Nursery of ISA.
Figure 4 - Map with location of the 9 individuals (I1, I2, I3,…,I9) of Retama sphaerocarpa evaluated in
8
Figure 5 - Retama sphaerocarpa selected seeds separated by lots of different colours (P – black,
C – brown and V – green).
Figure 6 - Germination chamber with the 3 trays corresponding to the 3 repetitions (left) and the tray
corresponding to one repetition with 27 lots of seeds (right).
A pre-germination treatment of seeds for breaking seed-coat dormancy was performed soaking the seeds in hot water (80ºC) with maintenance in the same water for 24 hours (long scald), following the recommendations of Morón Agut (2012) after a previous study with seeds from the same origin.
Following the scarification procedure, the wet seeds were sorted by 3 lots of different colours – black (P), brown (C) and green (V) (Figure 5) – for each of the nine different mother trees. Each treatment (seed coat colour) had 3 replicates with 15 seeds each per mother plant (i. e, 3 colours X 9 mother plants X 3 replicates). Overall, a total of 1215 seeds were used.
Seeds were disinfected by immersion in a 1% sodium hypochlorite solution for ca. 10 minutes and thoroughly rinsed with distilled water (Robles et al., 2005). After disinfection, each replicate was randomly distributed over a separate filter paper base, maintained wet in modified Jacobsen’s individual apparatus trays within a germination chamber at 21±3º C, in the absence of light and without humidity control, in a fully randomized design (Figure 6; Appendix I).
9 Figure 7 – Detail of a seed lot containing some germinated seeds and contaminated with fungal
mycelia (left), and of healthy germinated seeds (right).
The experiment was monitored weekly to count the number of germinated seeds and of those showing signs of decay by fungus. The latter were disinfected by a fast immersion in a sodium hypochlorite solution. The chamber environment was maintained wet and, whenever needed, the trays were watered with deionised water. A seed was considered germinated if presenting a radicle extending at least 1 mm to the exterior (López, et al, 1999). Seeds entirely covered by fungus mycelium and with a soft consistency were counted as non-germinated (Figure 7).
The number and percentage of germinated seeds were compiled across seed coat colours and individuals until the 28th day ISTA (2003) (most recommended time span for assessing
the final germination rate), but the end of the experiment was extended until 162 days to allow some stabilization of the germination curves, following a procedure usually recommended whenever germination rate is still increasing clearly after a shorter time-span. Vigour index (V) of each seed lot was determined following the methodology described by López et al. (1999):
V = (a/1 + b/2 + c/3 + d/4 +…+ x/n) x 100/S
where a, b, c… x represent the number of germinated seed after 1, 2, 3… trial days, n is the duration time span (in days) of the experiment and S is the total number of seeds in each replicate. The value of V was classified into ‘null’ (V=0), ‘slow’ (0<V<5.0), ‘medium’ (5.0≤V<11.1), ‘fast’ (11.1≤V<33.3), or ‘very fast’ (33.3≤V≤100), also following López et al. (1999). The proportion of germinated seeds in each replicate was also rated into ‘null’ (0%), ‘low’ (0-29%), ‘moderate’ (30-69%), ‘high’ (70-99%) and ‘maximum’ (100%), adapting a scale proposed by Jurado & Westoby (1992).
10
2.2. Statistical Analysis
Data on seed germination rate and vigour was organized and preliminarily analysed across mother plants and seed coat colours using Microsoft Office Excel 2007 SP2 worksheets (®Microsoft Corporation, 2007). Statistical analysis was performed using the statistics software package SPSS 19 (PASW Statistics, Release 19.0.0, 2010; ®SPSS Inc.), considering a significance level of 5% (α=0.05).
Normality of distribution was verified using the Kolmogorov-Smirnov Test and homogeneity of variances by Levene Test. When both the assumptions were fulfilled, comparisons between treatment means were performed by Analysis of Variance (ANOVA) and Tukey Test. The Dunnett T3 test was used whenever distribution fitted normality, but homogeneity of variance was not equal, even with data transformation. Non-parametric tests, Kruskal-Wallis test followed by the Games-Howell test (Maroco, 2010) were performed to compare means when both assumptions of analysis of variance were not verified.
Differences between seed coat colours in each mother plant were not tested, due to the low number of replicates (n=3) available for such a statistical test.
11
3. Results
3.1. Seed Colour Influence
After 28 days (ISTA, (2003) recommended time span for assessing the final germination rate), germination rate across the different mother plants was significantly higher (p<0.05, Kruskal-Wallis and Games-Howell tests) in brown-coated seeds (24.2%) than in the green- and black-coated (respectively 19.8 and 7.4%, not significantly different) (Figure 8, left). Nevertheless, after 162 days (end of the experiment) those differences increased and become all significant (p<0.05, Kruskal-Wallis and Games-Howell tests), ranking green-coated (55.1%) > brown-green-coated (36.8%) > black-green-coated seeds (11.6%) (Figure 8, right) and therefore an inversion between the germination performances of brown- and green-coated seeds occurred after the increased time period.
Figure 8 - Percentage of germinated seeds after 28 days and at the end of the trial (162 days),
grouped by colour. The black bars represent the standard error and the different characters indicate the significant differences (p<0.05).
The vigour index was also significantly higher (Kruskal-Wallis and Games-Howell tests) after 28 days in brown-coated seeds (10.5, medium) than in black- and green-coated (respectively 2.7 and 3.3, slow), the latter not being statistically different (Figure 9). The increased time-span of 162 days enhanced all the values until 26.2 (brown-coated, fast), 20.7 (green, fast), and 7.5 (black, medium). Differences between the vigour indexes of green- and black-coated seeds become also significant at the end of the experiment (Figure 9).
12 Figure 9 - Vigour index at 28 days and at the end of the experimental period (162 days), grouped by
seed coat colour. The black bars represent the standard error and the different characters indicate the significant differences (p<0.05).
3.2. Mother Plant Influence
Germination rate by mother plant across seed coat colours after 28 days is shown in Figure 10. The mother plants I5, I6 and I7 ranked the highest values (34.8-37.0%, moderate), the first being significantly different (p<0.05, Kruskal-Wallis and Games-Howell tests) from I1, I3 and I9, which ranked the lowest germination rates (3.7-5.9%, low). The extended time period of 162 days enhanced the value of the germination rate of I5 until 57.0% (moderate), followed by I2 (48,9%), I8 (45.9%), I7 (40.0%) and I6 (37.0%), also showing a moderate germination rate. The individuals I3 and I9 showed the lowest germination rates (respectively 14.0 and 16.3%, low), and a statistically significant difference to I5 (Figure 10). Although not significantly different from the others, I1 and I4 also had germination rates below 26%.
13 Figure 10 - Seed germination rates after 28 days and 162 days (at the end of the experiment) per
mother plant (I1, I2, I3, …, I9). The black bars represent the standard error and the different characters indicate the significant differences (p<0.05).
Regarding the vigour index after 28 days (Figure 11), the mother plant I5 was the most noteworthy, exhibiting an index value of 12.1 (fast), significantly higher (p<0.05, Kruskal-Wallis and Games-Howell tests) than those determined for I1 and I9 (respectively 1.9 and 1.5, slow). Mother plants I3 and I6 also had a slow vigour index, whereas I2, I4, I7 and I8 were medium, but with no significant differences. As observed for the germination rate, the extension of the experiment until 162 days enhanced the values maintaining most of the differences between the different mother plants (Figure 11): I5 maintained the highest vigour
14 Figure 11 - Vigour index per mother plant at 28 days and at the end of the experimental period
(162 days). The black bars represent the standard error and the different characters indicate the significant differences (p<0.05).
index (35.3, very fast), significantly different from those of I3 and I9 (7.9 and 6.0, medium). Although not significantly different from the remaining, I2, I4, I6, I7 and I8 also had a very fast vigour index, and I1 only increased the vigour until a medium level.
15 Figure 12 - Percentage of germinated seeds grouped by color (black, brown and green) and by
mother plant (I1, I2, …,I9) in the end of the trial (162 days). The black bars represent the standard error.
Details on germination by mother plant and seed coat colour at the end of the experiment (162 days) are shown in Figure 12. Green-coated seeds usually had the highest germination percentages, reaching 88.8% in I8 and 86.7% in I7. These mother plants and I1, I6 and I9 attained the highest germination rates in green-coated seeds, the latter with values only rated as ‘moderate’. Green-coated seeds obtained from mother plant I3 had a low germination rate (15.6%), but this individual had the highest germination rate from brown-coated seeds. Brown-brown-coated seeds also ranked the highest germination rates in I2, I4 and I5, reaching 80.0% in the former and 77.8% in the latter, whereas I1 showed null germination for these seeds. Black-coated seeds almost always had the lowest germination percentages (except for mother plant I1 and I9, ranking higher than brown-coated), attaining the highest value in mother plant I5, with 24.4%, followed by 17.8% in I1 (both still moderate).
Vigour index followed a different trend of germination rate, with highest values in brown-coated seeds (Figure 13), and 6 mother plants having the maximum vigour in this type of seeds. Mother plants I2 and I5 ranked ‘very fast’ for brown seeds, attaining respectively 47.4 and 61.3, and I3, I4, I6 and I8 had vigour index values within the range 18.3-29.4, still ranking as fast. The remaining 3 mother plants (I1, I7 and I9) showed the highest vigour index in green-coated seeds: I7 with 53.6 (very fast), whereas the other two ranked fast (I1) to medium (I9). Black-coated seeds had a generalized slow to medium vigour, with the exception of mother plants I4 and I5, which ranked ‘fast’.
16
Figure 13 - Average Vigour Index by color (Black, Brown and Green) and by individual (I1, I2,…,I9)
at the end of the trial (162 days). The black bars represent the standard error.
Figure 14 shows the germination curves for green-, brown- and black-coated seeds across mother plants. Green-coated seeds attained a germination rate higher than the others after ca. 75 days, but they were slower in the germination process than the brown-coated ones, which exhibited the highest germination rates until the 40th day after the establishment of the
experiment. During a short period of ca 15 days at the beginning of the experiment, even the black-coated seeds germinated faster than the green-coated ones. Nevertheless, after ca 90 days both black- and brown-coated seeds tended to have their germination stabilized, whereas green-coated seeds were still increasing.
17 Figure 14 - Germination curves of Retama sphaerocarpa by colour seed lots over the experiment time-span (162 days). The black bars represent the standard error.
18
4. Discussion
Germination rate across individuals after 28 days was significantly higher in brown-coated seeds (24.2%) than in the other, which suggest that this colour type responds faster when placed into germination. Nevertheless, after 162 days, the highest germination rate was observed in green-coated seeds (55.1%); an hypothesis to explain this results can be the fact that they are the more recently formed seeds in the mother plants of Retama
sphaerocarpa. By the other hand the black-coated seeds presented the worst results relative
to germination (11.6%). As mentioned an inversion between the germination performances of brown- and green-coated seeds after the increased time period was observed suggesting that brown-coated seeds respond faster to germination conditions over green-coated seeds although after a large time span green-coated had the best performance.
The vigour index determination may be very important from a practical point of view, because by providing additional information to the germination percentages it may contribute to estimate more accurately the success of plants after establishment under field conditions (López et al., 1999).
The obtained values of the vigour index after 28 and 162 days suggest that brown- and green-coated seeds may represent different germination strategies of seeds from the same mother plants, the former reacting more rapidly to favourable germinating conditions than the latter, but approaching to similar germination rates and vigour index values after a more extended time-span. A possible explanation for this difference, as suggested by Fabião et al. (2014) in a slightly different context, may arise from a slight delay in seed maturation status of green-coated seeds when compared to brown-coated ones, or from differences in seed coat resistance to water absorption (seed coat dormancy, or hardness), even after scarification with hot water. Seed coat thickness in legumes may considerably vary (Piotto & Ciccarese 1999). As a consequence, the treatments applied in this study might provoke an unwanted selection, destroying seeds with lighter coats and simultaneously not scarifying seeds with thicker coats (Piotto & Ciccarese 1999). As reported by these authors, mechanical scarification may be a safer and less damaging alternative to chemical or thermal treatments.
Nevertheless, this does not clarify why black-coated seeds exhibited the lowest values of germination rate and vigour index, suggesting that further research must be addressed to seed maturation seasons, seed coat structure and thickness, and physiological processes involved in seed maturation. Retama shpaerocarpa shrubs tend to keep some seed pods for more than one year (Haase et al., 2000), Such seeds may be more prone to fungal contamination, reducing their germination capacity, which seems to have been confirmed in our experiment by the number of removed seeds all over the germination survey after fungal contamination: 357 black coated, 254 brown and 112 green.
19 Our results also confirmed a strong influence of mother plants in seed germination performances, with mother plants identified as I5, I6 and I7 ranking the highest germination rates (34.8-37.0%) after 28 days. Only I7 is coincident with the mother plants identified by Fabião et al. (2014) as having the highest seed germination rates after the same germination time-span, the others being I2 and I3 instead (47.2-52.2%). After the extended germination time period of 162 days the mother plants I2 and I8 attained the highest germination rates (37.0-57.0%) together with the above mentioned I5-I7, but still maintaining some differences relative to the results obtained by Fabião (2011) after a germination time period also extended to achieve the stabilization of the germination curves. This suggests an environmental mediation of the genetic aptitudes of the observed mother plants, nevertheless confirming that I2 and I7 maintained a good performance in both studies.
This experiment also confirmed a high vigour index in the germination of seeds obtained from mother plant I2 after 28 days, as reported by Fabião et al. (2014). Nevertheless, in our experiment the vigour index values of the mother plants with the best performances (I2, I4, I5, I7 and I8) only ranked 5.4-12.1, which was clearly lower than the 6.0-24.5 ranked by all the mother plants (I1-I9) in that study. After the extended time period of 162 days our results become closer to those reported by Fabião (2011) for a similarly extended time-span: 15.6-35.3 for the best mother plants (I2 and I4-I7) in the present experiment, to compare to 36.8 (very fast) in I2 and a fast vigour in all the other mother plants, whereas in this study I1, I3 and I9 maintained a medium vigour index after 162 days.
Most of the highest germination rates and vigour index results after 162 days (I2 and I5-I8) were clearly due to brown- (I2 and I5) and green-coated seeds (I7 and I8), with I6 showing the best germination rate in green-coated seeds and the best vigour index in brown-coated ones. Black-coated seeds only had a marginal contribution for seed germination performances in all the mother plants, only occasionally reaching 20% of germination rate (I5), or 10.0 (medium) in vigour index (I4 and I5).
The overall obtained results could be expected better than observed, considering that many Fabaceae species have seed physical dormancy braked more efficiently by mechanical scarification than by hot water or chemical methods, as reported by Baskin and Baskin (2001). Mondoni et al. (2013) mentions that the use of percussion to break seed physical dormancy has been proposed as a further alternative, successfully tested on several herbaceous species of the Fabaceae family. However, mechanical scarification of large amounts of seed (as those needed for commercial seedling production in nursery) may require specific and highly expensive machinery (Fabião et al., 2014).
20
5. Conclusions
The results obtained in this study only partially confirmed the performances reported for seeds obtained from the same mother plants in a previous experiment (Fabião, 2011; Fabião
et al., 2014), suggesting that environmental mediation may also influence the mother plant
performance for seed germination rate and vigour index. Nevertheless, two mother plants, I2 and I7, consistently maintained a good performance in seed germination rate and vigour index in both studies.
The hypothesis of a relationship between the wetted seed-coat colour and germination performance was confirmed by significant differences in germination rate and vigour index between black-, brown- and green-coated seeds after 28 and 162 days of germination time period, showing a faster response to favourable germination condition by brown-coated seeds, followed by the green-coated ones, whereas black-coated seeds had modest germination rate and vigour index, with significantly lower values after 162 days. The results showed that the green-coated seeds reached a germination rate significantly higher than the others after 162 days, followed by the brown-coated ones. Further research will be needed to clarify the relationship between the wetted seed-coated colour and the structural and physiological seed properties that induce these different germination behaviours.
21
6. Bibliographic References
Barceló, M., Parera, J. 2005. Jardinería e escasez de agua. Bricojardinería y Paisajismo 133: 24-27.
Baskin, C., Baskin, J. 2001. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. Academic Press. Kentucky. Pp 333-337.
Bingre, P., Aguiar, C., Espírito-Santo, D., Arsénio, P. & Monteiro-Henriques, T. (Coords.), 2007. Guia de Campo – as Árvores e Arbustos de Portugal Continental. In: J. Sande-Silva (Coord. ed.). Colecção Árvores e Florestas de Portugal, Volume. 9, Jornal Público / Fundação Luso-Americana para o Desenvolvimento / Liga para a Protecção da Natureza. Lisboa.
Bonet, A., 2004. Secondary succession of semi-arid Mediterranean old-fields in southeastern Spain: insights for conservation and restoration of degraded lands. J. Arid Environ. 56: 213– 233.
Boulila, F., Depret, G., Boulila, A., Belhadi, D., Benallaoua, S., Laguerre, G. 2009. Retama species growing in different ecological-climatic areas of northeastern algeria have a narrow range of rhizobia that form a novel phylogenetic clade within the Bradyrhizobium genus.
Syst. Appl. Microbiol. 32: 531–532.
Callaway, R.M. 1997. Positive interactions in plant communities and the individualistic continuum concept. Oecologia 112: 143–149.
Caravaca, F., Alguacil, M.M., Figueroa, D., Barea, J.M., Roldán, A. 2003. Re-establishment of Retama sphaerocarpa as a target species for reclamation of soil physical and biological properties in a semi-arid Mediterranean area. Forest Ecol. Manag. 182: 49–58.
Castroviejo, S., Aedo, C., Romro-Zarco, C., Sáez, L., Salgueiro, F.J., Talavera, S., Velayos, M. (Eds). 1999. Flora Ibérica. Plantas Vasculares de la Peninsula Iberica e Islas
Baleares: Vol. VII (I), Leguminosae (partim). Real Jardín Botánico, Servicio de
Publicaciones del C.S.I.C., Madrid. Pp. 137-138.
Comptonand, J.E., Boone, R.D., 2000. Long-term impacts of agriculture on soil carbon and nitrogen in New England forests. Ecology 81: 2314–2330.
22 DNFF. 2010. Culturas Energéticas Florestais – Primeira Abordagem do Levantamento da
Situação Actual. Direcção Nacional das Fileiras Florestais. Lisboa
Fabião, André M. D. 2011. Contributo para a Optimização da Produção em Viveiro de
Retama sphaerocarpa (L.) Boissier. Dissertação para a obtenção do Grau de Mestre em
Gestão e Conservação de Recursos Naturais. Instituto Superior de Agronomia / Universidade de Évora. Lisboa.
Fabião, André M. D., Morón Agut, R., Faria, C., Moreira, R., Carneiro, M., Almeida, M. H., Fabião, A. 2013. Contribuição para a optimização da produção em viveiro de Retama
sphaerocarpa (L.) Boissier. In: J. Bento, J. Lousada, M. S. Patrício (Eds.). 7 Congresso Florestal Nacional - Artigos / Comunicações. Sociedade Portuguesa de Ciências Florestais,
Vila Real e Bragança, Pp 35-45 (Edição em CD-ROM).
Fabião, André, Faria, C., Almeida, M.H., Fabião, António. 2014. Influence of mother plant and scarification agents on seed germination rate and vigor in Retama sphaerocarpa L. (Boissier). iForest – Biogeosciences and Forestry 7, 306-312.
Gabarón-Galeote, M. A., Trigalet, S., van Wesemael, B. 2015. Soil organic carbon evolution after land abandonment along a precipitation gradient in southern Spain. Agriculture,
Ecosystems and Environment 199: 114-123.
Guerrouj, K., Ruiz-Diez, B., Chahboune, R., Ramirez-Bahena, M.-H., Abdelmoumen,H., Quinones, M.A., Missbah El Idrissi, M., Velazquez, E., Fernandez-Pascual, M.,Bedmar, E.J., Peix, A. 2013. Definition of a novel symbiovar (sv. retamae) within Bradyrhizobium retama sp. nov.: nodulating Retama sphaerocarpa and Retama monosperma. Syst. Appl. Microbiol. 36: 218–223.
Haase, P., Pugnaire, F.I., Fernandez, E.M., Puigdefabregas, J., Clark, S.C., Incoll, L.D. 1996. An investigation of rooting depth of the semiarid shrub Retama sphaerocarpa (L) Boiss by labelling of ground water with a chemical tracer. J.Hydrol. 177: 23–31.
Haase, P., Pugnaire, F., Clark S., Incoll, L. 2000. Dynamics of cohorts of cladodes and related effects on reproduction in the shrub Retama sphaerocarpa in semi-arid south-eastern Spain. Plant Ecology 146: 105-115.
Hensen, I. 2002. Seed predation by ants in south-eastern Spain (Desierto de Tabernas, Almería). Anales de Biología 24: 89–96.
23 ICNF. 2013. IFN6 – Áreas dos Usos do Solo e das Espécies Florestais de Portugal
Continental. Resultados preliminares. Instituto da Conservação da Natureza e das Florestas.
Lisboa.
ISTA. 2003. International Rules for Seed Testing. International Seed Testing Association, Bassersdorf. Switzerland.
Jurado E. & Westoby M. 1992. Germination biology of selected central Australian plants.
Austral. J. Ecol. 17: 341-348.
Lopez González, G. 2004. Guia de los Árboles y Arbustos de la Península Ibérica y Baleares (2ª Edición). Ediciones Mundi-Prensa. Madrid.
López, J., Devesa, J.A., Ruiz, T., Ortega-Olivencia, A. 1999. Seed germination in Genisteae (Fabaceae) from South-West Spain. Phyton 39: 107-129.
Maroco, J., 2010. Análise Estatística com Utilização do SPSS (3ª Edição). Edições Sílabo. Lisboa.
Mondoni, A., Tazzari, E.R., Zubani, L., Orsenigo, S., Rossi, G. 2013. Percussion as an effective seed treatment for herbaceous legumes (Fabaceae): implications for habitat restoration and agriculture. Seed Science and Technology 41: 175-187.
Mooney, H. A & Dunn, E. O. 1970. Convergente evolution of Mediterranean sclerophyll shrubs. Evolution 24: 292-303.
Moro, M.J., Pugnaire, F.I., Haase, P., Puigdefábregas, J. 1997a. Effect of the canopy of
Retama sphaerocarpa on its understory in a semiarid environment. Functional Ecology 11:
425–431.
Moro, M.J., Pugnaire, F.I., Haase, P., Puigdefábregas, J. 1997b. Mechanisms of interaction between a leguminous shrub and its understory in a semi-arid environment. Ecography 20: 175–184.
Maroco, J., 2010. Análise Estatística com Utilização do SPSS (3ª Edição). Edições Sílabo. Lisboa.
24 Morón Agut, Rosa. 2012. Métodos de Tratamento de Pré-Germinação da Semente em
Leguminosas para Optimização da Produção de Plantas. Instituto Superior de Agronomia,
Lisboa / Universidad de Córdoba, Córdoba.
Piotto B., Ciccarese L. 1999. Storage of scarified seeds of shrubs and trees in the legume family. Canadian Tree Improvement Association News Bulletin 30: 6-7.
Poeplau, C., Don, A., Vesterdal, L., Leifeld, J., Van Wesemael, B., Schumacher, J., Gensior, A. 2011. Temporal dynamics of soil organic carbon after land-use change in the temperate zone — carbon response functions as a model approach. Glob. Chang. Biol. 17: 2415–2427.
Pugnaire, F.I., Haase, P., Puigdefábregas, J., 1996a. Facilitation between higher plant species in a semiarid environment. Ecology 77: 1420–1426.
Pugnaire,F.I., Haase, P., Puigdefábregas, J., Cueto M., Clark, S.C., Incoll, D. 1996b. Facilitation and succession under the canopy of a leguminous shrub, Retama sphaerocarpa, in a semiarid environment in south-east Spain. Oikos 76: 455–464.
Pugnaire, F.I., Luque, M.T., Armas, C., Gutiérrez, L. 2006. Colonization processes in semi-arid Mediterranean old-fields. Journal of Arid Environments 65: 591-603.
Rey Benayas, J.M. 2005. Restoration after land abandonment. In: Mansourian, S., Vallauri, D., Dudley, N. (Eds.), Forests Restoration in Landscapes: Beyond Planting Trees. Springer, New York, pp. 356–360.
Robles, A.B., Castro, J., González-Miras, E., Ramos, M.E. 2005. Effects of Ruminal Incubation and Goats Ingestion on Seed Germination of Two Legume Shrubs: Adenocarpus
decorticans Boiss. and Retama sphaerocarpa (L.) Boiss. In: Alcaide, E.M., Ben Salem, H.,
Biala, K., Morand-Fehr, P. (Eds), Sustainable Grazing, Nutritional Utilization and Quality of
Sheep and Goat Product – Proceedings of the First Joint Seminar of the FAO-CIHEAM Sheep and Goat Nutrition and Mountain and Mediterranean Pasture Sub-Networks.
CIHEAM/FAO/CSIC, Zaragoza, pp. 111-115.
Ruiz De La Torre, J., Carreras, C., García Viñas, I., Orti, M. 1996. Manual de la Flora
para la Restauración de Áreas Críticas y Diversificación en Masas Forestales . Junta de
Andalucía. Consejería de Medio Ambiente.
25 variability to the 2004–06 drought along a climatic gradient of the Littoral Betic Range (southern Spain). Geomorphology 103: 351–362.
Vallejo, V. 2009. Problems and Perspectives of Dryland Restoration. In: Bautista, S., Aronson, J., Vallejo, V. (Eds), Land Restoration to Combat Desertification: Innovative
Approaches, Quality Control and Project Evaluation. Fundación Centro de Estudios
26
27
Attachment I
Experimental design used in germination chamber for evaluation of germination and vigour capacity for the 9 individuals (1, 2, 3, …, 9) and 3 colours (P – black, C – brown and V – green). Each repetition corresponds to one tray in the germination chamber.
Repetition 1
7P 3V 7V 5V 4C
4V 2C 3P 8V 5C
2V 1C 7C 1P 3C
8C 6V 4P 9P 6C
2P 8P 6P 5P 1V
9C 9V
Repetition 2
2V 7C 8C 9C 8V
3P 5P 6P 8P 5V
7P 3V 4V 4C 2P
6C 2C 1C 1V 5C
3C 9V 7V 1P 6V
9P 4P
Repetition 3
9P 3C 2C 5P 3P
8V 9C 8C 2V 7C
7P 3V 4V 4C 2P
6C 7V 9V 5C 1V
6V 5V 6P 4P 8P
1P 1C
28
Attachment II
Germination (%) and vigour index for each seed colour (black, brown and greens) at the end of the 28 days (ISTA) and at the trial end (162 days) with indication of the existence or net of significant differences, with a significance level of 5%.
Germination (%) Vigour
28 days 162 days 28 days 162 days
Black
7,4 ± 1,6 a 11,6 ± 1,9 a 2,7 ± 0,6 a 7,5 ± 1,4 a
Brown 24,2 ± 4,3 b 36,8 ± 6,0 b 10,5 ± 2,1 b 26,2 ±4,5 b
Green 19,8 ± 4,5 ab 55,1 ± 5,6 c 3,3 ± 0,8 ab 20,7 ± 3,2 b
Attachment III
Germination (%) and Vigour Index for each one of the nine individuals (I1, I2, I3, …, I9) at the end of the 28 days (ISTA) and at the trial end (162 days), with indication of existence or not of significant differences, with a significance level of 5%.
Germination (%) Vigour
28 days 162 days 28 days 162 days
I1 3,7 ± 3,2 a 25,2 ± 9,8 abc 1,9 ± 0,8 a 9,5 ± 3,7 abc I2 17,8 ± 6,7 ab 48,9 ± 11,4 abc 6,3 ± 3,5 ab 22,6 ± 7,5 abc I3 5,2 ± 3,1 a 14,1 ± 4,8 b 2,9 ± 1,9 ab 7,9 ± 3,7 b I4 14,1 ± 3,2 ab 25,9 ± 5,6 abc 5,4 ± 1,7 ab 15,6 ± 3,6 abc I5 36,3 ± 7,8 b 57,0 ± 8,5 c 12,1 ± 4,4 b 35,3 ± 7,9 c I6 37,0 ± 4,8 ab 37,0 ± 9,5 abc 3,9 ± 1,5 ab 16,1 ± 4,1 abc I7 34,8 ± 11,2 ab 40,0 ± 13,4 abc 8,9 ± 2,8 ab 27,9 ± 8,6 abc I8 17,8 ± 6,2 ab 45,9 ± 13,1 abc 6,6 ± 2,4 ab 22,3 ± 5,7 abc I9 5,9 ± 2,8 a 16,3 ± 5,6 ab 1,5 ± 0,8 a 6,0 ± 2,3 ab
29
Attachment IV
Germination at 28 days. Statistics results with the Games-Howell test to ascertain between what individuals existed statistical significant differences.
Games-Howell
Germination 28 days Mean Difference (I-J) Std. Error Sig.
I1 vs I2 -12,389 10,552 0,951 I1 vs I3 4,611 9,074 1,000 I1 vs I4 -14,889 8,261 0,681 I1 vs I5 -33,389 8,368 0,024 I1 vs I6 -11,778 10,227 0,956 I1 vs I7 -22,056 12,105 0,670 I1 vs I8 -15,833 9,263 0,734 I1 vs I9 2,222 8,910 1,000 I2 vs I3 17,000 10,264 0,762 I2 vs I4 -2,500 9,553 1,000 I2 vs I5 -21,000 9,645 0,468 I2 vs I6 0,611 11,296 1,000 I2 vs I7 -0,967 13,021 0,997 I2 vs I8 -3,444 10,432 1,000 I2 vs I9 14,611 10,119 0,863 I3 vs I4 -19,500 7,890 0,316 I3 vs I5 -38,000 8,001 0,006 I3 vs I6 -16,389 9,930 0,765 I3 vs I7 -26,667 11,855 0,430 I3 vs I8 -20,444 8,933 0,401 I3 vs I9 -2,389 8,566 1,000 I4 vs I5 -18,500 7,066 0,252 I4 vs I6 3,111 9,193 1,000 I4 vs I7 -7,167 11,245 0,999 I4 vs I8 -0,944 8,107 1,000 I4 vs I9 17,111 7,701 0,438 I5 vs I6 21,611 9,289 0,389 I5 vs I7 11,333 11,323 0,978 I5 vs I8 17,555 8,215 0,486 I5 vs I9 35,611 7,815 0,008 I6 vs I7 -10,278 12,759 0,995 I6 vs I8 -4,056 10,103 1,000 I6 vs I9 14,000 9,779 0,869 I7 vs I8 6,222 12,000 1,000 I7 vs I9 24,278 11,730 0,528 I8 vs I9 18,056 8,767 0,529
30
Attachment V
Vigour at 28 days. Statistics results with the Games-Howell test to ascertain between what individuals existed statistical significant differences.
Games-Howell
Vigour 28 days Mean Difference (I-J) Std. Error Sig.
I1 vs I2 -8,778 10,625 0,994 I1 vs I3 1,889 9,994 1,000 I1 vs I4 -19,722 8,587 0,398 I1 vs I5 -30,000 8,386 0,050 I1 vs I6 -9,889 9,928 0,981 I1 vs I7 -18,889 11,584 0,775 I1 vs I8 -20,222 9,280 0,461 I1 vs I9 2,111 8,970 1,000 I2 vs I3 10,667 11,363 0,987 I2 vs I4 -10,944 10,147 0,968 I2 vs I5 -21,222 9,978 0,495 I2 vs I6 -1,111 11,305 1,000 I2 vs I7 -10,111 12,784 0,996 I2 vs I8 -11,444 10,746 0,971 I2 vs I9 10,889 10,474 0,975 I3 vs I4 -21,611 9,484 0,410 I3 vs I5 -31,889 9,303 0,069 I3 vs I6 -11,778 10,715 ,966 I3 vs I7 -20,778 12,265 0,742 I3 vs I8 -22,111 10,123 0,458 I3 vs I9 0,222 9,834 1,000 I4 vs I5 -10,278 7,772 0,910 I4 vs I6 9,833 9,416 0,974 I4 vs I7 0,833 11,148 1,000 I4 vs I8 -0,500 8,736 1,000 I4 vs I9 21,833 8,399 0,260 I5 vs I6 20,111 9,233 0,465 I5 vs I7 11,111 10,994 0,978 I5 vs I8 9,778 8,539 0,957 I5 vs I9 32,111 8,195 0,026 I6 vs I7 -9,000 12,211 0,997 I6 vs I8 -10,333 10,058 9,977 I6 vs I9 12,000 9,767 0,938 I7 vs I8 -1,333 11,695 1,000 I7 vs I9 21,000 11,446 0,662 I8 vs I9 22,333 9,114 0,323
31
Attachment VI
Germination at 162 days. Statistics results with the Games-Howell test to ascertain between what individuals existed statistical significant differences.
Games-Howell
Germination 162 days Mean Difference (I-J) Std. Error Sig.
I1 vs I2 -18,222 11,771 0,818 I1 vs I3 6,444 9,668 0,999 I1 vs I4 -5,056 9,491 1,000 I1 vs I5 -26,056 9,724 0,240 I1 vs I6 -11,333 10,961 0,976 I1 vs I7 -9,388 13,058 0,998 I1 vs I8 -15,056 12,487 0,944 I1 vs I9 4,167 9,898 1,000 I2 vs I3 24,667 9,903 0,316 I2 vs I4 13,167 9,731 0,886 I2 vs I5 -7,833 9,958 0,995 I2 vs I6 6,889 11,169 0,999 I2 vs I7 8,833 13,233 0,999 I2 vs I8 3,167 12,670 1,000 I2 vs I9 22,389 10,128 0,449 I3 vs I4 -11,500 7,043 0,775 I3 vs I5 -32,500 7,354 0,010 I3 vs I6 -17,778 8,926 0,570 I3 vs I7 -15,833 11,403 0,882 I3 vs I8 -21,500 10,745 0,568 I3 vs I9 -2,278 7,582 1,000 I4 vs I5 -21,000 7,120 0,148 I4 vs I6 -6,278 8,734 0,998 I4 vs I7 -4,333 11,253 1,000 I4 vs I8 -10,000 10,586 0,985 I4 vs I9 9,222 7,355 0,931 I5 vs I6 14,722 8,987 0,771 I5 vs I7 16,667 11,451 0,856 I5 vs I8 11,000 10,796 0,976 I5 vs I9 30,222 7,654 0,024 I6 vs I7 1,944 12,518 1,000 I6 vs I8 -3,722 11,921 1,000 I6 vs I9 15,500 9,175 0,744 I7 vs I8 -5,667 13,874 1,000 I7 vs I9 13,556 11,599 0,950 I8 vs I9 19,222 10,952 0,707
32
Attachment VII
Vigour at 162 days. Statistics results with the Games-Howell test to ascertain between what individuals existed statistical significant differences.
Games-Howell
Vigour 162 days Mean Difference (I-J) Std. Error Sig.
I1 vs I2 -14,833 11,308 0,913 I1 vs I3 2,722 9,939 1,000 I1 vs I4 -12,389 9,084 0,895 I1 vs I5 -30,778 9,206 0,078 I1 vs I6 -13,111 9,957 0,912 I1 vs I7 -17,000 13,178 0,919 I1 vs I8 -18,556 10,369 0,688 I1 vs I9 4,944 9,193 1,000 I2 vs I3 17,556 10,868 0,784 I2 vs I4 2,444 10,091 1,000 I2 vs I5 -15,944 10,201 0,809 I2 vs I6 1,722 10,884 1,000 I2 vs I7 -2,166 13,891 1,000 I2 vs I8 -3,722 11,262 1,000 I2 vs I9 19,778 10,190 0,601 I3 vs I4 -15,111 8,529 0,698 I3 vs I5 -33,500 8,659 0,029 I3 vs I6 -15,833 9,454 0,753 I3 vs I7 -19,722 12,802 0,819 I3 vs I8 -21,278 9,887 0,476 I3 vs I9 2,222 8,646 1,000 I4 vs I5 -18,889 7,663 0,346 I4 vs I6 -0,722 8,551 1,000 I4 vs I7 -4,611 12,151 1,000 I4 vs I8 -6.167,000 9,027 0,998 I4 vs I9 17,333 7,647 0,413 I5 vs I6 17,667 8,679 0,544 I5 vs I7 13,778 12,242 0,958 I5 vs I8 12,222 9,149 0,905 I5 vs I9 35,722 7,791 0,007 I6 vs I7 -3,889 12,816 1,000 I6 vs I8 -5,444 9,906 1,000 I6 vs I9 18,056 8,667 0,516 I7 vs I8 -1,556 13,139 1,000 I7 vs I9 21,944 12,232 0,686 I8 vs I9 23,500 9,137 0,275
33
Attachment VIII
Colour at 162 days for germination and vigour. Statistics results with the Games-Howell test to ascertain between what individuals existed statistical significant differences.
G e rmi na tio n
Games-Howell Mean Difference (I-J) Std. Error Sig.
Black vs Brown -16,981 5,426 0,009 Black vs Green -31,852 4,327 0,000 Brown vs Green -14,870 6,116 0,049 Vig o u r Black vs Brown -19,296 6,195 0,009 Black vs Green -18,426 4,885 0,001 Brown vs Green 0,870 6,716 0,991