Contents lists available atScienceDirect
European Journal of Pharmacology
journal homepage:www.elsevier.com/locate/ejphar
HIV antiretroviral drug Efavirenz induces anxiety-like and depression-like
behavior in rats: evaluation of neurotransmitter alterations in the striatum
Giuliana Ignácio Teixeira Cavalcante
a, Adriano José Maia Chaves Filho
a, Maria Isabel Linhares
a,
Camila Nayane de Carvalho Lima
a, Edith Teles Venâncio
a,b, Emiliano Ricardo
Vasconcelos Rios
a,b, Francisca Cléa Florenço de Souza
a, Silvânia Maria Mendes Vasconcelos
a,
Danielle Macêdo
a, Marta Maria de França Fonteles
a,b,⁎aDrug Research and Development Center, Department of Physiology and Pharmacology, Faculty of Medicine, Federal University of Ceará, Fortaleza, CE,
Brazil
bPharmacy Department, Faculty of Dentistry, Nursing and Pharmacy, Federal University of Ceará, Fortaleza, CE, Brazil
A R T I C L E I N F O
Keywords:
Antiretroviral drugs Efavirenz
Neuropsychiatric adverse events Monoamines
Neurotransmitters Striatum
A B S T R A C T
Efavirenz (EFV) is an effective antiretroviral drug with a favorable pharmacokinetic profile and widely used in combination regimens to treat HIV infection. However, there are major concerns about the safety of this drug. Patients treated with EFV often experience neuropsychiatric adverse effects, which frequently lead to switching to alternative EFV-free regimens. The mechanisms involved in the central action of EFV are intrinsically unclear. Thus, this study aimed to investigate the effects of acute and subchronic (2 weeks) EFV administration in a series of behavioral tests for anxiety-like and depression-like behavior in healthy rats. We also evaluated the effect of EFV treatment in striatal concentrations of monoamine neurotransmitters (serotonin, dopamine and noradrenaline) and their metabolites and the amino acid neurotransmitters glutamate and GABA. Our results showed that acute treatment with EFV induced an anxiogenic-like effect, while sub-chronic treatment induced both anxiogenic-like and depressive-like behavior which was dose related.. Additionally, EFV treatment caused marked alterations in the striatal concentrations of monoamines and their metabolites (and turnover rates) and the amino acid neurotransmitters glutamate and GABA. These changes were influenced by treatment duration and dose. Thesefindings add more evidence about the neuropsychiatric adverse effects of EFV and propose potential new mechanisms for the toxic action of this drug in the central nervous system.
1. Introduction
Efavirenz (EFV), belonging to antiretroviral class of non-nucleoside reverse transcriptase inhibitors (NNRTI), is a potent anti-HIV drug with a long half-life which readily crosses the blood-brain barrier (Arribas, 2003; von Giesen et al., 2002). For these reasons, EFV is a key component of the most frequently prescribed treatment combinations for HIV infection (Hammer et al., 2008; Thompson et al., 2012). Despite this, its use has often been associated with significant adverse neuropsychiatric effects, such as insomnia, dizziness, nightmares, and affective states, e.g., anxiety, depression and, sometimes, suicidal ideation, mainly during thefirst weeks of therapy (Meier et al., 2015; Mothapo et al., 2015).
The mechanisms for the central toxicity of EFV remain poorly understood. Studies addressing this question could lead to improve-ment in antiretroviral therapy, given the impact that EFV
neuropsy-chiatric effects have on adherence to therapy, the most important factor for good prognosis of HIV-positive patients (Ammassari et al., 2001; Bezabhe et al., 2016).
Therefore, the present study aimed to investigate the effects of acute and subchronic (2 weeks) EFV treatment on behavioral tests for anxiety-like and depression-like behavior in healthy adult rats. Furthermore, we investigated the effects of EFV on the striatal concentrations of monoamines (and their metabolites) and amino acid neurotransmitters glutamate and gamma-aminobutyric acid (GABA). Given the importance of these neurotransmitters for the regulation of mood, motivation and reactivity to stress, and their involvement in the pathophysiology of neuropsychiatric disorders, especially affective disorders (Hamon and Blier, 2013; Möhler, 2012), we hypothesize that changes in these neurotransmitter systems are potentially involved in the mechanisms of EFV neuropsychiatric effects.
http://dx.doi.org/10.1016/j.ejphar.2017.02.009
Received 18 November 2016; Received in revised form 7 February 2017; Accepted 7 February 2017
⁎Correspondence to: Department of Physiology and Pharmacology, Federal University of Ceará, Rua Cel.Cel. Nunes de Melo 1127, CEP: 60430-270, Fortaleza, Ceará, Brazil.
E-mail address:[email protected](M.M. de França Fonteles).
Available online 08 February 2017
0014-2999/ © 2017 Elsevier B.V. All rights reserved.
2. Material and methods
2.1. Animals
Male adult Wistar rats, weighting 200–250 g, were used for behavioral and neurochemical tests. The animals were housed two per cage in standard polycarbonate rat cages (42×20.5×20 cm) and standard environmental conditions (22 ± 1 °C; humidity 60 ± 5%; reversed 12-h light/dark cycle with lights on at 19:00) with access to food and water ad libitum. All experimental procedures were con-ducted between 8:00 and 14:00 h and were carried out in accordance with the NIH Guide for the Care and Use of Laboratory Animals (NIH, 2011) and the Brazilian College of Animal Experimentation (COBEA). This research protocol was approved by the local ethical committee of Federal University of Ceará, with Protocol Number 09/2011.
2.2. Drugs
Stocrin®Efavirenz (EFV), (4 S)-6-chloro-4-(2-cyclopropylethynyl)-4-(trifluoromethyl)-2,4-dihydro-1H-3,1-benzoxazin-2-one (Merck Sharp & Dohme Corp., NJ, USA), imipramine (IMI; Sigma and Aldrich Corp., St Louis, USA) and diazepam (DZP, Sigma and Aldrich Corp., St Louis, USA) were used. EFV was dissolved in distilled water slightly warmed with 1% Tween-80. IMI and DZP was dissolved in 0,9% saline. The drugs were made up freshly for the study. All other chemicals used were of analytical grade.
2.3. Experimental design
Rats were randomly assigned into groups with 8–10 animals each. For the acute treatment, EFV (25 or 50 mg/kg) was administered orally by gavage (v.o.) and 60 min later, behavioral tests were performed. For the sub-chronic treatment, EFV (25 or 50 mg/kg, v.o.) was adminis-tered for 14 days, and 60 min after the last administration, the behavioral tests were performed. The controls received distilled water with 1% Tween-80 (vehicle) (v.o), and followed the same protocols as the EFV treated groups. Further, as standard drugs, we conducted groups intraperitoneally treated with IMI (10 mg/kg) and DZP (2 mg/ kg) 30 min before the behavioral tests. The weight of animals was monitored daily before each drug administration. The behavioral tests were organized from the least to the most stressful: openfield test, elevated plus maze test and forced swimming test respectively (Paylor et al., 2006), and two experienced raters who were blind to the treatment group independently assessed rodent behavior. To avoid the possible confounding effect of behavioral tests on the neurochem-ical parameters, different groups of animals were assigned to behavior-al assessment and biochemicbehavior-al anbehavior-alysis. Animbehavior-als intended for neuro-chemical evaluation were killed by decapitation at the end of the treatment period and brains were rapidly removed from the skull placed in ice cold. After this, the striatal tissue was micro-surgically dissected accordingly the procedure described byChiu et al. (2007) and immediately stored at−70 °C.
The rat dose of 50 mg/kg of EFV corresponds to the oral ther-apeutic human dose of 600 mg per day or approximately 10 mg/kg (Dhoro et al., 2015; Gatanaga et al., 2007). The dose calculation was based on the human to rat body surface area normalization method (Mastan and Kumar, 2009; Reagan-Shaw et al., 2008). In addition, we included the dose of 25 mg/kg of EFV, since several studies applied lower doses of EFV ranging from 3 to 30 mg/kg in their experimental protocols (Gatch et al., 2013; O'Mahony et al., 2005; Romão et al., 2011). The IMI and DZP dose were selected from previous literature evaluating respectively the antidepressant-like and anxiolytic-like effects of these drugs in rodents (Berrocoso et al., 2012; Santos et al., 2017).
2.4. Behavioral determinations
2.4.1. Openfield test
Locomotor activity and exploratory behavior were measured in an openfield area, which consist of a square plastic board (40×40 cm, walls 40 cm high) divided into nine squares, based on a previously described model (Archer, 1973). The rat was placed into a corner of the area and observed for 5 min under conditions of red light (28 lx in the center). Each rat was tested once and between two rats, the apparatus was cleaned with 20% ethanol solution. The parameters analyzed during the test were: the number of animal crossings with four legs (spontaneous locomotion), the number of rearings (exploratory vior) and the number of self-cleaning behaviors (self-grooming beha-vior).
2.4.2. Elevated plus maze test
The elevated plus maze apparatus, described by Lister (1987), consisted of two open arms (50×10 cm, walls 40 cm high) and two closed arms (50×10 cm, walls 40 cm high) connected by a central square (10×10 cm), and elevated to a height of 50 cm. The rat was placed in the apparatus center facing one of the open arms and allowed to explore for 5 min under conditions of red light (28 lx in the center). The rat was considered to have entered an arm when all four limbs were inside the arm. Each rat was tested once and between two rats, the apparatus was cleaned with 20% ethanol solution. The behavioral measures observed during the test were the number of entries in open and closed arms and the time spent in each of these sections. These data were used to calculate the number of total entries, the percentage of open entries (i.e., open entries/total entries×100) and the percen-tage of time in open arms (i.e., time in open arms/total duration×100).
2.4.3. Forced swimming test
The forced swimming test was performed over 2 days as previously described (Porsolt et al., 1977). On thefirst day, the pre-swimming session, rats were individually placed for 15 min into an acrylic cylinder (40 cm height, 22 cm diameter) containing 20 cm of water maintained at 23 ± 1 °C. Twenty-four hs later, rats underwent the test session. They were again placed in cylindersfilled with water, and the duration of immobility was recorded for 5 min. The immobility time was defined as the time spent by the rat in the absence of active, escape-oriented behaviors, such as swimming, jumping, rearing, sniff -ing, or div-ing, making only those movements necessary to keep its head above the water.
2.5. Neurochemical determinations
2.5.1. Determination of the levels of monoamines and their metabolites
The striatal levels of dopamine (DA), noradrenaline (NE), serotonin (5-HT) and their non-conjugated metabolites 3,4-hydroxyphenylacetic acid (DOPAC), homovanilic acid (HVA), and 5-hydroxy-indoleacetic acid (5-HIAA) were assayed by reverse-phase high-performance liquid chromatography (HPLC) with electrochemical detection. A C18 reverse phase column (Shim-pack, CLC-ODS 150×4.6 mm; Shimadzu, Kyoto, Japan), an amperometric detector (Shimadzu,L-ECD-6A) and a liquid
ng/g tissue wet weight.
2.5.2. Determination of the levels of glutamate and GABA
Striatal levels of amino acids glutamate and GABA were determined by HPLC with afluorimetric detector (330–450 nm) (Model RF-535 Shimadzu, Japan) coupled to an integrator. The C18 chromatographic column was used (CLC-ODS 150×4.6 nm, Shimadzu, Kyoto, Japan) with flow rate of 1 ml/min. The mobile phase A consisted of 50 mM NaH2PO4 in 20% methanol adjusted to pH 5.5. The mobile phase B consisted of 100% methanol. Briefly, the brain tissues were weighed and homogenized in 10% (w / v) of 0.1 M perchloric acid. After centrifugation (15.000 rpm/15 min), supernatants were removed,fi l-tered through the membrane (Millipore, Brazil, 0.22 µm) and com-bined with a solution of pre-column derivatization, to obtainfl uores-cence in a ratio of 1:1. The volume was completed to 2.25 ml with borate buffer, pH 9.3. 20-ul of this final solution was analyzed by chromatography. The peak areas of the external standards were used to quantify the sample peaks. The values were expressed as µmol / g tissue wet weight.
2.6. Statistical analysis
The results of behavioral and neurochemical tests are expressed as the means ± S.E.M. (standard errors of the mean) and were analyzed with a regular two-way analysis of variance (ANOVA) followed by a post-hoc Tukey's test. The factors were“efavirenz treatment”(Vehicle, EFV25 or EFV50) and “duration of treatment”(acute or subchronic treatment). The significance level was set at P < 0.05. The statistical program used was GraphPad Prism 6.0 Version for Windows, GraphPad Software (San Diego, CA, USA).
3. Results
3.1. Effect of acute and subchronic efavirenz treatment in the open field test
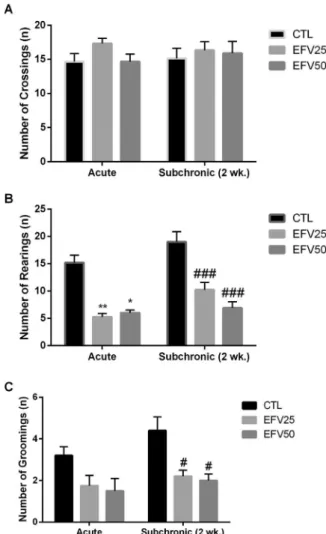
As shown inFig. 1A, regarding the number of crossings, we did not detect any significant difference between the groups analyzed (P > 0.05).
In relation to the number of rearings (Fig. 1B), two-way ANOVA test revealed a significant main effect of both factors: “efavirenz treatment”[F (2, 42) =44.70, P < 0.0001] and“duration of treatment” [F (1, 42) =10.31, P=0.0025] without significant interaction between them. Post hoc test showed a significant decrease in the number of rearings in the EFV25-Acute (P < 0.01) and EFV50-Acute (P < 0.05) groups compared to Vehicle-Acute group. Also, a significant decrease in this parameter was observed in subchronic EFV-treated animals, EFV25-subchronic (P < 0.001) and EFV50-subchronic (P < 0.0001) groups, compared to its respective controls. No significant difference was identified between groups of different time protocols.
The analysis of the number of groomings showed only a significant main effect of “efavirenz treatment” singly [F (2, 42) =10.90, P=0.0002] (Fig. 1C). In addition, post hoc test revealed a significant decrease in the number of groomings in the subchronic EFV-treated groups at both doses studied compared to Vehicle-subchronic group (P < 0.05).
Acute DZP (2 mg/kg), as expected, significantly increased the number of crossings (24.56 ± 1.492, n=10) and the number of rearings (33.57 ± 1.232, n=10) compared to vehicle-control groups of both schedule protocols (P < 0,001), which is compatible with the induction of an anxiolytic-like effect (data not shown).
3.2. Effect of acute and subchronic efavirenz treatment in the elevated plus maze test
As shown inFig. 2A, our results showed a significant main effect of
“efavirenz treatment”[F (2, 42) =13.56, P < 0.0001] in the time spent in open arms (% of Time in Open Arms/Total Duration), without significant interaction between the factors [F (2, 42) =1.131, P=0.3322]. Post hoc test revealed a significant decrease in this parameter in the acute EFV-treated groups compared to Vehicle-Acute group (P < 0.01). Additionally, a significant decrease in the time spent in open arms was observed in the EFV50-subchronic group compared to the Vehicle-subchronic group (P < 0.05).
Regarding the percentage of Open entries/Total entries (%), ANOVA test revealed a significant main effect of the “duration of treatment”[F (1, 42) =4.808, P=0.0339] and the interaction between the factors [F (2, 42) =3.825, P=0.0298]. Post hoc analysis detected a significant decrease in this parameter in EFV50-Subchronic group compared to Vehicle-Subchronic (P < 0.05) and EFV50-Acute groups (P < 0.05) (Fig. 2B). In relation to total number of entries, the analysis revealed no significant differences between the groups (Fig. 2C).
Finally, acute DZP (2 mg/kg) promoted a significant increase in % of Time in Open Arms/Total Duration (55.7 ± 2.367, n=10) and % of Open entries/Total entries (%) (68.9902 ± 5.675, n=10) in relation to control groups of both schedule protocols (P < 0.05), without affect the total number of entries (12.875 ± 1.423, n=10) (p > 0.05) (data not shown). Thus, confirming the induction a clear anxiolytic-like effect. Fig. 1.Effects of acute and subchronic EFV treatment (EFV25: 25 mg/kg; and EFV50: 50 mg/kg) on the number of crossings (A), number of rearings (B) and number of
groomings (C) in rats subjected to the openfield test. Bars represent ± mean S.E.M. of 8
animals/group.* P
≤0.05 vs CTL-Acute,**P
≤0.01 vs CTL-Acute;#P
≤0.05 vs
CTL-Subchronic,##P
≤0.01 vs CTL-Subchronic. All comparisons were performed according to
3.3. Effect of acute and subchronic efavirenz treatment in the forced swimming test
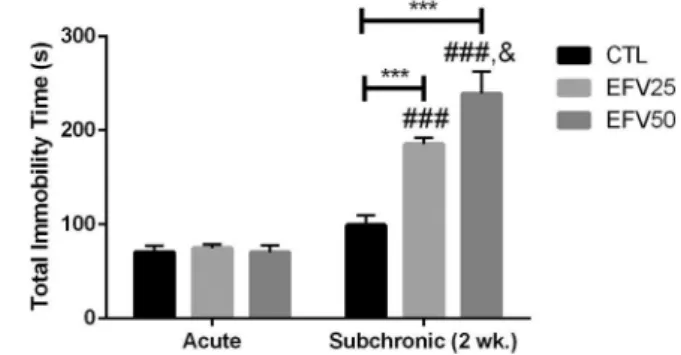
As shown in Fig. 3, we detected a significant main effect of “efavirenz treatment”[F (2, 42) =17,96, P < 0.0001] and“duration of treatment”[F (1, 42) =114.9, P < 0.0001] in the total immobility time, with a significant interaction between these factors [F (2, 42) =18.17, P < 0.0001]. Post hoc test revealed a significant increase in the total immobility time in the subchronic EFV-treated groups, EFV25-Subchronic (P < 0.001) and EFV50-EFV25-Subchronic (P < 0.0001), compared to Vehicle-Subchronic group. EFV50-Subchronic group also showed an increase in immobility time compared to EFV25-Subchronic group (P < 0.5). Additionally, an increase in immobility time was noted in subchronic EFV-treated groups compared to acute EFV-treated ones: EFV25- Subchronic versus (vs) EFV25-Acute (P < 0.0001) and EFV50-Subchronic vs EFV50-Acute (P < 0.0001).
As expected, acute IMI (10 mg/kg) significantly reduced the immobility time (37, 67 ± 9,58, n=10) when compared to acute (P < 0.05) and subchronic (P < 0.01) vehicle control groups, confirming its antidepressant-like effect (data not shown).
3.4. Effect of acute and subchronic efavirenz treatment in the striatal levels of monoamines and their metabolites
3.4.1. DA, DOPAC, HVA and DA turnover rate
As seen inFig. 4A, two-way ANOVA showed a significant main effect of “efavirenz treatment” [F (2, 42) =7.548, P=0.0016] and “duration of treatment”[F (1, 42) =43.63, P < 0.0001] in striatal DA levels with a significant interaction between these factors [F (2, 42) =23.64, P < 0.0001]. Post hoc test revealed a significant increase in DA levels in EFV25-Acute (P < 0.01) and EFV50-Acute (P < 0.0001) groups compared to acute controls. A significant increase in DA levels was also noted in EFV50-Acute group compared to EFV25-Acute group (P < 0.01). When compared the different duration protocols, we observed a significant decrease in striatal DA in subchronic EFV-treated groups compared to acute ones: EFV25-Subchronic vs EFV25-Acute (P < 0.05) and EFV50-Subchronic vs EFV50-Acute (P < 0.0001).
DA has two major metabolites: DOPAC, derived from monoamine oxidase (MAO) metabolism and HVA derived from catechol-O-methyl-transferase (COMT) metabolism (Konradi et al., 1992). Regarding DOPAC levels, we detected a significant main effect of “efavirenz treatment”[F (2, 42) =27.90, P < 0.0001] and“duration of treatment” [F (1, 42) =70.59, P < 0.0001], with significant interaction between these factors [F (2, 42) =24.63, P < 0.0001]. Post hoc analysis showed a significant increase in DOPAC levels in EFV50-Acute group related to EFV25-Acute (P < 0.01) and vehicle control groups (P < 0.001). Comparing different time protocols, a significant decrease in DOPAC levels was noted in Subchronic group compared to EFV50-Acute (P < 0.0001) and Subchronic group compared to EFV25-Acute (P < 0.05) (Fig. 4B).
In relation to HVA levels, statistical analysis revealed a significant main effect of“efavirenz treatment”[F (2, 42) =23.06, P < 0.0001] and “duration of treatment”[F (1, 42) =5.825, P=0.0202], with significant interaction between these factors [F (2, 42) =4.519, P=0.0167]. After post hoc test, it was evidenced a significant increase in HVA levels in EFV50-Acute group compared to Vehicle-Acute (P < 0.0001) and EFV25-Acute groups (P < 0.0001). No significant difference was iden-tified between subchronic EFV-treated groups and its respective controls (Fig. 4C).
Finally, regarding DA turnover rate, two-way ANOVA showed a significant main effect of the factors“efavirenz treatment”[F (2, 42) =11.77, P < 0.0001] and “duration of treatment”[F (1, 42) =8.344, P=0.0061] separately, and its interaction [F (1, 42) =8.344, P=0.0061]. Post hoc analysis revealed a significant decrease in DA turnover in EFV25-Acute group compared to acute Vehicle controls (P < 0.01). On the other hand, an increase in DA turnover was found in EFV50-Subchronic group compared to Vehicle-EFV50-Subchronic (P < 0.0001) and EFV25- Subchronic (P < 0.0001) groups. In addition, when compared different protocols, we observed a significant increase in DA turnover Fig. 2.Effects of acute and subchronic EFV treatment (EFV25: 25 mg/kg; and EFV50:
50 mg/kg) on the % of time in open arms (time in open arms/total duration×100) (A), % of open entries ( open entries/total entries×100) (B) and number of total entries (C) in rats subjected to elevated plus maze test . Bars represent ± mean S.E.M. of 8 animals/
group.**P
≤0.01 vs CTL- Acute,#P
≤0.05 vs CTL-Subchronic,&P
≤0.05 vs EFV50-Acute.
All comparisons were performed according to two-way ANOVA with Tukey's post hoc test. Abbreviations: CTL- Control, EFV- Efavirenz.
Fig. 3.Effects of acute and subchronic EFV treatment (EFV25: 25 mg/kg; and EFV50: 50 mg/kg) on the total immobility time in rats subjected to forced swimming test. Bars
represent ± mean S.E.M. of 8 animals/group. *** P
≤0.001 vs CTL-Subchronic; ###
P≤0.001 vs EFV25-Acute and EFV50-Acute; & P
≤0.05 vs EFV25-Subchronic. All
in EFV50-Subchronic compared to EFV50-Acute group (P < 0.001) (Fig. 4D).
3.4.2. 5-HT, 5-HIAA and 5-HT turnover rate
As shown inFig. 5A, the two-way ANOVA revealed a significant
main effect of“efavirenz treatment”[F (2, 42) =14.92, P < 0.0001] and “duration of treatment”[F (1, 42) =23.59, P < 0.0001] in striatal 5-HT levels, with a significant interaction between these factors [F (2, 42) =45.46, P < 0.0001]. Post hoc analysis showed a significant increase in 5-HT levels in EFV50-Acute group compared to Vehicle-Acute (P < Fig. 4.Effects of acute and subchronic EFV treatment (EFV25: 25 mg/kg; and EFV50: 50 mg/kg) on the levels of DA (A), DOPAC (B), HVA (C) and DA turnover rate (DOPAC + HVA/
DA) (D) in striatum. Bars represent ± mean S.E.M. of 8 animals/group.**P
≤0.01 vs CTL- Acute;***P
≤0.001 vs CTL- Acute or CTL-Subchronic;#P
≤0.05 vs EFV25-Acute;##P
≤0.01 vs
EFV25-Acute;###P
≤0.001 vs EFV25-Acute;& & &P
≤0.001 vs EFV50-Acute. All comparisons were performed according to two-way ANOVA with Tukey's post hoc test. Abbreviations:
CTL- Control, EFV- Efavirenz.
Fig. 5.Effects of acute and subchronic EFV treatment (EFV25: 25 mg/kg; and EFV50: 50 mg/kg) on the levels of 5-HT (A), 5-HIAA (B), 5-HT turnover rate (5-HIAA/5-HT) (C) and NE
(D) in striatum. Bars represent ± mean S.E.M. of 8 animals/group.**P
≤0.01 vs CTL- Acute or CTL-Subchronic;***P
≤0.001 vs CTL- Acute or CTL-Subchronic;#P
≤0.05 vs EFV25-Acute;
##P
≤0.01 vs EFV25-Acute;###P
≤0.001 vs EFV25-Acute;& & &P
≤0.001 vs EFV50-Acute. All comparisons were performed according to two-way ANOVA with Tukey's post hoc test.
0.05) and EFV25-Acute groups (P < 0.01). On the other hand, in subchronic protocol, a significant decrease in 5-HT levels was found in both EFV-treated groups compared to controls (P < 0.01). Additionally, when compared different duration protocols, we observed a decrease in 5-HT levels in EFV50-Subchronic group compared to EFV50-Acute group (P < 0.0001).
In relation to 5-HIAA levels, we detected a significant main effect of “efavirenz treatment”[F (2, 42) =18.44, P < 0.0001] and“duration of treatment”[F (1, 42) =27.19, P < 0.0001] in its striatal levels with a significant interaction between these factors [F (2, 42) =5.041, P=0.0109]. Post hoc test revealed a significant increase in 5-HIAA levels in EFV50-Acute group compared to Vehicle-Acute (P < 0.0001) and EFV25-Acute (P < 0.0001) groups. Additionally, a decrease in this metabolite was noted in Subchronic group compared to EFV50-Acute one (P < 0.0001) (Fig. 5B).
Regarding striatal 5-HT turnover rate, there was a significant interaction between the factors“efavirenz treatment”X “duration of treatment”[F (2, 42) =6.536, P=0.0034] with a main effect of both factors separately (P < 0.0001). Post hoc analysis showed a significant increase in 5-HT turnover rate in EFV25-Subchronic (P < 0.01) and EFV50-Subchronic (P < 0.0001) groups compared subchronic vehicle controls. When compared different protocols, we observed a significant increase in 5-HT turnover rate in EFV50-Subchronic group compared to EFV50-Acute group (P < 0.0001), and EFV25-Subchronic group compared to EFV25-Acute group (P < 0.05) (Fig. 5C).
3.4.3. NE levels
Statistical analysis revealed a significant interaction between “efa-virenz treatment”and“duration of treatment”[F (2, 42) =16.75, P < 0.0001] in striatal NE levels, although no significant main effect of these factors individually was detected. Post hoc test showed a significant increase in NE levels in EFV50-Acute group compared to Vehicle-Acute (P < 0.001) and EFV25-Acute (P < 0.05) groups. On the other hand, in subchronic treatment, a significant decrease in this neurotransmitter levels was found in EFV50-Subchronic group com-pared to subchronic control (P < 0.001). When comcom-pared different protocols, we observed a significant decrease in NE levels in EFV50-Subchronic group compared to EFV50-Acute group (P < 0.001) (Fig. 5D).
3.5. Effect of acute and subchronic efavirenz treatment in the striatal levels of glutamate and GABA
3.5.1. Glutamate
As shown inFig. 6A, two-way ANOVA revealed a significant main effect of “efavirenz treatment” [F (2, 42) =5.624, P=0.0069] and “duration of treatment” [F (1, 42) =11.62, P=0.0015] in striatal glutamate levels, with a significant interaction between these factors [F (2, 42) =3.288, P=0.0471]. Post hoc test showed a significant increase in glutamate levels in EFV25-Subchronic (P < 0.05) and EFV50-Subchronic (P < 0.001) groups compared to subchronic vehicle controls. When compared different protocols, we noted a significant increase in striatal glutamate in Subchronic group vs EFV50-Acute group (P < 0.01). Both subchronic EFV-treated groups presented increased glutamate levels compared to acute vehicle controls (P < 0.05). No significant difference was observed between acute EFV-treated groups and their respective controls.
3.5.2. GABA
As shown inFig. 6B, two-way ANOVA revealed a significant main effect of “efavirenz treatment” [F (2, 42) =80.62, P < 0.0001] and “duration of treatment”[F (1, 42) =4.901, P=0.0323] in striatal GABA levels, without a significant interaction between these factors [F (2, 42) =1.057, P=0.3565]. Post hoc test showed a significant decrease in GABA levels in EFV25-Acute (P < 0.0001) and EFV50-Acute (P < 0.0001) groups compared to acute vehicle controls. Also, a significant
decrease in striatal GABA levels was found in EFV25-Subchronic (P < 0.01) and EFV50-Subchronic (P < 0.0001) groups compared to sub-chronic controls. When compared different duration protocols, we identified a significant decrease in GABA levels in EFV25-Subchronic and EFV50-Subchronic groups compared to acute controls (P < 0.0001).
4. Discussion
The present study showed that EFV treatment induced significant behavioral changes in healthy rats with important influence of dose and treatment duration. EFV also caused marked alterations in the striatal concentrations of monoamines, their metabolites and the neurotrans-mitters glutamate and GABA. Taken together, these results have potential relevance in clarifying the underlying mechanisms involved in neuropsychiatric effects of EFV, and could contribute to the development of new strategies to attenuate these effects and improve antiretroviral therapy.
Assessment of anxiety in rodents is commonly based on exposure to aversive environments and observation of motor and emotional response (Savićet al., 2004; Walf and Frye, 2007). Previous studies
demonstrated that EFV induces important anxiogenic-like alterations in rodents in tests such as novel openfield (OF) test (Gatch et al., 2013) and elevated plus maze (EPM) test (Romão et al., 2011). In our experiments, acute EFV treatment at both doses tested promoted an anxiogenic-like effect in EPM test, reducing the time spent in open arms, without affecting the number of entries. In the OF test, these animals showed a decreased exploratory behavior (number of rearings) and no changes in spontaneous locomotion (number of crossings). In the subchronic protocol, only the animals treated with higher EFV dose (50 mg/kg) presented an anxiogenic-like behavior, indicating by the reduction in the entries and time spent in the open arms. Also, the subchronic EFV treatment impaired the exploratory behavior in OF test, without changing locomotor activity. Therefore, corroborating Fig. 6.Effects of acute and subchronic EFV treatment (EFV25: 25 mg/kg; and EFV50: 50 mg/kg) on the levels of glutamate (A) and GABA (B) in striatum. Bars represent ±
mean S.E.M. of 8 animals/group.*P
≤0.05 vs CTL- Subchronic;**P
≤0.01 vs
CTL-Subchronic;***P
≤0.001 vs CTL- Acute or CTL-Subchronic;& & &P
≤0.001 vs
previous evidence, our results point out to the induction of a genuine anxiogenic-like effect of EFV in both duration protocols especially with higher tested dose.
In rodents, depression-like behavior can be successfully evaluated by the forced swimming test (FST) (Custódio et al., 2013; Nollet et al., 2013). Few studies have investigated the effects of EFV in predictive tests for depression such as the FST. Romão et al. (2011) tested animals chronically treated with EFV using the FST, but did notfind any alteration in relation to controls (Romão et al., 2011). However, a study conducted by O'Mahony and coworkers found that rats treated with EFV (for 34 days) exhibited an increased susceptibility to develop depressive-like behavior after an acute stressor (O'Mahony et al., 2005).
In this study, no change was observed in FST in the EFV acutely treated animals compared to controls. However, a marked increase in immobility time was observed in the rats subchronically treated with EFV. Further, the group treated with higher EFV dose (50 mg/kg) showed an increase in this parameter compared to the group treated with the lowest one (25 mg/kg), indicating a possible dose-related response in the depressive-like effect induced by EFV. Therefore, our results demonstrate that subchronic EFV treatment induced a marked depressive-like behavior in rodents which was dose-related. Taken together, our results are in accordance with previous reports of more frequent and severe adverse neuropsychiatric events in patients using high EFV dosage and under longer duration treatment (Dhoro et al., 2015; Gutiérrez et al., 2005; Hawkins et al., 2005).
Major changes in monoaminergic neurotransmission has been implicated in the emergence of anxiety-like behavior in rodents (Morilak et al., 2005; Pusceddu et al., 2015). An increase in hippo-campal and striatal 5-HT release and 5-HT1Areceptor activation was observed in rodents subjected to social defeat stress when tested in OF and EPM tests (Rex et al., 2005; Watt et al., 2009). Also, a diverse array of acute stressors can induce an increase in NE and DA levels in limbic areas, which significantly correlated with anxiogenic-like alterations in behavioral tests (Leranth and Hajszan, 2007; Morilak et al., 2005). In our experiments, the acute EFV, mainly at the higher dose, promoted a marked increase in striatal levels of 5-HT, DA and NE, besides inducing important anxiogenic-like alterations in rats. In addition, these sub-jects presented a reduction in striatal DA turnover rate, which suggest a possible reduction in DA metabolism and a potential increase in its bioavailability in synaptic processes. Therefore, we suggest that the acute EFV treatment can promote an acute activation of these neurotransmission systems in striatum, which could be importantly involved in the induction of its anxiogenic-like effect.
The role of monoamines has also been classically highlighted in the pathophysiology of depression. The depression monoaminergic theory postulates that insufficient levels of monoamines would be responsible for the development of depressive symptoms (Elhwuegi, 2004; Hamon and Blier, 2013). In this study, besides inducing depression-like effect in healthy animals, subchronic EFV treatment markedly reduced the striatal levels of HT, DA and NE and increased the turnover rate of 5-HT and DA. The increase in these monoamine turnover rates suggests an increase in their metabolism and a potential reduction in their bioavailability for the neuronal processes. Furthermore, the effect of subchronic EFV on DA levels and its turnover ratio was considerably more prominent at the higher dose tested. This may explain (at least partially) the most potent depressive-like effect evidenced with this dose (Drago et al., 1986; Homberg et al., 2002).
There are few published studies evaluating the effects of EFV in the monoamine transmission.Gatch et al. (2013)demonstrated that EFV binds to serotonin receptors 5-HT2Cand 5-HT2Aand acutely blocks the presynaptic serotonin (SERT) and dopamine (DAT) transporters (Gatch et al., 2013). Another recent study showed that acute EFV inhibits MAO-A enzyme. These researchers also reported that pro-longed exposure to EFV reduced the 5-HT2Areceptor density and the responsiveness to 5-HT (Dalwadi et al., 2016). This evidence partially
corroborates ourfindings, since they pointed out to an acute increase in the brain levels of monoamines and reduction in metabolism rates after acute EFV treatment. Further, these data could provide possible explanations for the apparent opposite results observed here in the effects of acute and subchronic EFV treatment in the monoamine levels, since this change in the neurotransmitter levels could be a part of a time-dependent neuroadaptative process to EFV exposure with potential relevance to explain the emergence of different types of neuropsychiatric adverse effects over the course of treatment.
Recently, great emphasis has been given to the role of excitatory (glutamate) and inhibitory (GABA) neurotransmission in control of brain function, especially, emotional behavior. In this study, subchro-nic (and not acute) EFV treatment markedly increased glutamate levels in striatum. Previous evidence demonstrated that EFV has severe toxic effects on glial cells, mainly astrocytes. Since astrocytes are the most important cells involved in glutamate reuptake and metabolism in the CNS (Apostolova et al., 2015; Brandmann et al., 2013), it is possible to hypothesize that EFV may compromise astrocyte recycling and reup-take of glutamate, resulting in abnormal levels as observed in this study. Furthermore, given the role of the glutamatergic neurotransmis-sion to synaptic connectivity in important brain areas to mood control (Pilc et al., 2013), we propose that these abnormalities are potentially involved in the development chronic/subchronic neuropsychiatric effects of EFV, of note, its depression-like effect.
GABAergic dysfunction is also importantly involved in the devel-opment of affective disorders (Möhler, 2012). An overall deficit of GABA receptor function has been described in patients with anxiety disorders (Cameron et al., 2007; Hasler et al., 2007). Similarly, a reduction in GABAergic neurons (Rajkowska et al., 2007) and cere-brospinalfluid GABA levels was demonstrated in depressive patients (Sanacora et al., 2000). Further, drugs that positively modulate GABA A receptors usually have anxiolytic effect, while drugs that inhibit this transmission precipitate anxious states (Argyropoulos et al., 2000; Nemeroff, 2003).
In our experiments, acute and subchronic EFV, mainly at the higher tested dose, reduced GABA levels in striatum. Interestingly, the animals acutely treated with EFV showed anxiogenic-like alterations, while the subchronically treated ones showed both anxiogenic-like and depressive-like behavior. Therefore, since the GABAergic system is implicated in the development of both these symptoms, we suggest that EFV could affect this neurotransmission system since a single admin-istration and more prominently after a repeated exposure, contributing for the emergence and progression of its neuropsychiatric effects.
Some limitations need to be considered in this study. Firstly, we assessed the neurotransmitter concentrations in total tissue samples and were unable to draw specific conclusions about the extracellular levels of these compounds. Additionally, despite the mentioned re-levance of striatum to neurotransmitter circuits involved in mood-anxiety disorders, we did not evaluate other equally important brain areas, such as frontal cortex, hippocampus and amygdala, which remains as a target for future studies.
5. Conclusions
Conflict of interests
The authors declare no conflict of interests.
Acknowledgements
The authors thank the Brazilian Institutions Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico (FUNCAP) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for thefinancial support of this study. The authors thank to David John Woods (University of Otago, New Zealand) for his valuable comments and review of this study.
References
Ammassari, A., Murri, R., Pezzotti, P., Trotta, M.P., Ravasio, L., De Longis, P., Lo Caputo, S., Narciso, P., Pauluzzi, S., Carosi, G., Nappa, S., Piano, P., Izzo, C.M., Lichtner, M., Rezza, G., Monforte, A., Ippolito, G., d'Arminio Moroni, M., Wu, A.W., Antinori, A., 2001. Self-reported symptoms and medication side effects influence adherence to highly active antiretroviral therapy in persons with HIV infection. J. Acquir. Immune Defic. Syndr. 28, 445–449.
Apostolova, N., Funes, H.A., Blas-Garcia, A., Galindo, M.J., Alvarez, A., Esplugues, J.V., 2015. Efavirenz and the CNS: what we already know and questions that need to be
answered. J. Antimicrob. Chemother..http://dx.doi.org/10.1093/jac/dkv183.
Archer, J., 1973. Tests for emotionality in rats and mice: a review. Anim. Behav. 21, 205–235.
Argyropoulos, S.V., Sandford, J.J., Nutt, D.J., 2000. The psychobiology of anxiolytic drug. Part 2: Pharmacological treatments of anxiety. Pharmacol. Ther. 88, 213–227. Arribas, J.R., 2003. Efavirenz: enhancing the gold standard. Int. J. STD AIDS 14 (Suppl
1), 6–14.http://dx.doi.org/10.1258/095646203322491824.
Berrocoso, E., Ikeda, K., Sora, I., Uhl, G.R., Sánchez-Blázquez, P., Mico, J.A., 2012. Active behaviours produced by antidepressants and opioids in the mouse tail
suspension test. Int. J. Neuropsychopharmacol., 1–12.http://dx.doi.org/10.1017/
S1461145711001842.
Bezabhe, W.M., Chalmers, L., Bereznicki, L.R., Peterson, G.M., 2016. Adherence to antiretroviral therapy and virologic failure: a meta-analysis. Med. (Baltim.) 95,
e3361.http://dx.doi.org/10.1097/MD.0000000000003361.
Brandmann, M., Nehls, U., Dringen, R., 2013. 8-Hydroxy-efavirenz, the primary
metabolite of the antiretroviral drug Efavirenz, stimulates the glycolyticflux in
cultured rat astrocytes. Neurochem. Res. 38, 2524–2534.http://dx.doi.org/
10.1007/s11064-013-1165-2.
Cameron, O.G., Huang, G.C., Nichols, T., Koeppe, R.A., Minoshima, S., Rose, D., Frey, K.A., 2007. Reduced gamma-aminobutyric acid(A)-benzodiazepine binding sites in insular cortex of individuals with panic disorder. Arch. Gen. Psychiatry 64, 793–800.
http://dx.doi.org/10.1001/archpsyc.64.7.793.
Chiu, K., Lau, W.M., Lau, H.T., So, K.-F., Chang, R.C.-C., 2007. Micro-dissection of rat
brain for RNA or protein extraction from specific brain region. J. Vis. Exp. 269.
http://dx.doi.org/10.3791/269.
Custódio, C.S., Mello, B.S.F., Cordeiro, R.C., de Araújo, F.Y.R., Chaves, J.H., Vasconcelos, S.M.M., Nobre Júnior, H.V., de Sousa, F.C.F., Vale, M.L., Carvalho, A.F., Macêdo,
D.S., 2013. Time course of the effects of lipopolysaccharide on prepulse inhibition
and brain nitrite content in mice. Eur. J. Pharm. 713, 31–38.http://dx.doi.org/
10.1016/j.ejphar.2013.04.040.
Dalwadi, D.A., Kim, S., Amdani, S.M., Chen, Z., Huang, R.-Q., Schetz, J.A., 2016. Molecular mechanisms of serotonergic action of the HIV-1 antiretroviral efavirenz.
Pharmacol. Res. 110, 10–24.http://dx.doi.org/10.1016/j.phrs.2016.04.028.
Dhoro, M., Zvada, S., Ngara, B., Nhachi, C., Kadzirange, G., Chonzi, P., Masimirembwa, C., 2015. CYP2B6*6, CYP2B6*18, body weight and sex are predictors of efavirenz pharmacokinetics and treatment response: population pharmacokinetic modeling in
an HIV/AIDS and TB cohort in Zimbabwe. BMC Pharmacol. Toxicol. 16, 4.http://
dx.doi.org/10.1186/s40360-015-0004-2.
Drago, F., Caldwell, J.D., Pedersen, C.A., Continella, G., Scapagnini, U., Prange, A.J., 1986. Dopamine neurotransmission in the nucleus accumbens may be involved in oxytocin-enhanced grooming behavior of the rat. Pharmacol. Biochem. Behav. 24, 1185–1188.
Elhwuegi, A.S., 2004. Central monoamines and their role in major depression. Prog.
Neuro-Psychopharmacol. Biol. Psychiatry 28, 435–451.http://dx.doi.org/10.1016/
j.pnpbp.2003.11.018.
Gatanaga, H., Hayashida, T., Tsuchiya, K., Yoshino, M., Kuwahara, T., Tsukada, H., Fujimoto, K., Sato, I., Ueda, M., Horiba, M., Hamaguchi, M., Yamamoto, M., Takata, N., Kimura, A., Koike, T., Gejyo, F., Matsushita, S., Shirasaka, T., Kimura, S., Oka, S., 2007. Successful efavirenz dose reduction in HIV type 1-infected individuals with
cytochrome P450 2B6*6 and *26. Clin. Infect. Dis. 45, 1230–1237.http://
dx.doi.org/10.1086/522175.
Gatch, M.B., Kozlenkov, A., Huang, R.-Q., Yang, W., Nguyen, J.D., González-Maeso, J., Rice, K.C., France, C.P., Dillon, G.H., Forster, M.J., Schetz, J.A., 2013. The HIV antiretroviral drug efavirenz has LSD-like properties. Neuropsychopharmacology 38,
2373–2384.http://dx.doi.org/10.1038/npp.2013.135.
Gutiérrez, F., Navarro, A., Padilla, S., Antón, R., Masiá, M., Borrás, J., Martín-Hidalgo, A., 2005. Prediction of neuropsychiatric adverse events associated with long-term
efavirenz therapy, using plasma drug level monitoring. Clin. Infect. Dis. 41,
1648–1653.http://dx.doi.org/10.1086/497835.
Hammer, S.M., Eron, J.J., Reiss, P., Schooley, R.T., Thompson, M.A., Walmsley, S., Cahn, P., Fischl, M.A., Gatell, J.M., Hirsch, M.S., Jacobsen, D.M., Montaner, J.S.G., Richman, D.D., Yeni, P.G., Volberding, P.A., 2008. Antiretroviral treatment of adult HIV infection: 2008 recommendations of the International AIDS Society-USA panel.
JAMA 300, 555–570.http://dx.doi.org/10.1001/jama.300.5.555.
Hamon, M., Blier, P., 2013. Monoamine neurocircuitry in depression and strategies for
new treatments. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 45, 54–63.http://
dx.doi.org/10.1016/j.pnpbp.2013.04.009.
Hasler, G., van der Veen, J.W., Tumonis, T., Meyers, N., Shen, J., Drevets, W.C., 2007. Reduced prefrontal glutamate/glutamine and gamma-aminobutyric acid levels in major depression determined using proton magnetic resonance spectroscopy. Arch.
Gen. Psychiatry 64, 193–200.http://dx.doi.org/10.1001/archpsyc.64.2.193.
Hawkins, T., Geist, C., Young, B., Giblin, A., Mercier, R.C., Thornton, K., Haubrich, R.,
2005. Comparison of neuropsychiatric side effects in an observational cohort of
efavirenz- and protease inhibitor-treated patients. HIV Clin. Trials 6, 187–196.
http://dx.doi.org/10.1310/hct.2005.6.4.002.
Homberg, J.R., van den Akker, M., Raasø, H.S., Wardeh, G., Binnekade, R., Schoffelmeer, A.N.M., de Vries, T.J., 2002. Enhanced motivation to self-administer cocaine is predicted by self-grooming behaviour and relates to dopamine release in the rat medial prefrontal cortex and amygdala. Eur. J. Neurosci. 15, 1542–1550.
Konradi, C., Kornhuber, J., Sofic, E., Heckers, S., Riederer, P., Beckmann, H., 1992.
Variations of monoamines and their metabolites in the human brain putamen. Brain
Res. 579, 285–290.http://dx.doi.org/10.1016/0006-8993(92)90062-E.
Leranth, C., Hajszan, T., 2007. Extrinsic afferent systems to the dentate gyrus. Prog.
Brain Res. 163, 63–84.http://dx.doi.org/10.1016/S0079-6123(07)63004-0.
Lister, R.G., 1987. The use of a plus-maze to measure anxiety in the mouse. Psychopharmacol. (Berl.) 92, 180–185.
Mastan, S., Kumar, K.E., 2009. Influence of non-nucleoside reverse transcriptase
inhibitors (efavirenz and nevirapine) on the pharmacodynamic activity of gliclazide
in animal models. Diabetol. Metab. Syndr. 1, 15.
http://dx.doi.org/10.1186/1758-5996-1-15.
Meier, T.B., Drevets, W.C., Wurfel, B.E., Ford, B.N., Morris, H.M., Victor, T.A., Bodurka, J., Kent Teague, T., Dantzer, R., Savitz, J., 2015. Relationship between neurotoxic kynurenine metabolites and reductions in right medial prefrontal cortical thickness
in major depressive disorder. Brain Behav. Immun..http://dx.doi.org/10.1016/
j.bbi.2015.11.003.
Möhler, H., 2012. The GABA system in anxiety and depression and its therapeutic
potential. Neuropharmacology 62, 42–53.http://dx.doi.org/10.1016/
j.neuropharm.2011.08.040.
Morilak, D.A., Barrera, G., Echevarria, D.J., Garcia, A.S., Hernandez, A., Ma, S., Petre, C.O., 2005. Role of brain norepinephrine in the behavioral response to stress. Prog.
Neuropsychopharmacol. Biol. Psychiatry 29, 1214–1224.http://dx.doi.org/
10.1016/j.pnpbp.2005.08.007.
Mothapo, K.M., Schellekens, A., van Crevel, R., Keuter, M., Grintjes-Huisman, K., Koopmans, P., van der Ven, A., 2015. Improvement of depression and anxiety after discontinuation of long- term Efavirenz treatment. CNS Neurol. Disord. Drug Targets 14, 811–818.
Nemeroff, C.B., 2003. The role of GABA in the pathophysiology and treatment of anxiety disorders. Psychopharmacol. Bull. 37, 133–146.
NIH, N.R.C. (US) C. for the U. of the G. for the C. and U. of L, 2011. Guide for the Care and Use of Laboratory Animals. National Academies Press, Washington, D.C..
http://dx.doi.org/10.17226/12910.
Nollet, M., Le Guisquet, A.-M., Belzung, C., 2013. Models of depression: unpredictable
chronic mild stress in mice. Curr. Protoc. Pharmacol..http://dx.doi.org/10.1002/
0471141755.ph0565s61.
O'Mahony, S.M., Myint, A.M., Steinbusch, H., Leonard, B.E., 2005. Efavirenz induces depressive-like behaviour, increased stress response and changes in the immune
response in rats. Neuroimmunomodulation 12, 293–298.http://dx.doi.org/
10.1159/000087107.
Paylor, R., Spencer, C.M., Yuva-Paylor, L. a., Pieke-Dahl, S., 2006. The use of behavioral
test batteries, II: Effect of test interval. Physiol. Behav. 87, 95–102.http://
dx.doi.org/10.1016/j.physbeh.2005.09.002.
Pilc, A., Wierońska, J.M., Skolnick, P., 2013. Glutamate-based antidepressants:
preclinical psychopharmacology. Biol. Psychiatry 73, 1125–1132.http://dx.doi.org/
10.1016/j.biopsych.2013.01.021.
Porsolt, R.D., Le Pichon, M., Jalfre, M., 1977. Depression: a new animal model sensitive to antidepressant treatments. Nature 266, 730–732.
Pusceddu, M.M., Nolan, Y.M., Green, H.F., Robertson, R.C., Stanton, C., Kelly, P., Cryan, J.F., Dinan, T.G., 2015. The omega-3 polyunsaturated fatty acid docosahexaenoic acid (DHA) reverses corticosterone-induced changes in cortical neurons. Int. J.
Neuropsychopharmacol..http://dx.doi.org/10.1093/ijnp/pyv130.
Rajkowska, G., O'Dwyer, G., Teleki, Z., Stockmeier, C.A., Miguel-Hidalgo, J.J., 2007. GABAergic neurons immunoreactive for calcium binding proteins are reduced in the prefrontal cortex in major depression. Neuropsychopharmacology 32, 471–482.
http://dx.doi.org/10.1038/sj.npp.1301234.
Reagan-Shaw, S., Nihal, M., Ahmad, N., 2008. Dose translation from animal to human
studies revisited. FASEB J. 22, 659–661.http://dx.doi.org/10.1096/fj.07-9574LSF.
Rex, A., Voigt, J.P., Fink, H., 2005. Anxiety but not arousal increases
5-hydroxytryptamine release in the rat ventral hippocampus in vivo. Eur. J. Neurosci.
22, 1185–1189.http://dx.doi.org/10.1111/j.1460-9568.2005.04251.x.
Romão, P.R.T., Lemos, J.C., Moreira, J., De Chaves, G., Moretti, M., Castro, A. a., Andrade, V.M., Boeck, C.R., Quevedo, J., Gavioli, E.C., 2011. Anti-HIV drugs
nevirapine and efavirenz affect anxiety-related behavior and cognitive performance
Sanacora, G., Mason, G.F., Krystal, J.H., 2000. Impairment of GABAergic transmission in depression: new insights from neuroimaging studies. Crit. Rev. Neurobiol. 14, 23–45.
Santos, P., Herrmann, A.P., Benvenutti, R., Noetzold, G., Giongo, F., Gama, C.S., Piato, A.L., Elisabetsky, E., 2017. Anxiolytic properties of N-acetylcysteine in mice. Behav.
Brain Res. 317, 461–469.http://dx.doi.org/10.1016/j.bbr.2016.10.010.
Savić, M.M., Obradović, D.I., Ugresić, N.D., Cook, J.M., Yin, W., Bokonjić, D.R., 2004.
Bidirectional effects of benzodiazepine binding site ligands in the elevated
plus-maze: differential antagonism byflumazenil and beta-CCt. Pharmacol. Biochem.
Behav. 79, 279–290.http://dx.doi.org/10.1016/j.pbb.2004.07.013.
Thompson, M.A., Aberg, J.A., Hoy, J.F., Telenti, A., Benson, C., Cahn, P., Eron, J.J., Günthard, H.F., Hammer, S.M., Reiss, P., Richman, D.D., Rizzardini, G., Thomas,
D.L., Jacobsen, D.M., Volberding, P.A., 2012. Antiretroviral treatment of adult HIV infection: 2012 recommendations of the International Antiviral Society-USA panel.
JAMA 308, 387–402.http://dx.doi.org/10.1001/jama.2012.7961.
von Giesen, H.J., Köller, H., Theisen, A., Arendt, G., 2002. Therapeutic effects of nonnucleoside reverse transcriptase inhibitors on the central nervous system in HIV-1-infected patients. J. Acquir. Immune Defic. Syndr. 29, 363–367.
Walf, A.A., Frye, C.A., 2007. The use of the elevated plus maze as an assay of
anxiety-related behavior in rodents. Nat. Protoc. 2, 322–328.http://dx.doi.org/10.1038/
nprot.2007.44.
Watt, M.J., Burke, A.R., Renner, K.J., Forster, G.L., 2009. Adolescent male rats exposed to social defeat exhibit altered anxiety behavior and limbic monoamines as adults.