Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 www.scielo.br/aabc
Structure, histochemistry and phytochemical profile of the bark of the
sobol and aerial stem of
Tontelea micrantha
(Celastraceae - Hippocrateoideae)
MARIA OLÍVIA MERCADANTE-SIMÕES1, HELLEN C. MAZZOTTINI-DOS-SANTOS1, LAYS A. NERY1, PERACIO R.B. FERREIRA2, LEONARDO M. RIBEIRO3,
VANESSA A. ROYO2 and DARIO A. DE OLIVEIRA2
1Laboratório de Anatomia Vegetal, Universidade Estadual de Montes Claros, Campus Darcy Ribeiro, Vila Mauricéia, 39401-089 Montes Claros, MG, Brasil 2
Laboratório de Bioprospecção e Recursos Genéticos, Universidade Estadual de Montes Claros, Campus Darcy Ribeiro, Vila Mauricéia, 39401-089 Montes Claros, MG, Brasil 3Laboratório de Micropropagação Vegetal, Universidade Estadual de Montes Claros,
Campus Darcy Ribeiro, Vila Mauricéia, 39401-089 Montes Claros, MG, Brasil
Manuscript received on March 18, 2013; accepted for publication on August 12, 2013
ABSTRACT
The bark of the underground stem of Tontelea micrantha (Mart. ex. Schult.) A. C. Sm., a native Brazilian Cerrado species, is used in folk medicine for treating kidney ailments. The structures of the underground and the aerial stems were examined and their barks were analyzed for the presence of secondary metabolites. Bark fragments were processed according to conventional techniques in plant anatomy and their chemical compositions examined using histochemical and phytochemical tests, thin layer chromatography, and high-efficiency liquid chromatography. The underground stem is a sobol with unusual cambial activity. Laticifers that secrete terpenoids were present in the cortex and phloem of both organs and can contribute to the identification of the species in field. Druses were present in both barks, but mono-crystals were only observed in the sobol. Tannins, flavonoids, alkaloids, and terpenoids occurred in both types of bark, but carotenoids were only detected in the sobol. The similarities between these two organs indicate that the aerial stem bark has potential medicinal use and represents a plausible alternative to harvesting the sobol, which could contribute to the preservation of natural populations of this species.
Key words: natural products, pharmacognosy, secondary metabolites, sobol.
Correspondence to: Maria Olívia Mercadante-Simões E-mail: [email protected]
INTRODUCTION
Representatives of the family Celastraceae are well
known in popular medicine for their pharmacological
properties, especially species of the genus
Maytenus
used for treating gastric ulcers (Leite et al. 2010,
Santos et al. 2007, Silva et al. 2011).
Cheiloclinium
and
Salacia
likewise comprise species that have been
examined in the search for bioactive compounds
with analgesic and anti-inflammatory activities
or for controlling diabetes (Duarte et al.
2010,
Tanabe
et al.
2008).
Tontelea micrantha
(Mart. ex. Schult.) A. C.
Sm.
is a native medicinal species of the Brazilian
Cerrado (savanna), and alcoholic extracts of the
bark of its underground stem are used for treating
kidney problems. The oil extracted from its seeds
is a potent anti-inflammatory and represents a
significant source of income for people living in
areas of Cerrado vegetation (Dias and Laureano
2010). Individuals of this species occur as clumps
(clones) of shrubs whose aerial shoots emerge
from an underground stem with a rather complex
anatomical structure (Wanderley et al.
2003). This
subterranean structure is responsible for re-sprouting
above-ground structures damaged by fires (a
common occurrence
in the Cerrado biome) (Silva et
al.
2009). The intensive human exploitation of these
plant organs for medicinal purposes threatens natural
populations of these plants. Although not usually
considered, harvesting only the aerial portions of
these plants instead of their subterranean structures
could minimize negative effects on plant growth
(Zschocke et al.
2000).
Governmental agencies responsible for
regu-lating and registering the production and use of natural
medicinal products require the correct botanical
identification of the species in order to prevent their
incorrect use and risks to consumer health (WHO
1987, Brasil 2000). Scientific research characterizing
the anatomical structures and chemical contents of
species utilized in traditional medicine has contributed
to controlling the quality of the dried and powdered
drugs obtained from them (Coelho et al.
2012, Cruz et
al. 2012, Ferreira et al. 2011).
The present study sought to: (i) identify
anatomical and chemical characteristics of the bark of
the underground and aerial stems of
T. micrantha
that
could be used as diagnostic features for the species;
(ii) identify similarities in the distributions of the
secondary metabolite classes found in the two organs
in order to evaluate the potential medicinal use of
aerial stem as a substitute for the underground system.
MATERIALS AND METHODS
PLANT MATERIAL
The “bark”,
sensu
Fahn (1990), of the underground
and aerial stems and the shoot apices of
T. micrantha
were examined. Study material was collected from
10 individuals in a natural population of this species
growing in the Cerrado (savanna) vegetation in
the municipality of Montes Claros, state of Minas
Gerais, Brazil (16° 52’ 15” S, 44° 00’ 58” W). Voucher
material was deposited in the BHCB herbarium of
the Departamento de Botânica of the Instituto de
Ciências Biológicas of the Universidade Federal de
Minas Gerais (Mercadante-Simões 2; registry number
214463; identified by Dr. Julio Lombardi).
STRUCTURAL EVALUATION
The material was fixed in Karnovsky’s solution
(Karnovsky 1965) under vacuum (560 mm Hg) for
12 hours, dehydrated in an ethanol series (Jensen
1962), and cold-embedded (Paiva et al.
2011) in
glycol-methacrylate resin (Leica Microsystem Inc.,
Heidenbeg, Germany). Transversal and longitudinal
sections (5 μm) were made using a rotary microtome
(Atako, Japan) and stained with toluidine blue, pH
4.7 (modified from O’Brien et al. 1964), fuchsin
(Johansen 1940),
floroglucinol (Johansen 1940), and
Sudan IV (Pearse 1980). Paradermal sections were
cleared in a 20% hypochlorite solution and stained
with safranin to examine the epidermal cells of the
aerial stem in frontal view (Johansen 1940). The
presence of calcium oxalate was verified using HCl
(Chamberlain 1932). Permanent slides were mounted
using Itacril acrylic resin (Itacril, Itaquaquecetuba,
Brazil). Photo-documentation was conducted using a
Canon A 620 digital camera (Canon, Tokyo, Japan)
coupled on a Nikon Eclipse E-200 optical microscope
(Nikon, Tokyo, Japan) and a digital camera (Zeiss
AxioCam HRc, Göttinger, Germany), using Axion
Vision image-capturing software, coupled on an
Olympus Optical model AX70 TRF light microscope
with a U-photo system.
HISTOCHEMICAL ANALYSES
Histochemical tests were performed on transverse
sections of fresh material obtained from the bark of
the underground and aerial stems of
T. micrantha
Rolemberg & Bhering, Belo Horizonte, Minas
Gerais, Brazil) using the following reagents: Lugol’s
solution for starch (Jensen 1962), Sudan IV (Pearse
1980) for lipids, bromophenol blue (Mazia et al.
1953) and Xilidine Ponceau for proteins (Vidal 1977);
vanillin-HCl for tannins (Mace and Howell 1974);
DMACA (
p
-dimethylaminocinannamaldehyde)
for flavonoids (Arnous 2002, Feucht et al. 1986);
Dittmar and Wagner reagents for alkaloids (Furr and
Mahlberg 1981); and NADI (naphtol and
dimethyl-paraphenylene-diamine) for terpenoids (David and
Carde 1964). Image documentation was performed
using a Canon A 620 digital camera (Canon, Tokyo,
Japan) coupled on a Nikon Eclipse E-200 optical
microscope (Nikon, Tokyo, Japan)
and a digital
camera (Zeiss AxioCam HRc, Göttinger, Germany),
using Axion Vision image-capturing software,
coupled on an Olympus Optical model AX70 TRF
light microscope with a U-photo system.
PHYTOCHEMICAL PROSPECTION
Bark from the underground and aerial stems of
T.
micrantha
was dried at room temperature and powdered
in a Willey-TE 64 mill (TECNAL, Pira cicaba, Brazil).
The resulting powder was stored at -18°C. Aqueous and
ethanol extracts of the bark were obtained by weighing
the powder (using an analytical balance; Shimadzu
BL320H, Tokyo, Japan) and macerating it in a water/
ethanol solution (1:10,V/V) three times every 24 hours;
the extracts were then filtered and concentrated under
reduced pressure at 35°C. The protocols described
by Barbosa et al.
(2001) and Mouco et al. (2003)
were employed for the extraction and identification
of tannins (using ferric chlorate), flavonoids (using
Shinoda and Bornträger’s reagent), alkaloids (using
Dragendorff and Mayer’s reagent), and terpenoids
(using Salkawski and Liberman-Burchard’s reagent).
HIGH PERFORMANCE LIQUID CHROMATOGRAPHY (HPLC)
The chemical constituents of the barks of the
underground and aerial stems of
T. micrantha
were
identified using a Waters
®chromatograph coupled
to a photodiode detector array and a phenomenex
ODS2 chromatography column (250 mm x 4.6 mm
x 5 μm) (Waters, Minneapolis, USA) with a flux rate
of 1 mL/minute and an injection volume of 20 μL.
The identifications of the chemical compounds were
performed by comparing their retention times with
external standards (using Empower 2 software).
Tannins: 4.0 g of the bark powder was extracted
with 10 mL of butanol. The liquid phase was then dried
at 35°C, and 115 mg of the extract was subsequently
diluted in 50 mL of methanol in an ultra-bath for 10
min. The chromatograph detector was adjusted to
270 nm, with butanol as the mobile phase and using
gallic, tannic, and ellagic acids as standards; the
run-time was 10 min at 25°C (Santos and Melo 2003).
Flavonoids: 20.0 g of the bark powder was
extracted with 30 mL of 85% ethanol at 45°C. Extract
was then filtered, dried at 35°C and resuspended in
methanol/acetonitrile/water (40:15:45 v/v/v) + 1%
acetic acid as the mobile phase. The chromatograph
detector was adjusted to 257 nm, with rutin and
quercetin as standards; the runtime was 15 min at
25°C (Lu et al.
2006).
Alkaloids: 3.0 g of the bark powder was
extracted with 6.0 g of magnesium oxide in 100
mL of distilled water at 100°C for 15 min. The
mixture was subsequently cooled and weighed, and
any water lost through evaporation was replaced
to 100 g above the original weight; chloroform
was used as the mobile phase. The mixture was
then centrifuged at 2000 rpm for 5 min and the
supernatant filtered through a 0.45 µm membrane.
The chromatograph detector was adjusted to 273
nm with a running time of 35 minutes at 25°C;
caffeine and theophylline were used as internal
standards (Alves and Bragagnolo 2002).
SPECTROPHOTOMETRY
from 1.0 g of the bark powder using 5.0 mL of acetone.
The extract was then fi ltered and dried at 35°C, resulting
in a residue of 0.015 g that was resuspended in 3 mL of
an ethanol/water solution (1:1) and diluted 1:30. The
chromatograph detector was adjusted for absorbance
measurements at 450 nm and carotenoid
concen-trations were determined in triplicate using standard
beta-carotene (Kimura and Rodriguez-Amaya 2002).
THIN LAYER CHROMATOGRAPHY (TLC)
To identify terpenoids and other volatile compounds,
200 g of the bark powder was extracted in 500 mL
of distilled water in four extraction sessions (50 g
of powder each) using a Clevenger apparatus; the
water phase was partitioned with hexane, which
was subsequently stored under freezing conditions
in the dark. The partition obtained was examined
by TLC using hexane: ethanol (9:1) as the mobile
phase; visualizations of the terpenoids and volatile
compounds were performed using sublimated iodine.
RESULTS
STRUCTURE AND HISTOCHEMISTRY
Underground stem
The underground stem of
T. micrantha
has a woody
consistency with an intense natural orange external
coloration that facilitates its identifi cation in the fi eld
(Figs. 1A-C). The stem-like nature of this organ
was confi rmed by the presence of pith (Fig. 1B).
Unusual cambial activity that produces concentric
and alternating layers of secondary xylem and
phloem can be observed in more advanced stages of
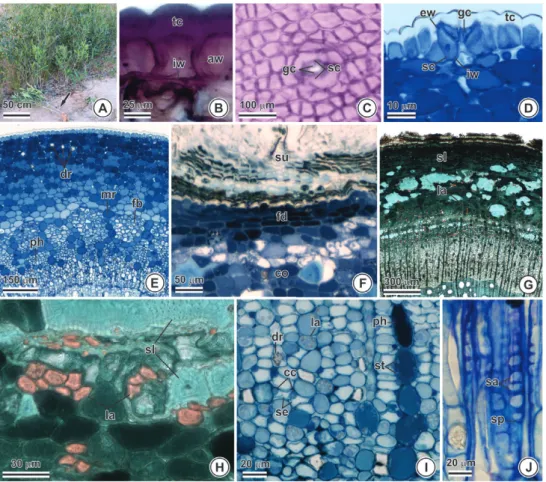
Figure 1 - Underground stem of Tontelea micrantha. (A-C) General aspect, showing its natural orange coloration (presence of carotenoids). (D-G) Transverse sections. (I) Longitudinal section. (B) Stem-like structure (presence of pith). (C) Unusual cambial activity. (D) Adventitious bud (arrow). Periderm and cortex.
development (Fig. 1C). Vegetative buds give rise to
aerial branches that form interconnected clumps of
plants that can cover large areas of land (Fig. 1D).
The periderm of the sobol has suber composed
of thin-walled cells that are predominantly
suberized, although some are lignified, with the
orange color typical of that structure; lenticels
can be observed; phellogen activity gives rise to a
compact phelloderm composed of layers of radially
disposed cells (Fig. E). The primary cortex is
well-developed on the secondary structure (Figs. B, E).
The parenchymatous layers of the phelloderm and
cortex show isolated or grouped large irregularly
shaped sclereids (with many pits); druses and
mono-crystals of calcium oxalate were also
observed (Figs. F-G). The secondary phloem shows
uniseriate rays composed of radially elongated
cells; non-articulated and non-anastomosing
laticifers with thick walls and viscous lipophilic
contents could be observed in the axially elongated
parenchyma; laticifers also occur in the cortex and
phelloderm (Figs. 1G-H).
Table I and Figure 2 show the results
of histochemical tests performed on the
underground stem bark. The natural color of
the material can be seen in the sections not
treated with reagents (Fig. 2A); the suber shows
carotenoids (Fig. 2B); the cortex contain starch
grains (Fig. 2C), protein reserves (Fig. 2D-E),
and secondary metabolic compounds such as
tannins (Fig. 2F), flavonoids (Fig. 2G), alkaloids
(Fig. 2H) and terpenoids (Fig. 2I-J).
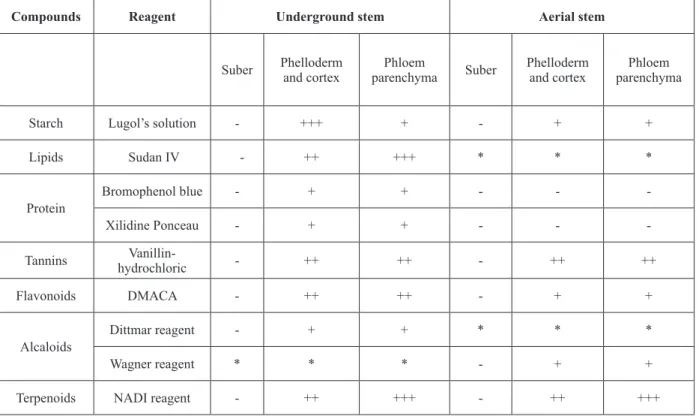
TABLE I
Results of the histochemical tests in the underground and aerial stems of T. micrantha. (+) presence; (-) absence.
Compounds Reagent Underground stem Aerial stem
Suber Phelloderm and cortex
Phloem
parenchyma Suber
Phelloderm and cortex
Phloem parenchyma
Starch Lugol’s solution - +++ + - + +
Lipids Sudan IV - ++ +++ * * *
Protein
Bromophenol blue - + + - -
-Xilidine Ponceau - + + - -
-Tannins
Vanillin-hydrochloric - ++ ++ - ++ ++
Flavonoids DMACA - ++ ++ - + +
Alcaloids
Dittmar reagent - + + * * *
Wagner reagent * * * - + +
Terpenoids NADI reagent - ++ +++ - ++ +++
Aerial stem
Adult individuals of
T. micrantha
produce nu merous
long and thin axially oriented branches that arise
from the underground stem, forming clumps of
clonal shrubs (Fig. 3A). The uniseriate epidermis on
the young branches is composed of cells covered by
a very thick cuticle that extends along the anticlinal
and inner periclinal walls (Fig. 3B). A frontal view
of the epidermis shows the polyhedral outlines of
the cells and the presence of ciclocytic stomata (Fig.
3C). The subsidiary cells have periclinal external
walls that are thinner than the internal walls, giving
them pyramid shapes in transverse section (Fig. 3D).
The cortex has small intercellular spaces, cells
with phenolic contents, druses and laticifers; the
stem structure is eustelic; the primary phloem has
a conspicuous cap of fi bers with pectic-cellulosic
walls and medullary rays that show accumulations
of phenolic compounds (Fig. 3E). The secondary
structure shows a multilayered suber that originated
from phellogen activity in the sub-epidermal layers
(Fig. 3F); the phelloderm is not well-developed and
Figure 2 - Transverse sections of the bark of the underground stem of Tontelea micrantha subjected to different histochemical tests (arrows show positive reactions). (A) Material not subjected to any reagents (white).
(B) Lipids stained red with Sudan IV (presence of carotenoids). (C) Starch grains stained purple with Lugol solution. (D, E) Proteins stained red with Ponceau xilidine and blue with bromophenol blue, respectively. (F)
Tannin stained reddish-brown with vanillin hydrochloride. (G) Flavonoids stained blue with DMACA reagent.
has tangentially elongated cells that are smaller
than the cortical cells (Fig. 3G); the cortex is
well-preserved in the secondary structure; groups
of large sclereids of variable sizes with small
lumens and walls with conspicuous pits can be
observed, as well as laticifers with thickened
walls and lipophilic contents (Fig. 3G-H). The
secondary phloem is well-developed, with radial
cells with phenolic contents, druses, and starch
grains (Fig. 3I). The sieve elements have oblique
sieve plates and numerous sieve areas (Fig. 3J).
Laticifers can be observed with the naked eye
when the stem is injured.
The results of the histochemical tests of the
aerial stems are presented in Table I and Figure 4.
The natural color of the material can be seen in the
sections not treated with reagents. (Fig. 4A). The
cortex shows the presence of starch grains (Fig. 4B),
tannins (Fig. 4C), fl avonoids (Fig. 4D) and alkaloids
(Fig. 4E). A conspicuous presence of elongated laticifers
was observed in the axial system (Figs. 4F-G); the
laticifers have elastic walls (Fig. 4G).
Figure 3 - Aerial stem of Tontelea micrantha. (A) General view. (B, D-I) Cross section. (B) Paradermal section. (J) Longitudinal section. (A) General aspect of the clone emerging from the underground stem (arrow). (B) Epidermal cells with the cuticle coating the anticlinal and periclinal walls. (C-D) Stomata.
(E) Primary structure. (F-J) Secondary structure. (F) Periderm and cortex. (G) Periderm, cortex with sclereids and laticifers, and secondary phloem. (H) Detail of the cortex showing sclereids and laticifers.
PHYTOCHEMICAL PROFILE
The results of the phytochemical tests corroborated
the results of the histochemical tests, indicating the
presence of tannins (Shalcowski: purple; Lieberman
Buchard: purple; and ferric chloride: green),
fl avonoids (Shinoda: red; and Bornträger: pink), and
alkaloids (Dragendorff: purple; and Mayer: turbid)
in both the underground and aerial stems.
High-performance liquid chromatography (HPLC),
using the liquid phase standards listed in Table II,
indicated the presence of tannins, fl avonoids,
and alkaloids. The retention times of the peaks,
considering tannic acid as a standard, were different
between the underground stem (3.683; 4.931; 6.998;
and 7.466) and the aerial stems (2.487 and 2.631).
The peaks for gallic acid and tannic acid were
the same (3.683; 4.931; 6.998; and 7.466) in the
underground stem; no retention peaks were observed
in the aerial stem using gallic acid.
In terms of the presence of fl avonoids, and
consid-ering the patterns generated by rutin, two peaks were
shared by the underground stem and the aerial stems
(2.843 and 3.276), with one additional peak exclusive to
the aerial stem (3.897). No peaks were seen when using
the quercetin standard. In terms of the analyzed alkaloids,
and in relation to theophylline as a standard, three peaks
were observed for the underground system (1.975; 2.845
and 3.426); and one different peak for the aerial stem
(3.553). No peaks were seen using the caffeine standard.
Spectrophotometric values indicated the presence
of carotenoids at concentrations of 0.0104 g/L (1% of
the plant material), as compared to the standard.
Caro-tenoids were only observed in the underground stem.
TABLE II
Results of the high-performance liquid chromatography (HPLC) in the underground and aerial stem of T. micrantha.
(A) major peak.
Metabolic group Standard Retention time (in minutes)
Underground stem Aerial stem
Tannins Tannic acid 3.683; 4.931
A; 6.998 and 7.466 2.487 and 2.631A
Gallic acid 3.683; 4.931A; 6.998 and 7.466
-Flavonoids Rutin 2.843
A and 3.276 2.843 A; 3.276 and 3.897
Quercetin -
-Alcaloids Teophylline 1.975; 2.845
A and 3.426 3.553
Caffeine -
-Thin layer
chromatography used to identify
terpenoids and other volatile compounds indicated
the presence of two bands from
the extraction of the
underground stem; the band with more apolar
charac-teristics was not seen in the stem extract (Fig. 5).
DISCUSSION
The stem-like nature of the underground stem of
T. micrantha
is indicated by the inner position of
the pith. The underground stem of this plant was
described as a xylopodium by Wanderley et al.
(2003), although xylopodium are tuberized structures
derived from the hypocotyl and primary root. The
morphology and anatomy of the underground stem
of
T. micrantha,
as described here, suggests that its
classifi cation as being a sobol, to be more correct
– a diffuse stem-like underground system growing
horizontally below the soil surface
(Appezzato-da-Glória 2003, Maroso et al. 2009).
Barks constitute the majority of all plant
sub-products used for folk medicinal purposes (Sen et al.
2010). Extracts of the root bark of
M. illicifolia
, for
example, have antifungal activity due to the presence
of the triterpenoids maitenin and pristimerin (Gullo
et al.
2012); the bark of the roots of
M. segalensis
has antimicrobial activity due to the presence of
maitenonic acid (Lindsey et al.
2012).
Anatomical attributes of the periderm are often
useful in identifying plant species (Pace et al. 2011).
Epidermis remnants of the bark situated externally
to the suber have diagnostic properties in drugs
prepared from those structures because of their
high mechanical resistance and their preservation
even in crushed and dehydrated material (Farias
et al. 2009). Ciclocytic stomata, with
pyramid-shaped subsidiary cells, have been observed on
the epidermis of the leaves of various species of
Celastraceae (Gomes et al. 2005).
Sclerenchyma organization is a taxonomic
character in this group, and has special importance
in identifying plant-derived drugs as they remain
well-preserved even after fragmentation (Soffiatti
and Angyalossy-Alfonso 1999).
T. micrantha
demonstrates a diffuse disposition of the sclereids
and fibers present in the cortical strip, which is
preserved in the secondary structures of both stems,
different from the continuous arrangement seen in
M. ilicifolia
(Duarte and Debur 2005). The presence
of druses is considered a universal characteristic
within the family Celastraceae and therefore has
no diagnostic value (Gomes et al. 2005), but the
observed presence of mono-crystals only in the
sobol could aid in distinguishing materials derived
from either the underground system or aerial stem
(Gomes et al. 2010).
The occurrence of laticifers in the Celastraceae
family can be considered a diagnostic character
for some species (Gomes et al. 2005, 2010). Their
presence has been associated with the vascular
system and with tissues that arose from the
fundamental meristem (Gomes et al.
2005, Lopes
et al. 2009), and they can accumulate secondary
metabolic compounds of medicinal value
(Monacelli et al. 2005). This is in agreement with
their occurrence in the cortex and phloem of both,
the underground and the aerial stem of
T. micrantha
.
The distribution of laticifers within different plant
organs constitutes an easily recognizable taxonomic
character, and the chemical compositions of their
protoplasts and the thicknesses of their cell walls
allows them to be readily identified in any tissue
in which they may occur (Jacomassi et al. 2007,
Pickard 2008). The elastic aspect of the laticifer
contents of
T. micrantha
can be used as a diagnostic
character in the field (Dias and Laureano 2010) and
is consistent with their terpenoid chemical nature.
The well-developed phloem of
T. micrantha
holds
significant numbers of laticifers.
Anatomical characters identified in the bark of
both the sobol and the aerial stem of
T. micrantha
which can be used as diagnostic criteria for identifying
that species include: the occurrence of lignified
layers in the suber; the disposition and morphology
of sclereids present in the cortex; and the occurrence
of resiniferous laticifers in the phloem.
The occurrence of secondary metabolic
compounds in the periderm, cortex and phloem
seems to be related to the medicinal value of the
bark of the sobol. The presence of these compounds
in the aerial stem suggests the possibility of
using those stems as an alternative to the
under-ground structure, which would contribute to the
preservation of the species.
Phenolic compounds have wide spectra of
medicinal uses (Santos and Mello 2003) and tannins
with anti-microbial activities have been identified in
Celastraceae species (Silva et al. 2011) used to treat
kidney inflammation (Pansera et al. 2003). These
obser-vations corroborate the popular use of
T. micrantha
in
treating renal infections (Dias and Laureano 2010).
The presence of rutin in Celastraceae has been
associated with the wound-healing properties and
anti-oxidant activity of this plant group (Tiberti et
al. 2007). Flavonoids act as co-factors of vitamin
C, lending it anti-inflammatory and antibacterial
properties (Zuanazzi and Montanha 2003).
presence in only some plant groups. Their detection
in
T. micrantha
and absence from the bark of
M.
rigida
(Estevam et al. 2009) indicates their possible
utility as a marker for these species.
Terpenoids have been described in various
species of Celastraceae and are known to have
numerous therapeutic properties (Costa et al. 2007,
Lorenzi and Matos 2002), and a number of species
of this family show potential for treating cancers
(Wang et al.
2012). Carotenoids, a chemical group
within the general class of tetraterpenoids, have
known biological activities (Maoka 2009, Niizu
and Rodriguez-Amaya 2005). The orange color of
the suber of the sobol of
T. micrantha
is the result of
the accumulation of carotenoids in that organ and
can be used to identify it in the field (Wanderley
et al. 2003, Dias and Laureano 2010). Considering
the medicinal importance of this class of plant
secondary metabolic products, the identification
of terpenoids in
T. micrantha
indicates its potential
value in anti-cancer screening projects.
CONCLUSIONS
The most distinctive difference between the sobol
and the aerial stem is the presence of carotenoids in
the former that can be easily identified by their strong
orange color (visible to the naked eye) and the
occur-rence of mono-crystals. The presence of compounds
with proven medicinal properties such as tannins,
alkaloids, flavonoids, and terpenoids in
T. micrantha
indicates its potential value in bio-prospection to
obtain new herbal medicines. The possibility of using
the bark of the aerial stem of
T. micrantha
(in place
of the sobol bark) would enable the sustainable use
of this species – one of many Cerrado plants that are
poorly known but severely threatened.
ACKNOWLEDGMENTS
The authors would like to thank the Fundação de
Amparo a Pesquisa do Minas Gerais (FAPEMIG)
for the Incentivo à Pesquisa e ao Desenvolvimento
Tecnológico grants, awarded to M.O.
Mercadante-Simões (CRA-BIPID-00152-12) and L. M. Ribeiro
(CRA-BIPIT-00137-11); Suelaine Barbosa for her
technical assistance; and Valdeci Leite Fonseca for
indicating the collection locality.
RESUMO
Tontelea micrantha (Mart. Ex. Schult.) A. C. Sm. é uma espécie nativa do Cerrado brasileiro cuja casca do caule subterrâneo é utilizada como medicinal no tratamento de doenças renais. As estruturas dos caules subterrâneo e aéreo foram estudadas e suas cascas avaliadas para a presença de classes de metabólitos secundários. Fragmentos das cascas foram processados de acordo com metodologias usuais em anatomia vegetal e submetidos às análises fitoquímicas colorimétricas, cromatografia em camada delgada e identificação química por cromatografia líquida de alta eficiência. O caule subterrâneo é um sóbole e apresenta atividade cambial não usual. Laticíferos que secretam terpenóides estavam presentes no córtex e floema de ambos os órgãos e podem contribuir para a identificação da espécie no campo. Drusas estão presentes em ambas as cascas, mas mono-cristais são observados apenas no sóbole. Taninos, flavonóides, alcalóides e terpenóides ocorrem em ambas as cascas, mas carotenóides são detectados apenas no sóbole. As semelhanças entre estes dois órgãos indicam que a casca do caule aéreo tem potencial para uso medicinal, representando uma alternativa plausível para o uso do sóbole, o que pode contribuir para a preservação de populações naturais da espécie.
Palavras-chave: produtos naturais, farmacognosia, meta bólitos secundários, sóbole.
REFERENCES
ALVES AB AND BRAGAGNOLO N. 2002. Determinação simultânea de teobromina, teofilina e cafeína em chás por cromatografia líquida de alta eficiência. Rev Bras Cienc Farm 38: 237-243. APPEZZATO-DA-GLÓRIA B. 2003.Morfologia de sistemas sub-terrâneos: histórico e evolução do conhecimento no Brasil, Ribeirão Preto: A.S. Pinto, 80 p.
ARNOUS A. 2002. Correlation of pigment and flavanol content
BARBOSA WLR. 2001. Manual para análise fitoquímica e
cromatográfica de extratos vegetais, Belém: U Fed Pará. BARNES PJ. 2003. Theophylline: new perspectives for an old
drug. Am J Resp Crit Care 167(6): 813-818.
BRASIL MS. 2000. Farmacopéia Brasileira, Brasília: Ministério da Saúde.
CHAMBERLAIN CJ. 1932. Methods in plant histology, Chicago: University of Chicago Press, 416 p.
COELHO VPM, LEITE JPV, NUNES LG AND VENTRELLA MC.
2012. Anatomy, histochemistry and phytochemical profile
of leaf and stem bark of Bathysa cuspidata (Rubiaceae). Aust J Bot 60: 49-60.
COSTA EA, SANTOS LR, PONTES IS, MATOS LG, SILVA GA AND
LIÃO LM. 2007. Analgesic and anti-inflammatory effects
of Cheiloclinium cognatum root barks. Braz J Pharmacog
17: 508-513.
CRUZ BP, CHEDIER LM, PEIXOTO PHP, FABRI RL AND PIMENTA
DS. 2012. Effects of light intensity on the distribution of anthocyanins in Kalanchoe brasiliensis Camb. And Kalanchoe pinnata (Lamk.) Pers. An Acad Bras Cienc 84: 211-217. DAVID R AND CARDE JP. 1964. Coloration différentielle dês
inclusions lipidique et terpeniques dês pseudophylles du
Pin maritime au moyen du reactif nadi. C R Acad Sci Paris D 258: 1338-1340.
DIAS JE AND LAUREANO LC. 2010. Farmacopéia popular do cerrado, Góias: Articulação Pacari, 352 p.
DUARTE LP, FIGUEIREDO RC, SOUSA GF, SOARES DBS,
RODRIGUES SBV, SILVA FC AND SILVA GDF. 2010.
Chemical constituents of Salacia elliptica (Celastraceae). Quim Nova 33: 900-903.
DUARTE MR AND DEBUR MC. 2005. Stem and leaf morpho-anatomy of Maytenus ilicifolia. Fitoterapia 76: 41-49. ESTEVAM CS, CAVALCANTI AM, CAMBUI EVF, ARAÚJO-NETO V,
LEOPOLDO PTG, FERNANDES RPM, ARAÚJO BS, PORFÍRIO
Z AND SANT’ANA AEG. 2009. Perfil fitoquímico e ensaio
microbiológico dos extratos da entrecasca de Maytenus rigida Mart. (Celastraceae). Rev Bras Farmacog19: 299-303. FAHN A. 1990. Plant anatomy, Oxford: Butteerworth-Heinemann Ltd. FARIAS V, ROCHA LD, PREUSSLER KH AND MARANHO
LT. 2009. Leaf structural organization of Pimenta pseudocaryophyllus (Gomes) L.R. Landrum, Myrtaceae. Acta Bot Bras 23: 398-406.
FERREIRA PRF, MENDES CSO, REIS SB, RODRIGUES CG,
OLIVEIRA DA AND MERCADANTE-SIMÕES MO. 2011.
Morphoanatomy, histochemistry and phytochemistry of
Psidium guineense Sw. (Myrtaceae) leaves. J Pharm Res
4: 942-944.
FEUCHT W, SCHMID PPS AND MARTINS RP. 1986. Distribution of flavonols in meristematic and mature tissues of Prunnus avium shoots. J Plant Physiol 125: 1-8.
FURR M AND MAHLBERG PG. 1981. Histochemical analyses of laticifers and glandular trichomes in Cannabis sativa. J
Nat Prod 44: 153-159.
GOMES SMA AND LOMBARDI JA. 2010. Leaf anatomy as a contribution to the taxonomy of Salacioideae N. Hallé ex Thorne & Reveal (Celastraceae). Plant Syst Evol 289: 13-33.
GOMES SMA, SILVA EAM, LOMBARDI JA, AZEVEDO AA
AND VALE FHA. 2005. Anatomia foliar como subsidio à taxonomia de Hippocrateoideae (Celastraceae) no Sudeste do Brasil. Acta Bot Bras 19: 945-961.
GULLO FP ET AL. 2012. Antifungal activity of maytenin and pristimerin. Evid Based Complement Alternat Med2012: 1-6. HENRIQUES AT, LIMBERGER RP, KERBER VA AND MORENO
PRH. 2003. Alcalóides: generalidades e aspectos básicos. In: SIMÕES CMO ET AL. (Eds), Farmacognosia: da planta ao medicamento, Porto Alegre/Florianópolis: UFRGS/ UFSC, Rio Grande do Sul/Santa Catarina, BR, p. 765-792.
JACOMASSI E, MOSCHETA IS AND MACHADO SR. 2007.
Morfoanatomia e histoquímica de Brosimum gaudichaudii
Trécul (Moraceae). Acta Bot Bras 21: 575-597.
JENSEN WA. 1962. Botanical histochemistry: principles and practice, San Francisco: W.H. Freeman & Company. JOHANSEN DA. 1940. Plant microtecnique. New York:
Macgraw-Hill Book.
KARNOVSKY MJ. 1965. A formaldehyde-glutaraldehyde
fixative of high osmolarity for use in electron microscopy.
J Cell Biol: 27: 137-138.
KIMURA M AND RODRIGUEZ-AMAYA DB. 2002. A scheme for obtaining standards and HPLC quantification of leafy vegetable carotenoids. Food Chem 78: 389-398.
LEITE JPV, BRAGA FC, ROMUSSI G, PERSOLI RM, TABACH R,
CARLINI EA AND OLIVEIRA AB. 2010. Constituents from
Maytenus ilicifolia. Leaves and bioguided fractionation for gastroprotective activity. J Brazil Chem Soc 21: 248-254. LINDSEY KL, BUDESINSKY M, KOHOUT L AND VAN STADEN
J. 2012. Antibacterial activity of maytenonic acid isolated from the root-bark of Maytenus senegalensis. S Afr J Bot
72: 473-477.
LOPES KLB, THADEO M, AZEVEDO AA AND MEIRA RMSA.
2009. Articulated laticifers in the vegetative organs of
Mandevilla atroviolaceae (Apocynaceae, Apocynoideae). Can J Bot 87: 202-209.
LORENZI H AND MATOS FJA. 2002. Plantas medicinais no Brasil: nativas e exóticas cultivadas. Instituto Plantarum de Estudos da Flora Ltda, São Paulo: Pantarum.
LU J, PAPP LV, FANG J, RODRIGUEZ-NIETO S, ZHIVOTOVSKY
B AND HOLMGREN A. 2006. Inhibition of Mammalian Thioredoxin Reductase by Some Flavonoids: Implications for Myricetin and Quercetin Anticancer Activity. Cancer Res 66: 4410-4418.
MACE ME AND HOWELL CR. 1974. Histochemistry and identification of condensed tannin precursor in roots of cotton seedlings. Can J Bot 52: 2423-2426.
MAOKA T. 2009. Sterically hindered carotenoids with 3 Z, 5 Z configuration from the seeds of oriental bitter sweet. Celastrus orbiculatus. Phytochemistry 70: 920-923. MAROSO RP, CARNEIRO CM, SCHEFFER-BASSO SM AND FAVERO
D. 2009. Morphological and anatomical aspects of birdsfoot trefoil and big trefoil. Rev Bras Zootectn 38: 1663-1667.
MONACELLI B, VALLETTA A, RASCIO N, MORO I AND PASQUA
G. 2005. Laticifers in Camptotheca acuminata Decne: distribution and structure. Protoplasma 226: 155-161.
MOUCO GB, BERNARDINO MJ AND CORNÉLIO ML. 2003.
Controle de qualidade de ervas medicinais. Biotecnol Ciênc Desenvolv 31: 68-73.
NIIZU PY AND RODRIGUEZ-AMAYA DB. 2005. New data on the carotenoid composition of raw salad vegetable. J Food Comp Anal 18: 739-749.
O’BRIEN TP, FEDER N AND MCCULLY ME. 1964. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 59:368-373.
PACE MR, LOHMANN LG AND ANGYALOSSY V. 2011. Evolution of disparity between the regular and variant phloem in Bignonieae (Bignoniaceae). Am J Bot 98: 602-618. PAIVA EAS, PINHO SZ AND OLIVEIRA DMT. 2011. Large
plant samples: how to process for GMA embedding? In: CHIARINI-GARCIA H ET AL. (Eds), Light microscopy: methods and protocols, New York: Springer/Humana Press, New York, USA, p. 37-49.
PANSERA MR, SANTOS ACA, PAESE K, WASUM R, ROSSATO M,
ROTA LD, PAULETTI GF AND SERAFINI LA. 2003. Análise
de taninos totais em plantas aromáticas e medicinais cultivadas no Nordeste do Rio Grande do Sul. Rev Bras Farmacog 13: 17-22.
PEARSE AGE. 1980. Histhochemistry theoretical and applied. Longman Group Limited.
PICKARD WF. 2008. Laticifers and secretory ducts: two other tube systems in plants. New Phytol 177: 877-888. SANTOS SC AND MELLO JCP. 2003. Taninos. In: SIMÕES CMO
ET AL. (Eds), Farmacognosia: da planta ao medicamento, Porto Alegre: UFSC, p. 615-656.
SANTOS VL, COSTA VBM, AGRA MF, SILVA BA AND BATISTA
LM. 2007. Pharmacological studies of ethanolic extracts of Maytenus rigida Mart (Celastraceae) in animal models. Rev Bras Farmacog17: 336-342.
SEN A, MIRANDA I, SANTOS S, GRAÇA J AND PEREIRA H. 2010. The chemical composition of cork and phloem in the rhytidome of Quercus cerris bark. Ind Crop Prod 31: 417-422. SILVA FC, DUARTE LP, SILVA GDF, FILHO SAV, LULA IS,
TAKAHASHIC JA AND SALLUM WST. 2011. Chemical constituents from branches of Maytenus gonoclada
(Celastraceae) and evaluation of antimicrobial activity. J Brazil Chem Soc 22: 943-949.
SILVA IA, VALENTI MW AND SILVA-MATOS DM. 2009. Fire effects on the population structure of Zanthoxylum rhoifolium Lam (Rutaceae) in a brazilian savanna. Braz J
Biol 69: 813-818.
SOFFIATTI P AND ANGYALOSSY-ALFONSO V. 1999. Estudo anatômico comparativo do lenho e da casca de duas espécies de Eugenia L. (Myrtaceae). Rev Bras Bot 22: 175-184.
TANABE G, SAKANO M, MINEMATSU T, MATUSDA H,
YOSHIKAWA M AND MURAOKA O. 2008. Synthesis and elucidation of absolute stereochemistry of salaprinol, another thiosugar sulfonium sulfate from the ayurvedic traditional medicine Salacia prinoides. Tetrahedron 64: 10080-10086.
TIBERTI LA, YARIWAKE HJ, NDJOKO K AND HOSTETTMANN
K. 2007. Identification of flavonols in leaves of Maytenus ilicifolia and M. aquifolium (Celastraceae) by LC/UV/MS analysis. J Chrom B846: 378-384.
VIDAL BC. 1977. Acid glycosaminoglycans and endochondral ossification: microespectrophotometric evaluation and macromolecular orientation. Cell Mol Biol 22: 45-64. WANDERLEY MG, SHEPHERD GJ, GIULLIETTI AM AND
MELLHEM TS. 2003. Flora Fanerogâmica do Estado de São Paulo, 3, São Paulo: Rima, p. 367.
WANG Y ET AL. 2012. Pristimerin causes G1 arrest, induces apoptosis, and enhances the chemosensitivity to gemcitabine in pancreatic cancer cells. Plos One 7: e43826. WHO. 1987. Quality control methods for medicinal plant
materials. Genebra: World Health Organization.
ZSCHOCKE S, RABE T, TAYLOR J, JAGER A AND VAN STADEN J.
2000. Plant part substitution - a way to conserve endangered medicinal plants? J Ethnopharmacol 71: 281-292.