Pró-Reitoria de Pós-Graduação e Pesquisa
Stricto Sensu em Ciências Genômicas e Biotecnologia
CARACTERIZAÇÃO BIOQUÍMICA E MOLECULAR DA
β
-GALACTOSIDASE DURANTE A MATURAÇÃO DE
FRUTOS DE
COFFEA ARABICA
Brasília - DF
2011
SÉRGIO ARAUJO FIGUEIREDO
CARACTERIZAÇÃO BIOQUÍMICA E MOLECULAR DA -GALACTOSIDASE DURANTE A MATURAÇÃO DE FRUTOS DE COFFEA ARABICA
Tese apresentada ao Programa de
Pós-Graduação Stricto Sensu em Ciências
Genômicas e Biotecnologia da Universidade Católica de Brasília, como requisito parcial para obtenção do Título de Doutor em Ciências Genômicas e Biotecnologia.
Orientador: Dr. Francisco José Lima Aragão
F475b Figueiredo, Sérgio Araujo
Caracterização bioquímica e molecular da β-galactosidase durante a maturação de frutos de Coffea arabica. / Sérgio Araujo Figueiredo – 2011.
244f.: il.; 30 cm
Tese (doutorado) – Universidade Católica de Brasília, 2011. Orientação: Francisco José Lima Aragão
1. Coffea arabica. 2. Polissacarídeos. 3. Biotecnologia. 4. Enzimas extracelulares. I. Aragão, Francisco José Lima, orient. II.Título.
Dedico este trabalho àqueles que têm estado ao meu lado ao longo desta jornada, sempre confiando em mim, amparando minhas incontáveis quedas com suas palavras de
encorajamento, com seus ombros amigos, com suas orações, com seu amor ...
Aos meus pais, Matosinho e Francisquinha; À minha eterna namorada, Cláudia;
AGRADECIMENTO
Registrar os nomes de todas as pessoas que me ajudaram ao longo deste tempo em que compartilhamos os mesmos objetivos é uma tarefa difícil de ser feita.
Nas parcas vitórias e nas vastas frustrações, na alegria contagiante de um resultado animador e nas lágrimas do desânimo por mais uma decepção. Na esperança do começo de um projeto novo e no desespero por ver esta esperança se esvair quando os experimentos, aparentemente, não dão certo. Nas refeições compartilhadas com alegria sincera e na divisão de um simples chocolate para matar a fome avassaladora de quem tem que tem ainda um longo caminho nas noites do laboratório. Nas conversas em que se aprende não somente o conhecimento técnico, mas se absorve um pouco do ser que nos ampara a ignorância. Um aprendizado constante e infinito, que não termina com a defesa desta tese, ciente de que esta não passa de mais uma pequena peça no grande quebra-cabeça do conhecimento humano.
Nomes serão esquecidos, isto é questão de memória; as pessoas com as quais aprendemos jamais o serão porque passaram a viver dentro de nós. Impossível citar todos, impossível não mencionar alguns ...
Minha eterna gratidão ao meu mentor, meu orientador e amigo, Francisco (Dr. Francisco José Lima Aragão) que me acolheu em seu laboratório quando o mundo fechava todas as portas para mim, ensinando-me não somente com suas palavras e experiências, mas com seus atos de nobreza moral;
À minha companheira de bancada, porto-seguro de todos no laboratório, que conquistou meu mais profundo respeito e amizade pelo ombro amigo, pelo silêncio nos meus dasabafos, pelos conhecimentos compartilhados em cima de uma ata de laboratório e pelos conselhos que nascem de um coração voltado para o bem maior, minha eterna gratidão à Elsa (Elsa Oliveira Paranaguá e Lago Nogueira);
Aos colegas, amigos e amigas que estiveram comigo durante estes anos, compartilhando experiências, lágrimas e sorrisos, fosse pelo tempo que fosse. A todos estes cujos nomes vibram em meu ser, minha profunda gratidão por tudo que passamos juntos;
Aos professores do Departamento de Botânica da Universidade de Brasília, pelo apoio e incentivo para que eu, mesmo cumprindo minhas obrigações acadêmicas junto à UnB, pudesse concluir esta etapa de minha formação profissional, em especial aos professores da área de Fisiologia Vegetal, prof. Dr. Augusto César Franco, prof. Dr. Fabian Borghetti, profa. Dr. Cristiane da Silva Ferreira e à profa. Dr. Lourdes Isabel Velho do Amaral;
À Lourdes (profa. Dra. Lourdes Isabel Velho do Amaral) não somente pelo aprendizado compartilhado na condução da disciplina na UnB, mas especialmente pelo incansável apoio nas incontáveis horas de discussão e de análises no HPLC, guiando-me os passos frágeis de um ser que ainda não sabe andar com os próprios pés nesta área específica que ela domina com propriedade. Nos medos compartilhados, nas decepções e nas alegrias de ver um simples gráfico de uma amostra sendo analisada em pleno final de semana em que ela poderia estar descansando ou recuperando-se de seu frágil estado de saúde. Minha eterna amizade e gratidão ...
para me ajudar com sua experiência e seu laboratório para a parte de cultura de tecidos do cafeeiro;
Aos membros de minha banca de defesa de tese, Vera (Dra. Vera Tavares de Campos Carneiro), Betânia (Dra. Betânia Ferraz Quirino), Lourdes (profa. Dra. Lourdes Isabel Velho do Amaral) e Thales (Dr. Thales Lima Rocha) pela sempre presteza e boa-vontade em me ajudar no que fosse necessário, minha profunda gratidão;
À D. Bel (Maria Isabel Martins da Costa) que, no anonimato e simplicidade de seu importante trabalho, sempre me deu o precioso apoio de seu sorriso, de sua experiência de vida e de sua fé inabalável em Deus, deixo registrado minha enorme gratidão;
À Universidade Católica de Brasília pela oportunidade de cursar o doutorado num ambiente de respeito e profissionalismo, ampliando minhas percepções acadêmicas com disciplinas ministradas por docentes altamente qualificados;
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pelo apoio financeiro e pela oportunidade de poder concluir meu doutoramento, minha gratidão;
Aos amigos do plano espiritual que, no silêncio do meu íntimo e no anonimato, estiveram ao meu lado não para cumprir aquilo que era de minha obrigação fazer, mas pelo apoio, incentivo e encorajamento para seguir adiante em todas as circunstâncias;
Ao Mestre Nazareno, pastor de minh’alma, meu guia, meu modelo, meu irmão-maior
e ...
“Há dois tipos de sabedoria: a inferior e a
superior.
A sabedoria inferior é dada pelo quanto uma pessoa sabe e a superior é dada pelo quanto ela tem consciência de que não sabe.
Tenha a sabedoria superior. Seja um eterno aprendiz na escola da vida.”
RESUMO
Referência: FIGUEIREDO, Sérgio Araujo. Título: Caracterização bioquímica e molecular
da -galactosidase durante a maturação de frutos de Coffea arabica. 2011. 243 folhas.
Tese (Ciências Genômicas e Biotecnologia) - Universidade Católica de Brasília, Brasília (DF), 2011.
As -galactosidases são uma classe de glicosil-hidrolases que atuam na parede primária das células vegetais, hidrolisando resíduos -D-galactosis de extremidades não redutoras de -D-galactosídeos presentes em diversas moléculas biológicas. Inicialmente foi feita uma caracterização dos monossacarídeos presentes na parede primária do pericarpo e do endosperma de frutos de Coffea arabica em distintas fases de maturação, identificando os polissacarídeos presentes nesta região celular, juntamente com os prováveis carboidratos-alvo para as -galactosidases. Em paralelo, foi realizada uma caracterização molecular e bioquímica das -galactosidases. Foi realizada uma caracterização parcial da estrutura do seu DNA genômico, juntamente com uma análise do nível de transcrição e da atividade bioquímica in vitro e in situ foram realizadas, identificando picos de expressão nas fases iniciais de crescimento e nos frutos completamente maduros. Por fim, visando avaliar os efeitos das -galactosidases na maturação de frutos de café, calos embriogênicos de C. arabica foram transformados por biobalística, utilizando a técnica do RNA interferente, com a finalidade de obtenção de plantas geneticamente modificadas de cafeeiro para o silenciamento da expressão do gene das -galactosidases. Foram obtidos três calos transgênicos crescendo em meio seletivo com glufosinato de amônio, dentre os quais dois continham o fragmento deste gene. Estes calos encontram-se em fase de formação de embriões somáticos e as plântulas resultantes desta regeneração serão analisadas, a posteriori, para confirmação da presença dos transgenes e avaliação dos efeitos do silenciamento do gene das -galactosidases sobre a maturação de frutos de café.
ABSTRACT
-galactosidases are a class of glycosyl-hydrolases that act on the plant cell primary walls, hydrolyzing -D-galactose at the nonreducing ends of -D-galactosides present in several biological molecules. Initially a characterization of the monosaccharides present in the primary wall of the pericarp and endosperm of Coffea arabica fruits at different ripening stages was performed, identifying the polysaccharides present in these regions, along with the putative carbohydrate target for the -galactosidase. In parallel, a molecular and a biochemical characterization of -galactosidase was performed. A partial characterization of
-galactosidase genomic DNA structure, along with a transcription analysis and an in vitro and in situ biochemical activity were performed, identifying peaks of expression in the early stages of growth and in fully ripe fruit. Finally, in order to evaluate the -galactosidase effects on coffee fruit ripening, C. arabica calli were transformed by biolistic using RNA interference approach, in order to obtain genetically modified coffee plants with a silenced -galactosidase expression. Three transgenic calli growing on selective medium containing ammonium glufosinate were obtained, two of which contained the -galactosidase gene fragment. These calli are under embryogenic regeneration and the resulting seedlings will be further analyzed in order to confirm the presence of the transgenes and to assess of the effects of -galactosidase gene silencing on coffee fruit ripening.
LISTA DE FIGURAS
U
Figura 1. Gráfico representativo da produção de café no Brasil (A) e da área plantada no território nacional (B) nos anos de 2005, 2006 e 2007. [Fonte: Brasil (2010)]U ... 22
U
Figura 2. Gráfico representativo das principais unidades federativas produtoras de café no Brasil na safra de 2009, considerando o número de sacas beneficiadas. [Fonte: Brasil (2010)]U
... 22
U
Figura 3. Produção dos principais países produtores de café na safra de 2010. [Fonte: Brasil (2010)]U ... 23
U
Figura 4. Classificação taxonômica e botânica de Coffea arabica.U ... 25
U
Figura 5. Fotos representativas do florescimento e maturação de frutos de C. arabica no campo. (A) Cafeeiro florescendo após a primeira chuva, findo o período de estresse hídrico indutor; (B e C) Cafeeiro com múltiplas florações, apresentando ramos com frutos e flores concomitantemente e (D) Amostragem de uma derriça parcial de frutos de café de um ramo com frutos em diversas fases de maturação.U ... 28
U
Figura 6. Esquema representativo do desenvolvimento de frutos de café. (A) Corte transversal de fruto maduro de café; (B) Fases do desenvolvimento de frutos de café ao longo do processo de maturação (Fonte: DE CASTRO e MARRACCINI, 2006).U ... 29
U
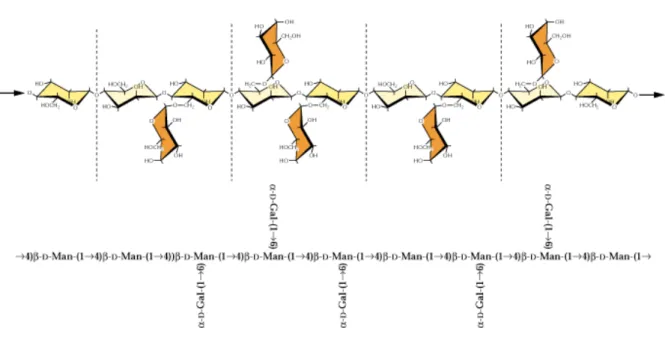
Figura 7. Esquema estrutural proposto para uma pectina da classe de arabinogalactanos de Coffea arabica (Fonte: REDGWELL et al., 2002).U ... 52
U
Figura 8. Esquema da estrutura da hemicelulose do tipo galactomananos encontrada na parede celular primária das plantas (Fonte: CARPITA e McCANN, 2000).U ... 53
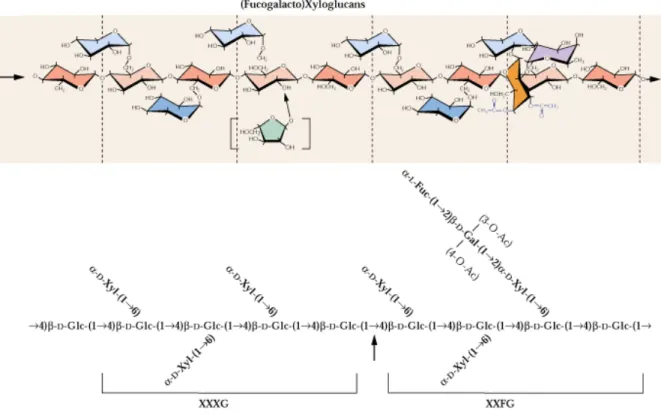
U
Figura 9. Esquema da estrutura da hemicelulose do tipo xiloglucano encontrada na parede celular primária das plantas (Fonte: CARPITA e McCANN, 2000).U ... 54
U
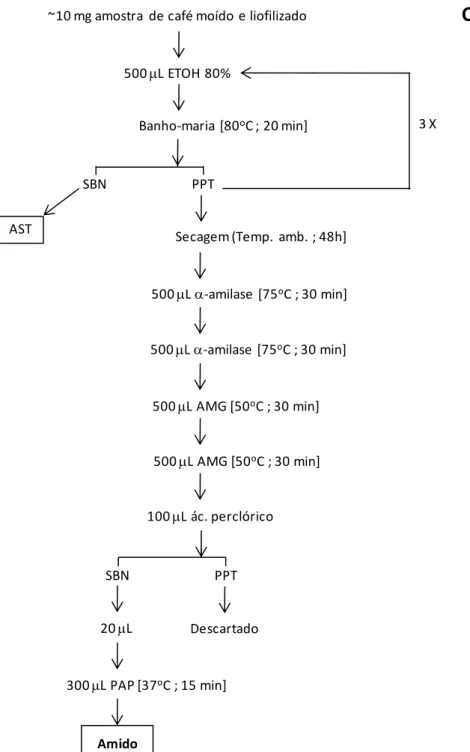
Figura 10. Esquema geral do protocolo para purificação dos monossacarídeos presentes na parede celular primária do pericarpo e do endosperma de frutos de C. arabica em diferentes estádios de desenvolvimento. (A) Procedimento para extração de lipídeos, ASTs, pool de arabinogalactano/galactomanano, proteínas, amido e para fracionamento da parede celular primária (extração com oxalato de amônio e série alcalina); (B) Procedimento de processamento dos componentes da parede celular primária para análise no ICS-3000 e de purificação/quantificação de celulose nas amostras; (C) Procedimento para quantificação de amido nas amostras de frutos de café.U ... 68
U
U
Figura 12. Percentagem (m/m) da fração lipídica presente no pericarpo e no endosperma de frutos de C. arabica em três fases distintas de maturação.U ... 71
Figura 13. Teores de açúcares solúveis totais presentes no pericarpo e no endosperma de frutos de C. arabica em três fases distintas de maturação.U ... 72
U
Figura 14. Quantidade de proteínas solúveis presentes no pericarpo e no endosperma de frutos de C. arabica em três fases distintas de maturação, com base na matéria seca.U ... 74
U
Figura 15. Percentagem (m/m) de amido presente no pericarpo e no endosperma de frutos de C. arabica em três fases distintas de maturação.U ... 76
U
Figura 16. Percentagem (m/m) de celulose presente no pericarpo e no endosperma de frutos de C. arabica em três fases distintas de maturação.U ... 77
U
Figura 17. Percentagem (m/m) de ácidos urônicos presentes no pericarpo e no endosperma de frutos de C. arabica em três fases distintas de maturação.U ... 78
U
Figura 18. Exemplo de cromatogramas obtidos de amostras hidrolisadas e purificadas do pericarpo e do endosperma de frutos de C. arabica em fases distintas de maturação, extraídos com diferentes soluções, no equipamento ICS-3000 (Dionex) com a coluna de separação CarboPac PA10 (Dionex) e pós-coluna com NaOH 0,5M. (A) Padrão de 10 M dos sete monossacarídeos básicos presentes na parede celular primária; (B) Pericarpo de frutos de café no estádio 5, extraído com oxalato de amônio 0,5%; (C) Pericarpo de frutos de café no estádio 4, extraído com NaOH 4M; (D) Pericarpo de frutos de café no estádio 7, extraído com NaOH 4M; (E) Endosperma de frutos de café no estádio 4, extraído com NaOH 1M; (F) Endosperma de frutos de café no estádio 7, extraído com NaOH 1M.U ... 81
U
Figura 19. Perfil da distribuição de monossacarídeos presentes no pericarpo de frutos de C. arabica em fases distintas de maturação (CV: pericarpo no estádio 4; CMV: pericarpo no estádio 5; CM: pericarpo no estádio 7; Fuc: fucose; Ara: arabinose; Rha: ramnose; Gal: galactose; Gli: glicose; Xil: xilose; Man: manose).U ... 85
U
Figura 20. Esquema proposto para as pectinas da classe dos ramnogalacturonano tipo I (RGI) (Fonte: CARPITA e McCANN, 2000).U ... 86
U
Figura 21. Esquema proposto para as hemiceluloses da classe dos glucuronoarabinoxilanos (GAX) (Fonte: CARPITA e McCANN, 2000).U ... 88
U
Figura 22. Perfil da distribuição de monossacarídeos presentes no endosperma de frutos de C. arabica em fases distintas de maturação (EV: endosperma no estádio 4; EMV: endosperma no estádio 5; EM: endosperma no estádio 7; Fuc: fucose; Ara: arabinose; Rha: ramnose; Gal: galactose; Gli: glicose; Xil: xilose; Man: manose).U ... 90
U
U
Figura 24. Provável sequência parcial codante do gene da -galactosidase de Coffea arabica. (A) Sequência de nucleotídeos (linha superior) e sua sequência de aminoácidos traduzida in silico (linha inferior). Aminoácidos estão apresentados por letra maiúscula. Predito peptídeo sinal está indicado em negrito. Predito sítio ativo da família da 35 glicosil hidrolases está sublinhado. (B) Diagrama esquemático da sequência genômica parcial clonada de CaGal indicando os exons (retângulos preenchidos) e os introns (linhas).U ... 120
U
Figura 25. Alinhamento de provável sequência de aminoácidos da -galactosidase de C. arabica, traduzida in silico, com a sequência de aminoácidos da -galactosidase de A. thaliana (CAB 64742). Quadrado vazado indica a localização do sítio ativo da família 35 glicosil hidrolase.U ... 123
U
Figura 26. Expressão da -galactosidase no pericarpo de frutos de C. arabica, crescido em condições de campo, em diferentes fases de maturação. (A) RT-PCR específicos para CaGal estão apresentados no painel superior; RT-PCR do gene usado como controle constitutivo Fator de alongamento 1 (EF-1) estão apresentados no painel inferior; (+) controle positivo: plasmídeo contendo um fragmento do gene de CaGal; (-) controle negativo: água; (B) Expressão relativa de CaGal nas diferentes fases de desenvolvimento de frutos de café.U .. 125
U
Figura 27. Expressão da -galactosidase no endosperma de frutos de C. arabica, crescida em condições de campo, em diferentes fases de maturação. (A) RT-PCR específicos para CaGal estão apresentados no painel superior; RT-PCR do gene usado como controle constitutivo Fator de alongamento 1 (EF-1) estão apresentados no painel inferior; (+) controle positivo: plasmídeo contendo um fragmento do gene de CaGal; (-) controle negativo: água; (B) Expressão relativa de CaGal nas diferentes fases de desenvolvimento de frutos de café.U .. 126
U
Figura 28. Atividade enzimática da -galactosidase, sob condições in vitro, expressa no pericarpo e no endosperma de frutos de C. arabica em diferentes fases de desenvolvimento (média de três repetições).U ... 127
U
Figura 29. Análise de Southern-blot de C. arabica Icatu Vermelho e C. arabica Catuai Vermelho de DNA genômico digerido com as enzimas de restrição DraI, EcoRI and HindIII, separadamente, e hibridizadas com sonda para o gene da -galactosidase. Marcadores de massas moleculares estão indicados.U ... 129
U
Figura 30. Expressão por RT-PCR de CaGal em órgãos vegetativos e reprodutivos de Coffea arabica; (+) controle positivo: plasmídeo contendo fragmentos do gene da -galactosidase; (-) controle negativo: água.U ... 131
U
U
Figura 32. Hipótese filogenética consenso da sequência deduzida de aminoácidos da -galactosidase de C. arabica com 36 sequências protéicas da -galactosidase usando a Máxima Parcimônia. A árvore filogenética foi construída usando o programa MEGA4 com 50% de cutoff. A percentagem de bootstrap aparece em cada ramificação da árvore.U ... 136
U
Figura 33. Esquema do funcionamento da técnica do RNA interferente (RNAi) em plantas [Fonte: MELLO e CONTE JR et al. (2007)]U ... 148
U
Figura 34. Esquema de construção do vetor para silenciamento do gene da -galactosidase de Coffea arabica por RNAi. (A) e (B) etapas da construção do vetor de transformação pKANGALRNAi.U ... 158
U
Figura 35. Sequência de preparo de calos embriogênicos de C. arabica para bombardeamento. (A) calos embriogênicos em meio indutor sólido de calos; (B) calos embriogênicos em meio líquido sob agitação; (C e D) detalhes do crescimento de calos embriogênicos em meio líquido; (E) calos embriogênicos em meio de multiplicação sólido contendo 0,5 M de manitol imediatamente antes de ser bombardeado com o vetor de transformação para silenciamento do gene da -galactosidase; (F) equipamento de transformação por biobalística utilizado na transformação de calos embriogênicos de café (EMBRAPA Recursos Genéticos e Biotecnologia – Laboratório de Transferência de Genes).U ... 163
U
Figura 36. Vetor pBI426 utilizado nos testes de expressão transiente do gene uidA para definição dos parâmetros de transformação de calos embriogênicos de C. arabica por biobalística.U ... 164
U
Figura 37. Expressão transiente do gene gus em calos embriogênicos de C. arabica. (A) controle não bombardeado; (B) calos bombardeados com espermidina; (C e D) calos bombardeados com protamina. Setas indicam pontos azulados devido à expressão da -glucuronidase. Barra: 5 mm.U ... 165
U
Figura 38. Amostras representativas da curva de seleção de calos embriogênicos de C. arabica submetidos a diferentes concentrações de glufosinato de amônio, após 20 dias de tratamento no escuro.U ... 166
U
Figura 39. Crescimento de calos embriogênicos de C. arabica transformados com vetor pKANGALRNAi por biobalística em meio de multiplicação de calos (meio CPmod) contendo agente seletivo (5 mg.L-1 de herbicida glufosinato de amônio). (A) calos bombardeados sem regeneração (B) calos bombardeados apresentando setores embriogênicos se multiplicando; setas indicam os calos no 1 e no 2; (C) ampliação do setor embriogênico no 1; (D) ampliação do setor embriogênico no 2. Barra: 5 mm.U ... 167
U
U
Figura 41. Teste imunológico para detecção da proteína PAT utilizando o kit imunológico
Trait™ LL Test Strip. (1) Controle positivo: semente transgênica de Zea mays expressando a
proteína PAT (não fervida); (2) Controle positivo: semente transgênica de Zea mays expressando a proteína PAT (fervida); (3) Controle negativo: calos de C. arabica não transgênico (não fervido); (4) Controle negativo: calos de C. arabica não transgênico (fervido); (5) Provável calo transgênico no 1 de C. arabica (não fervido); (6) Provável calo transgênico no 1 de C. arabica (fervido); (7) Provável calo transgênico no 2 de C. arabica (fervido); (8) Provável calo transgênico no 3 de C. arabica (fervido). Seta indica reação positiva para a proteína PAT.U ... 169
U
LISTA DE TABELAS
U
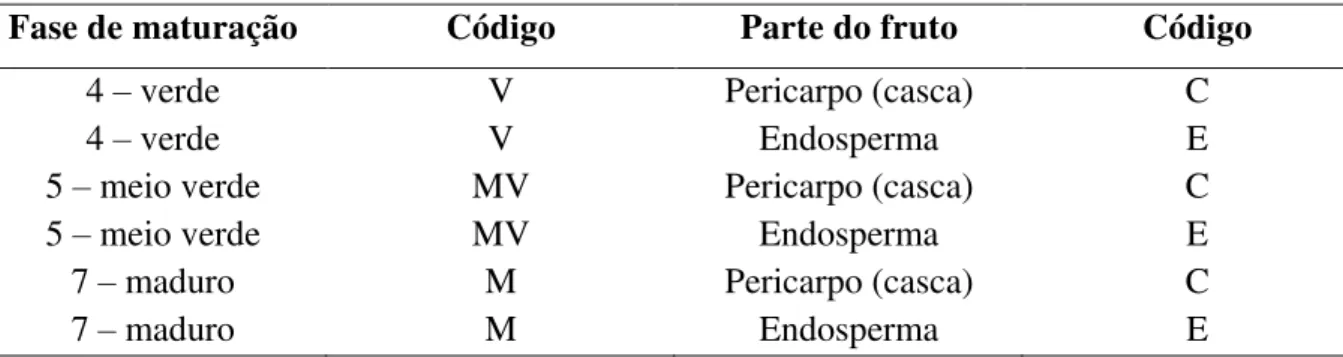
Tabela 1. Códigos utilizados para a identificação das amostras de frutos de C. arabica analisadas quanto à composição bioquímica dos componentes presentes nos diversos estádios de seu desenvolvimento.U ... 70
U
Tabela 2. Composição dos monossacarídeos que constituem as paredes celulares primárias do pericarpo e do endosperma de frutos de Coffea arabica em diferentes fases de desenvolvimento.U ... 82
U
Tabela 3. Rendimentos da extração das frações das amostras de pericarpo e de endosperma de frutos de C. arabica em diversos estádios de maturação (CV: pericarpo de fruto no estádio 4; CMV: pericarpo de fruto no estádio 5; CM: pericarpo de fruto no estádio 7; EV: endosperma de fruto no estádio 4; EMV: endosperma de fruto no estádio 5; EM: endosperma de fruto no estádio 7)U ... 83
U
Tabela 4. Resumo geral das classes de polissacarídeos encontrados no pericarpo e no endosperma de frutos de Coffea arabica em três fases de maturação (estádios 4, 5 e 7).U ... 92
U
Tabela 5. Histórico dos métodos de transformação de plantas utilizados em Coffea sp.U ... 152
U
Tabela 6. Resumo geral das classes de polissacarídeos encontrados em frutos de Coffea arabica em diversas fases de maturação, identificando as classes que contém resíduos de galactose na sua estrutura e o respectivo tipo de ligação deste monossacarídeo.U ... 183
U
SUMÁRIO
SUMÁRIO ... 16
Introdução Geral e Revisão Bibliográfica ... 18
Revisão bibliográfica ... 19
1.1. Café: origem e história ... 19
1.2. O café e a economia brasileira ... 21
1.3. Caracterização genética e botânica do cafeeiro ... 24
1.4. O florescimento e frutificação do cafeeiro ... 26
1.5. A maturação dos frutos de café: aspectos agronômicos e bioquímicos ... 26
1.6. As -galactosidases ... 31
1.7. Caracterização estrutural da parede celular vegetal ... 32
1.8. Polissacarídeos da parede celular primária de plantas ... 34
1.9. Silenciamento gênico por RNA interferente ... 37
Justificativas ... 39
Hipótese geral da tese ... 41
Objetivo (geral) ... 42
Objetivos específicos ... 42
Referências ... 43
Capítulo I – Caracterização dos carboidratos da parede primária de frutos de Coffea arabica em diferentes fases de desenvolvimento ... 49
Introdução ... 50
Objetivos ... 56
Material e Métodos ... 57
Resultados ... 69
Discussão ... 93
Referências ... 98
Capítulo II – Clonagem do gene da -galactosidase e análise molecular e bioquímica da sua expressão durante o desenvolvimento de frutos de Coffea arabica ... 103
Objetivos ... 108
Material e Métodos ... 108
Resultados e Discussões ... 116
Referências ... 138
Capítulo III – Silenciamento do gene da -galactosidase em plantas de Coffea arabica utilizando RNAi (RNA interferente) ... 144
Introdução ... 145
Objetivo Geral ... 154
Objetivos Específicos ... 155
Material e Métodos ... 155
Resultados e Discussões ... 163
Referências ... 175
Capítulo IV – Discussão Geral e Perspectivas ... 181
Discussão Geral e Perspectivas ... 182
Referências ... 188
Anexo 1 – Sequência do gene da -galactosidase de Coffea arabica depositada no GenBank (NCBI) ... 191
Anexo 2 – Artigo relacionado com a tese, publicado na Journal of Experimental Botany .... 195
REVISÃO BIBLIOGRÁFICA
1.1. CAFÉ: ORIGEM E HISTÓRIA
Apesar do consenso de que o centro de origem do café seja a região nordeste do continente Africano na região de Kaffa [(região compreendida entre a Etiópia e o Sudão (LASHERMES et al., 1999)], foram os árabes os responsáveis pela propagação do seu cultivo e sua disseminação, o que resultou na denominação científica da principal espécie de café cultivada, Coffea arabica. O café chegou à Arábia no séc. XIII, de onde foi transferido para a Turquia durante o século XVI. Foi somente no início do século XVII que o café começou a ser conhecido e apreciado na Europa, trazido pelos comerciantes em suas expedições pelo Oriente Médio (ASSOCIAÇÃO BRASILEIRA DA INDÚSTRIA DO CAFÉ, 2009). As primeiras mudas foram plantadas no Jardim Botânico de Paris e Jardim Botânico de Amsterdam, mas as condições climáticas européias não eram favoráveis à cafeicultura. Em 1714, os holandeses trouxeram o café para o Novo Mundo, onde o clima se mostrou favorável à cultura, rapidamente espalhando-se pelo continente americano (JOLY e HERMÓGENES, 1979).
No Brasil, o café foi introduzido em 1727, inicialmente na região Norte, mais precisamente em Belém, trazido da Guiana Francesa para o Brasil pelo Sargento-Mor Francisco de Mello Palheta a pedido do governador do Maranhão e Grão Pará, que o enviara às Guianas com essa missão. Graças às condições climáticas favoráveis, o cultivo de café rapidamente se espalhou pelas demais regiões brasileiras. O café chegou ao Rio de Janeiro por volta do ano 1760, vindo do Maranhão, plantadas por padres capuchinhos em sua horta. Dessa plantação, mudas e sementes se espalharam pelo atual estado do Rio de Janeiro, tendo a província de Vassouras se transformado na capital do café brasileiro nas primeiras décadas do séc. XIX.
Num espaço de tempo relativamente curto, o café assumiu a posição de produto-base da economia brasileira. Findo o ciclo da mineração, o ciclo do café durou por quase um século, sendo esse fruto considerado a grande riqueza brasileira. As divisas geradas pela economia cafeeira aceleraram o desenvolvimento do Brasil e o inseriram nas relações comerciais internacionais, possibilitando inclusive o surgimento de cidades e dinamização de importantes centros urbanos por todo o interior do Estado de São Paulo, sul de Minas Gerais e norte do Paraná. Foram construídas ferrovias para permitir o escoamento da produção, substituindo o transporte animal e impulsionando o comércio inter-regional de outras importantes mercadorias.
Findo o período da escravatura no Brasil (1888), a cultura cafeeira trouxe grandes contingentes de imigrantes no final do século XIX e início do século XX, vindos principalmente da Europa, consolidando a expansão da classe média, diversificando investimentos e incentivando movimentos culturais. O impacto da cafeicultura na economia brasileira neste período chegou a influenciar de forma significativa o meio político da
incipiente República Brasileira com a política do “café com leite”. Esta política foi um acordo
firmado entre as oligarquias estaduais e o governo federal durante a República Velha para que os presidentes da República fossem escolhidos entre os políticos de São Paulo e Minas Gerais e a origem deste nome era uma alusão à economia de São Paulo e Minas, grandes produtores de café e leite, respectivamente. Formalmente, a política do café-com-leite teve início em 1898, no governo do paulista Manuel Ferraz de Campos Salles e encerrou-se em 1930, com a chegada de Getúlio Vargas ao poder. Deste período em diante, o café e o povo brasileiro passam a ser indissociáveis.
A economia brasileira, dependente da exportação do café no cenário internacional, sofreu fortíssimo abalo com a quebra na bolsa de Nova York em outubro de 1929, modificando o cenário da cafeicultura no Brasil devido à brusca queda do seu preço. Milhões de sacas de café estocadas foram queimadas e milhões de pés de café foram erradicados, na tentativa de estancar a queda contínua de preços provocada pelos excedentes de produção.
mesmo perdendo mercado para outros países produtores, o Brasil ainda se mantém como maior produtor de café do mundo.
Das suas épocas áureas, ainda nos restaram as sedes das fazendas coloniais, um extenso material técnico-científico, plantações centenárias e o hábito nacional do cafezinho.
1.2. O CAFÉ E A ECONOMIA BRASILEIRA
O cafeeiro é uma cultura perene de alto valor sócio-econômico e uma das principais
commodities nos diversos países produtores de café, inclusive no Brasil, destacando-se no cenário econômico nacional e internacional como uma commodity e sendo negociada na BM&FBOVESPA (Bolsa de Mercadorias e Futuro – BOVESPA – Brasil) e na Bolsa de Nova Iorque (New York Stock Exchange – EUA). Esta valorização do produto “café” deve-se à demanda do mercado consumidor por esta commodity, cujos grãos são processados para obtenção de bebida de alta aceitação em diversas partes do mundo.
Considerando apenas as culturas perenes plantadas no Brasil, segundo os dados do Ministério da Agricultura, Pecuária e Abastecimento (BRASIL, 2010) em termos de produção total, a cafeicultura ocupa a terceira posição no cenário nacional, superada apenas pela cultura da laranja e da banana (Figura 1 – A). No entanto, em termos de área plantada, a cafeicultura ocupa uma área de, aproximadamente, 2,3 milhões de hectares, ou seja, três vezes maior que a área plantada com a cultura da laranja (Figura 1 – B).
0 2000 4000 6000 8000 10000 12000 14000 16000 18000 20000
Café Banana Laranja Uva Cacau
Pr
o
d
u
çã
o
(m
il
to
n
)
2005
2006
2007
A
Figura 1. Gráfico representativo da produção de café no Brasil (A) e da área plantada no território nacional (B) nos anos de 2005, 2006 e 2007. [Fonte: Brasil (2010)]
Dentre as diversas espécies de café descritas na literatura, no Brasil aproximadamente 70% da produção total de grãos de café in natura é de Coffea arabica;os demais 30% são de
Coffea canephora. Considerando os estados produtores de café, Minas Gerais destaca-se no cenário nacional por ser o principal produtor com, aproximadamente, 50% do total de sacas beneficiadas (BRASIL, 2010) (Figura 2).
Figura 2. Gráfico representativo das principais unidades federativas produtoras de café no Brasil na safra de 2009, considerando o número de sacas beneficiadas. [Fonte: Brasil (2010)]
0 500 1000 1500 2000 2500
Café Banana Laranja Uva Cacau
Á
re
a
p
la
n
ta
d
a
(m
il
h
a)
2005
2006
2007
B
50.3%
25.9% 8.4%
3.8% 4.7% 4.1%
0.4% 0.5% 0.7% 1.2%
Minas Gerais Espírito Santo São Paulo Paraná Bahia Rondônia Mato Grosso Pará
Segundo o Informe Estatístico do Café, publicado pelo Ministério da Agricultura, Pecuária e Abastecimento (BRASIL, 2010), em 2009, a produção de café no Brasil foi da ordem de 39 milhões de sacas, dos quais 50,5% foram destinados à exportação. Em termos de exportação, estes valores representam, aproximadamente, 6,1% do total de commodities exportado em 2009.
Apesar da grande competição no cenário internacional, o Brasil ainda se mantém como o principal país exportador de café (Figura 3), seguido do Vietnã e da Colômbia (BRASIL, 2010; Food and Agriculture Organization of the United Nations, 2010).
Figura 3. Produção dos principais países produtores de café na safra de 2010. [Fonte: Brasil (2010)]
No mercado interno, o consumo de café teve um incremento crescente desde 1990, passando de 8,4 milhões de sacas em 1990 para 18,2 milhões de sacas de café em 2009, ou seja, mais que dobrou nos últimos 20 anos (ASSOCIAÇÃO BRASILEIRA DA INDÚSTRIA DO CAFÉ, 2009), indicando seu impacto e sua importância para a economia brasileira.
Devido ao seu destacado aspecto sócio-econômico, existe um significativo investimento público e privado no agronegócio do café. No setor público nacional, diversos institutos de pesquisa e fundações têm programas direcionados à cultura do cafeeiro como, por exemplo, a Empresa Brasileira de Pesquisa Agropecuária voltada à cafeicultura (EMBRAPA Café), o Instituto Agronômico de Campinas (IAC) e o Instituto Agropecuário do Paraná (IAPAR), dentre outras. Além destes, diversas universidades públicas federais como a
32%
15%
7% 9%
4% 4% 4% 3% 3% 3% 2% 1%
1%
12%
Brasil
Vietnã
Colômbia
Indonésia
Etiópia
Índia
México
Guatemala
Peru
Honduras
Costa do Marfim
Nicaragua
El Salvador
Universidade Federal de Viçosa (UFV) e a Universidade Federal de Lavras (UFLa) também investem no desenvolvimento científico-tecnológico do cafeeiro. Vale destacar o Projeto Genoma do Café (Brazilian Coffee Genome Project), de cunho nacional e multi-institucional, que visa à formação de um banco de ESTs (Expressed Sequence Tags) de várias espécies de café (C. arabica, C. canephora e C. racemosa) de forma a ampliar os conhecimentos moleculares relacionados com esta cultura, fornecendo subsídios para o melhoramento genético do café.
Investimentos significativos na pesquisa da cultura do café no cenário internacional têm sido direcionados visando a ampliação dos conhecimentos na área da genômica desta cultura, dada a sua importância econômica como uma commodity de alto valor. Kochko et al. (2010) faz uma revisão ampla neste tópico que merece a apreciação dos pesquisadores envolvidos na área de genômica do cafeeiro.
No âmbito privado nacional, diversas empresas têm concentrado esforços para o aumento da produção e comercialização desta commodity como, por exemplo, a Fundação Procafé, uma entidade privada composta por sindicatos e cooperativas vinculadas ao café.
Neste cenário, merece destaque a publicação de um artigo na revista Science em 2010 no qual foi relatado que o Brasil é o país com o maior número de publicações relacionadas com culturas de grande impacto na economia mundial como a cana-de-açúcar, a laranja e o café (REGALADO, 2010). Vale ressaltar que os investimentos em pesquisa na cultura do cafeeiro estão relacionados não somente com o aumento da produtividade e/ou resistência ao ataque de insetos e fitopatógenos, mas também na busca dos fatores genéticos e bioquímicos que poderiam estar relacionados com a qualidade da bebida de café, características estas de elevada importância para o mercado consumidor (LEROY et al., 2006), principalmente porque há uma relação direta entre a qualidade da bebida de café e seu valor de mercado.
Em função da destacada importância sócio-econômica desta cultura no cenário nacional e internacional, percebe-se que a cafeicultura representa um dos principais agronegócios de impacto na balança comercial brasileira.
1.3. CARACTERIZAÇÃO GENÉTICA E BOTÂNICA DO CAFEEIRO
Segundo o National Resources Conservation Services (UNITED STATES, 2009), a planta de café apresenta a seguinte classificação taxonômica (Figura 4):
Super-reino Eucariota
Reino Viridiplantae
Subreino Tracheofita
Superdivisão Espermatofita
Divisão Magnoliofita
Classe Magnoliopsida
Subclasse Asteridae
Ordem Rubiales
Família Rubiaceae
Gênero Coffea
Espécie Coffea arabica L.
Figura 4. Classificação taxonômica e botânica de Coffea arabica.
Dentro do gênero Coffea estão catalogadas 103 espécies e, considerando apenas as espécies utilizadas na produção de bebida, as mais utilizadas no cenário mundial são Coffea arabica, Coffea canephora e Coffea liberica (DAVIS et al., 2006). No Brasil, as principais espécies cultivadas e que dominam o mercado são Coffea arabica (70%) e Coffea canephora (30%). Estas duas espécies possuem diferenças genéticas e reprodutivas contrastantes: C.
arabica é uma espécie alotetraplóide (2n – 4x – 44 cromossomos) e, predominantemente, autógama com, aproximadamente, 90% de autofecundação, enquanto C. canephora e outras espécies de café são diplóides (2n – 2x – 22 cromossomos) e são alógamas obrigatórias, apresentando autoincompatibilidade (FAZUOLI et al., 1999). Esta característica reprodutiva faz com que exista uma maior variabilidade genética em variedades de C. canephora do que em C. arabica, com implicações diretas sobre os programas de melhoramento do cafeeiro.
1.4. O FLORESCIMENTO E FRUTIFICAÇÃO DO CAFEEIRO
As plantas do gênero Coffea apresentam um ciclo fenológico composto por uma fase vegetativa e por uma fase reprodutiva subdividida em gema dormente, gema intumescida, abotoado, florada, pós-florada, chumbinho, expansão dos frutos, grão verde, verde cana, cereja, passa e seco (PEZZOPANE et al., 2003). Considerando a sazonalidade fenológica do café, o florescimento ocorre na primavera, a frutificação no verão, a maturação dos frutos no outono e o repouso no inverno.
O órgão da planta de café com interesse comercial é o fruto. Este fruto é do tipo drupa, com duas lojas independentes de polinização, os quais poderão dar origem a duas sementes. Na sua constituição, os frutos apresentam (i) o pericarpo (composto por exocarpo – casca que apresenta a coloração típica do fruto; mesocarpo – polpa ou mucilagem; endocarpo –
pergaminho), (ii) película prateada (testa do tegumento) e (iii) endosperma (grão) (ANTUNES FILHO e CARVALHO, 1954).
Considerando o florescimento do cafeeiro, merecem destaque os fatores abióticos como fotoperiodismo e temperatura que podem agir como sinalizadores ambientais para a sua indução em plantas sensíveis a estes fatores, coordenando o início da fase reprodutiva (TAIZ e ZEIGER, 2009; VAZ et al., 2008). Entretanto, outros fatores ambientais podem ser considerados sinalizadores ambientais, agindo independentemente ou em conjunto com o fotoperiodismo e/ou temperatura para a indução do florescimento. Na cultura do cafeeiro, um destes fatores ambientais que se destaca é a água, ou melhor, a disponibilidade de água para as plantas após um período de estresse hídrico (déficit hídrico) (ALVIM, 1960). É sabido que o regime hídrico exerce um papel determinante no florescimento do cafeeiro, embora as razões fisiológicas envolvidas ainda não tenham sido elucidadas.
1.5. A MATURAÇÃO DOS FRUTOS DE CAFÉ: ASPECTOS AGRONÔMICOS E
BIOQUÍMICOS
Apesar de ser um dos principais produtos de exportação, o Brasil vem perdendo mercado devido ao aumento da competitividade do mercado internacional cada vez mais exigente por grãos de café de qualidade superior (CAIXETA, 2001).
Por outro lado, ao longo das diversas etapas de produção de café (desde o campo até o beneficiamento e estocagem), vários fatores podem afetar a qualidade final dos grãos de café como, por exemplo, a espécie cultivada, as práticas agronômicas de manejo da cultura (controle de pragas e doenças, adubação e irrigação, dentre outras), bem como as condições edafoclimáticas e as práticas de colheita, processamento e beneficiamento dos grãos de café (DE MELLO, 2001).
Figura 5. Fotos representativas do florescimento e maturação de frutos de C. arabica no campo. (A) Cafeeiro florescendo após a primeira chuva, findo o período de estresse hídrico indutor; (B e C) Cafeeiro com múltiplas florações, apresentando ramos com frutos e flores concomitantemente e (D) Amostragem de uma derriça parcial de frutos de café de um ramo com frutos em diversas fases de maturação.
Além disso, o lento e complexo processo de maturação de frutos pode refletir também, numa desuniformidade de maturação de frutos de café provenientes de uma mesma florada (DE CASTRO e MARRACCINI, 2006) (Figura 6).
C
A
D
Figura 6. Esquema representativo do desenvolvimento de frutos de café. (A) Corte transversal de fruto maduro de café; (B) Fases do desenvolvimento de frutos de café ao longo do processo de maturação (Fonte: DE CASTRO e MARRACCINI, 2006).
Desta forma, é comum a presença de frutos de café em diversos estágios de maturação no momento da colheita. Em muitas regiões de Minas Gerais, estado responsável por aproximadamente 50% da produção de café em 2009 (BRASIL, 2010) e de outros estados produtores de café, o relevo acidentado impossibilita a colheita mecanizada e, portanto, toda a colheita é feita manualmente. Isso aumenta os custos de produção e, em função desta desuniformidade de maturação dos frutos, muitos agricultores optam por realizar uma colheita mais tardia utilizando a técnica da derriça total sobre pano. Este procedimento reduz drasticamente a qualidade do café pelo fato de os grãos permanecerem mais tempo no campo, expostos ao ataque de insetos e patógenos e às intempéries, além do contato dos grãos com o solo no momento da colheita, alterando de forma negativa as características organolépticas da bebida do café, depreciando seu valor no mercado internacional.
Embora de interesse comercial, a obtenção do sincronismo da maturação de frutos de café de uma mesma floração é uma característica agronômica difícil de ser obtida pelo
A
B
Pe
ri
ca
rpo
Exocarpo Mesocarpo Endocarpo
Endosperma (grão)
Perisperma (fase maternal)
Desenvolvimento do endosperma
Endosperma
(perisperma = camada prateada)
Camadaprateada
Divisão celular / Alongamento do
perisperma
Divisão celular do endosperma
Alongamento celular do endosperma
Maturação do pericarpo
melhoramento genético tradicional em função do desconhecimento das vias metabólicas que regulam e coordenam este complexo processo.
Analisando-se a maturação de frutos, verificamos que é um processo complexo, envolvendo diversas vias metabólicas que resultam não somente da alteração de pigmentação e da consistência dos frutos, mas também da modificação do metabolismo de açúcares, ácidos orgânicos, proteínas e de compostos oriundos do metabolismo secundário que influenciam diretamente nas características organolépticas do café (ROGERS et al., 1999; DE CASTRO e MARRACCINI, 2006).
Dentre as modificações nas características físico-químicas que os frutos de café passam durante o seu amadurecimento, merece destaque a alteração da rigidez e da composição da parede celular. Estes parâmetros ocorrem devido à ação de enzimas específicas que hidrolisam os componentes da parede celular dos frutos de café. Apesar das informações a respeito da distribuição espaço-temporal da ação das diversas enzimas que atuam sobre a parede celular e da diversidade de composição da parede celular entre as diversas espécies, especial atenção tem sido dada a culturas-modelo como, por exemplo, tomate (BRUMMELL e HARPSTER, 2001) que tem sido referência para as demais espécies. Investimentos em pesquisa têm sido direcionados para ampliar os conhecimentos sobre o processo de maturação de frutos devido à sua importância comercial na fase de pós-colheita e processamento. Neste sentido, a biologia molecular tem contribuído de forma significativa para ampliar os conhecimentos básicos e aplicados da ação espaço-temporal das enzimas que atuam na parede celular de frutos em maturação, possibilitando não somente a identificação de genes envolvidos neste processo, mas também possibilitando um maior nível de controle do processo per se.
Diversos genes envolvidos na síntese de enzimas que atuam diretamente na maturação de frutos têm sido estudados nas mais diversas culturas, dentre eles, as -galactosidases.
Smith e Gross (2000) identificaram uma família multigênica composta por sete genes da -galactosidase em tomate, os quais se expressavam distintamente em diversas fases do desenvolvimento da planta e dos frutos. Smith et al. (2002) estudaram a supressão da
expressão de um membro da família da -galactosidase em tomate (TBG4) utilizando a técnica do RNA antisenso. Eles observaram que o nível de supressão era variável entre as plantas de tomateiro regeneradas e que uma linhagem de tomateiro geneticamente modificado apresentou firmeza do fruto transgênico 40% maior quando comparado com o controle. Brummell e Harpster (2001) relacionaram uma série destas enzimas e suas atividades em
frutos de tomate, indicando que as -galactosidases, juntamente com as expansinas, seriam das primeiras enzimas a serem expressas durante a maturação do tomate, restringindo e/ou controlando a expressão das demais enzimas envolvidas neste processo.
Embora haja informações sobre a atividade das -galactosidases na maturação de frutos de café (GOLDEN et al., 1993; SALMONA et al., 2008; JOËT et al., 2009), existe uma carência de informações a respeito de sua participação no sincronismo do processo de maturação destes frutos.
1.6. AS -GALACTOSIDASES
As -galactosidases (EC 3.2.1.23) constituem um grupo de enzimas que pertencem à família das glicosil-hidrolases e são caracterizadas por hidrolisarem resíduos terminais não-redutores de resíduos de -D-galactosil a partir de -D-galactosídeos (ESTEBAN et al., 2005). Sua capacidade de liberar galactose de galactolipídeos, de glicoproteínas e de componentes da parede celular durante os processos de expansão celular, senescência e amadurecimento de frutos tem sido identificada na literatura.
As -galactosidases são largamente distribuídas em diversos reinos, presentes em bactérias, fungos, plantas e, inclusive, animais. Em bactérias, especificamente em Escherichia
coli, têm sido amplamente utilizadas como ferramenta da biologia molecular para a seleção de colônias bacterianas transformadas com o fragmento de DNA de interesse como, por exemplo, nos vetores pGEM (Promega) e nos vetores TOPO (Invitrogen) que utilizam sistema de seleção blue-white screening que, por sua vez, está relacionado com a expressão da
somente informações moleculares, mas também métodos de análise quantitativo e qualitativo da atividade da -galactosidase em E. coli.
No campo da engenharia genética de plantas, na fase inicial das pesquisas visando a produção de plantas geneticamente modificadas, o gene da -galactosidase de E. coli foi utilizado como gene marcador heterólogo para identificação de eventos de transformação em girassol e em fumo (HELMER et al., 1984) mas, em função da disponibilidade de outros genes marcadores mais eficientes, seu uso no meio científico foi pouco difundido.
A atividade enzimática da -galactosidase foi identificada em diversas plantas superiores como, por exemplo, tomate (SMITH et al., 1998; SMITH et al., 2002), manga (PRASANNA et al., 2005) e, inclusive, café (GOLDEN et al., 1993). Maiores detalhes
relacionados com as -galactosidases em plantas serão apresentados no capítulo II desta tese
(Caracterização molecular e funcional do gene da -galactosidase em C. arabica).
1.7. CARACTERIZAÇÃO ESTRUTURAL DA PAREDE CELULAR VEGETAL
As técnicas de microscopia possibilitaram uma exploração da estrutura da parede celular, principalmente a microscopia eletrônica. No entanto, estas abordagens elucidam apenas a morfologia da parede celular e não a sua composição bioquímica. Com os avanços tecnológicos de análise bioquímica de alta resolução, principalmente com o uso do HPLC (Cromatografia Líquida de Alta Performance) e do GC-MS (Cromatografia Gasosa acoplada à Espectrometria de Massa), pode-se verificar que a estrutura da parede celular é complexa e apresenta um comportamento extremamente dinâmico, modificando-se nos diversos estádios de desenvolvimento das plantas como, por exemplo, na expansão celular, na abscisão de folhas e flores, bem como no amadurecimento de frutos (CAMPBELL e BRAAM, 1999).
Anatomicamente, nas células vegetais podem ser encontrados dois tipos de parede celular, a depender da função celular e do seu estádio de desenvolvimento: a parede celular primária e a parede celular secundária. Além desta classificação genérica, a análise da parede celular por microscopia eletrônica possibilitou a identificação de três domínios distintos: a lamela média, a parede celular primária e, quando presente, a parede celular secundária (APPEZZATO-DA-GLÓRIA e CARMELLO-GUERREIRO, 2006).
A lamela média é a primeira camada depositada na região do apoplasto na fase inicial de formação de uma célula, após a divisão celular. É uma camada extremamente fina composta, principalmente, por substâncias pécticas. Sua função primordial é a união de células adjacentes (CARPITA e McCANN, 2000; TAIZ e ZEIGER, 2009).
A parede celular primária é o segundo domínio a ser depositado no apoplasto, após a formação da lamela média. Está presente em todos os tipos celulares vegetais, sendo característico de células jovens e em crescimento, assim como em células metabolicamente ativas. Embora apresentem padrões similares nos diversos tipos celulares, sua ultra-estrutura é altamente variável e relacionada com a função da célula que reveste (CARPITA e McCANN, 2000). Em termos estruturais, na parede celular primária podem ser encontrados três domínios que interagem entre si, a saber: o primeiro domínio é formado por uma rede de celulose e hemicelulose; o segundo domínio, por uma mistura de polissacarídeos pécticos e um terceiro domínio formado por proteínas.
ultra-estrutural da parede secundária permite identificar a presença de três camadas: S1 (mais
externa), S2 (intermediária) e S3 (mais interna e próxima à plasmalema).
Diante desta estrutura particular, a parede celular secundária pode apresentar duas funções distintas: (i) função estrutural, conferindo resistência física às células e (ii) função de reserva, armazenando polissacarídeos na sua matriz, os quais poderão ser utilizados na mobilização de carboidratos para as sementes (BUCKERIDGE et al., 2000).
1.8. POLISSACARÍDEOS DA PAREDE CELULAR PRIMÁRIA DE PLANTAS
Bioquimicamente, a parede primária das células vegetais é composta por celulose, hemicelulose e pectina, embora também sejam encontradas proteínas dispersas na parede celular (CARPITA e McCANN, 2000; KERBAUY, 2008).
Apesar de fazerem parte da estrutura da parede celular, interagindo entre si na sua matriz, estes grupos de polissacarídeos podem ser individualizados com base nas características bioquímicas das moléculas que os constituem. Desta forma, cada fração pode ser separada das demais em função dos diferentes graus de solubilidade que apresentam (SILVA, 2006), conforme segue:
Extrator Solubilidade do polímero
Água Pectinas
Alcalino fraco Hemiceluloses fracamente ligadas
Alcalino forte Hemiceluloses fortemente ligadas
Insolúvel (resíduo) Celulose
Neste aspecto, a solubilidade é importante para distinguir os carboidratos da parede celular que são, relativamente, inertes quanto à sua reatividade química. Esta solubilidade dos polissacarídeos da parede celular é uma característica biológica e físico-química importante para as sementes, além de afetar os produtos primários do café (café torrado e moído e café solúvel), determinando o seu rendimento e, consequentemente, o custo de processamento.
No fruto maduro de café, os polissacarídeos compartimentados na espessa parede celular do endosperma, podem ser extraídos por meio de água quente, extrações em soluções de álcali ou tratamento com enzimas (BRADBURY, 2001 citado por SILVA, 2006) e a ordem para liberar esses polímeros da parede celular é arabinogalactano-galactomanano-celulose (SILVA, 2006).
polissacarídeos, porém compostos por diversos tipos de monossacarídeos, em uma organização estrutural complexa, formando diversas classes destes polímeros.
Diante deste cenário, segue um breve resumo de cada um dos componentes da parede celular primária das células vegetais.
Celulose
A celulose representa de 20% a 30% da parede celular primária e, aproximadamente, 40% da parede celular secundária, sendo considerado o composto orgânico mais abundante no mundo (McNEIL et al., 1984). Estruturalmente, a celulose é um polissacarídeo composto por
um único tipo de monossacarídeo, a -D-glucopiranose (glicose), ligadas pelos carbonos
adjacentes 1 e 4 por ligação glicosídica tipo -D-(14). Esta ligação promove uma inversão de uma glicose a cada dois monossacarídeos (ASPINALL, 1980 citado por AMARAL, 2005), de tal forma que cada molécula de glicose fica disposta em uma rotação de 180º em relação à molécula de glicose adjacente. Esta organização forma a unidade básica da celulose, um dissacarídeo denominado celobiose que se repete inúmeras vezes.
A enzima celulose sintetase, localizada na membrana plasmática das células vegetais em complexos protéicos denominados rosetas, é a responsável pelo alongamento das cadeias de celulose que são depositadas no apoplasto, formando microfibrilas que nada mais são do que cadeias lineares de unidades de celubiose repetitivas, dispostas de forma antiparalela (CARPITA e McCANN, 2000; BUCKERIDGE e CAVALARI, 2008).
Em função da disposição das hidroxilas das moléculas de glicose, elas formam um grande número de ligações de pontes de hidrogênio, as quais competem com a água de hidratação. Com isso, na formação das microfibrilas, apenas um número reduzido de água permanece no cristal e, consequentemente, forma uma estrutura cristalina compacta e desidratada (KERBAUY, 2008).
Hemiceluloses
As hemiceluloses são uma classe formada por polissacarídeos altamente maleáveis e heterogêneos que interagem firmemente com as microfibrilas de celulose por intermédio de pontes de hidrogênio e, em função desta interação, são mais solúveis em solventes alcalinos
fortes (2 – 4 M NaOH) (CARPITA e McCANN, 2000; TAIZ e ZEIGER, 2009). Além disto,
Vale destacar que a terminologia “hemicelulose” induz a um equívoco, uma vez que
nos impele a imaginarmos a sua estrutura semelhante à celulose, o que não condiz com a realidade. Enquanto as celuloses são estruturalmente simples, bioquimicamente homogêneas e dispostas linearmente formando os microtúbulos, as hemiceluloses são estrutural e bioquimicamente heterogêneas. Este termo foi, inicialmente, proposto para descrever os compostos da parede celular solúveis em solventes alcalinos.
As hemiceluloses são formadas por uma cadeia principal linear de xilose, manose ou
glicose, unidos por ligações do tipo (1,4), denominado eixo osídico, podendo apresentar ramificações com diversos tipos de monossacarídeos. Quando este eixo central é formado por glicose, recebe a denominação glucano como, por exemplo, os xiloglucanos (uma das principais hemiceluloses encontradas na natureza) possuem o eixo formado por glicose. Quando o eixo central é formado por xilose, recebe a denominação xilano como, por exemplo, os glucuronoarabinoxilanos (outra classe hemicelulósica) que possuem o eixo principal formado por xilose.
Existem diversas classes de hemiceluloses, dentre as quais se destacam os xiloglucanos (XG) (mais abundantes em dicotiledôneas), arabinoxilanos e os galactomananos (GM), dentre outros (COSGROVE, 2005).
Pectinas
As pectinas, por sua vez, formam uma mistura de polissacarídeos ricos em monossacarídeos ácidos (p.ex. ácido galacturônico) e monossacarídeos neutros (ramnose, galactose e arabinose), além de serem bastante heterogêneas, complexas e ramificadas. Diferentemente das hemiceluloses, as pectinas não interagem fortemente com as microfibrilas de celulose e, portanto, são mais solúveis que as hemicelulose, podendo ser extraídas com água quente ou quelantes de cálcio (TAIZ e ZEIGER, 2009). Vale destacar que as pectinas têm sido classificadas como materiais que podem ser extraídos da parede celular por agentes quelantes de cálcio, EDTA (ácido etilenodiamino tetra-acético) e EGTA (ácido etilenoglicol tetra-acético), influenciando diretamente na porosidade da parede, na adesão célula-a-célula na lamela média, além de fornecer moléculas de reconhecimento para interação com microorganismos fitopatogênicos, simbiontes e de insetos (CARPITA e McCANN, 2000).
Estes compostos são polissacarídeos ácidos, com uma cadeia central formada por
esta cadeia apresenta não somente unidades de ácido galacturônico, mas também unidades de ramnose, intercalando as unidades de ácido galacturônico, sendo que as ramificações (com galactanos ou arabinanos) ocorrem nas unidades de ramnose.
Dentro deste contexto, quantitativamente os três principais grupos de polímeros que compõem as pectinas são:
1 – Homogalacturonanos (HG): são pectinas não ramificadas formadas somente por
ácido galacturônico com ligações do tipo (1,4);
2 – Ramnogalacturonanos I (RG I): são pectinas ramificadas com cadeia principal apresentando unidades de ramnose, intercaladas com ácido galacturônico, cujas ramificações ocorrem nas ramnoses. As ramificações com galactanos podem ser com ligações do tipo
(1,4) e/ou ligações do tipo 1,3-1,6. As ramificações com arabinanos, por sua vez, são do tipo
(1,5) (BUCKERIDGE et al., 2008).
3 – Ramnogalacturonanos II (RG II): normalmente ocorrem em pequena quantidade na parede celular vegetal, com algumas características das pectinas, mas são altamente complexas e não entraremos em detalhes sobre a sua constituição.
Este breve resumo ilustra a complexidade estrutural dos polissacarídeos encontrados na parede primária de células vegetais, mas uma revisão bibliográfica específica e mais detalhada sobre a composição de carboidratos identificados em frutos de café é apresentada no capítulo I desta tese (Caracterização dos carboidratos da parede primária de frutos de
Coffea arabica em diferentes fases de desenvolvimento), juntamente com uma análise e discussão de sua composição durante diferentes estádios de sua maturação.
1.9. SILENCIAMENTO GÊNICO POR RNA INTERFERENTE
– double stranded RNA) que, por sua vez, é digerida por um complexo protéico chamado DICER (HAMILTON e BAULCOMBE, 1999; BERSTEIN et al., 2001). As fitas de siRNAs são, posteriormente, desenoveladas e uma fita é incorporada no complexo RISC
(RNA-induced silencing complex), o qual guia este complexo para um mRNA de sequência complementar, clivando-a e induzindo o silenciamento gênico (MEISTER e TUSCHL, 2004; BAULCOMBE, 2005).
O uso da técnica do RNAi para a descoberta e validação da função de determinado gene apresenta vantagens comparativamente com outras abordagens utilizadas com a mesma finalidade, mas precauções devem ser tomadas, uma vez que também existem desvantagens. Small (2007) faz um levantamento dos pontos positivos e negativos da estratégia do RNAi, dos quais destacamos os principais aspectos:
Vantagens do uso da técnica do RNAi:
1. RNAi é sequência-específica, possibilitando focar, especificamente, o
silenciamento do gene-alvo;
2. RNAi é dominante e, por esta razão, possibilita a identificação de mutantes na geração T1;
3. RNAi pode ser tecido-específico;
4. RNAi pode ser utilizado para regular a expressão de genes que compartilham homologia na sequência genômica alvo, facilitando o estudo de famílias multigênicas;
5. RNAi pode induzir um silenciamento total ou parcial da expressão gênica, produzindo diversos fenótipos mutantes, o que é crucial nos estudos de genes letais;
6. RNAi pode ser utilizado para o silenciamento de genes em genoma poliplóides (LAWRENCE e PIKAARD, 2003; TRAVELLA et al., 2006; FU et al., 2007); 7. Silenciamento por RNAi é estável, podendo seus efeitos serem avaliados na
progênie (CARTHEW, 2001);
8. RNAi induz uma resposta sistêmica, induzindo a degradação de mRNA
específicos em diversos órgãos/tecidos/células do organismo (HIMBER et al., 2003).
Desvantagens do uso da técnica do RNAi:
pelo fato de apresentarem homologia com os siRNAs (FEDOROV et al., 2006; KULKARNI et al., 2006);
2. Reduzida penetrância do silenciamento do RNA na progênie (WANG et al., 2005). No entanto, Carthew (2001) e Stoutjesdijk et al. (2002) observaram exatamente o oposto, com um estável silenciamento gênico induzido por RNAi em diversas gerações da progênie. Outro exemplo foi observado por Bonfin et al. (2007) que produziram plantas de Phaseolus vulgaris resistentes ao BGMV (Bean golden
mosaic virus) com o silenciamento do gene viral rep, envolvido na replicação do genoma viral. Estes pesquisadores acompanharam o nível de resistência da linhagem 5.1 por diversas gerações e observaram a manutenção desta resistência induzida por RNAi. No entanto, esta controvérsia requer maiores investigações por parte dos pesquisadores que atuam nesta área.
No caso do gene das -galactosidases, a utilização do silenciamento gênico com o uso da técnica do RNAi apresentou-se como a abordagem mais indicada por duas razões:
(i) Como as -galactosidases pertencem a uma família multigênica em diversas espécies vegetais (SMITH e GROSS, 2000; TRAINOTTI et al., 2001; TRIANTAFILLIDOU e GEORGATSOS, 2001; AHN et al., 2007;), poderia ser também o fosse na cultura do café (esta suspeita foi confirmada em C. arabica– vide capítulo II desta tese) e;
(ii) Como C. arabica é uma espécie alotetraplóide (FAZUOLI et al., 1999), a sua base genética contém múltiplas cópias de um mesmo gene.
Desta forma, a utilização da técnica do RNAi para o silenciamento (parcial ou total) do gene da -galactosidase aumentaria a probabilidade de redução da expressão das múltiplas cópias deste gene na planta de café, possibilitando uma avaliação do seu efeito no sincronismo da maturação de frutos desta cultura.
Maiores detalhes sobre a técnica do RNAi são abordados no capítulo III desta tese.
JUSTIFICATIVAS
Entretanto, pelo fato de o café ser uma cultura perene, os longos ciclos do melhoramento tradicional dificultam a incorporação de características de interesse num reduzido espaço de tempo. Além disso, muitas características não estão disponíveis no genoma da espécie, são multigênicas e/ou são reguladas por estímulos ambientais difíceis de serem controladas em condições de campo.
Neste sentido, a biologia molecular tem permitido a obtenção de informações mais precisas sobre os genes envolvidos nas diversas vias bioquímicas de interesse, inclusive na maturação de frutos. Estes conhecimentos poderão ser utilizados para elaborar mecanismos de interferência no processo de forma controlada e num menor espaço de tempo, comparativamente com os métodos tradicionais.
Sabe-se que a maturação de frutos é um processo complexo, multigênico, envolvendo várias reações bioquímicas sequenciais controladas espaço-temporalmente, além de ser influenciada por diversos fatores endógenos (p.ex. hormônios) e exógenos (disponibilidade de água, dentre outros). Apesar de sua enorme complexidade, a importância para a pós-colheita de frutos e seu impacto na balança comercial tem justificado investimentos em pesquisas que visam elucidar os processos relacionados com a maturação de frutos.
Genes de várias enzimas responsáveis pela hidrólise de componentes da parede celular de frutos de diversas espécies já foram clonados e estudados, inclusive em café (MARRACCINI et al., 2001; MARRACCINI et al., 2005). Apesar disso, há uma carência de informações moleculares a respeito da expressão das -galactosidases e de sua participação na maturação de frutos de café.
Em função das -galactosidases comporem uma família multigênica em outras espécies vegetais e devido à falta de informações em Coffea sp. no momento da elaboração desta tese, a utilização da técnica do RNA interferente (RNAi) apresentou-se como a mais promissora pelo fato de possibilitar a supressão, parcial ou total, de todos os membros desta provável família multigênica em cafeeiro.
Em função dessa carência de informações sobre as -galactosidases expressas em café e da sua possível participação no processo de maturação dos frutos do cafeeiro, um grupo de pesquisa da EMBRAPA Recursos Genéticos e Biotecnologia (Brasília/DF), liderado pelo Dr. Francisco J. L. Aragão, iniciou os estudos para a clonagem e silenciamento deste gene, utilizado a técnica do RNAi, em café.

![Figura 3. Produção dos principais países produtores de café na safra de 2010. [Fonte: Brasil (2010)]](https://thumb-eu.123doks.com/thumbv2/123dok_br/18209905.339377/24.893.235.716.444.760/figura-produção-principais-países-produtores-café-fonte-brasil.webp)