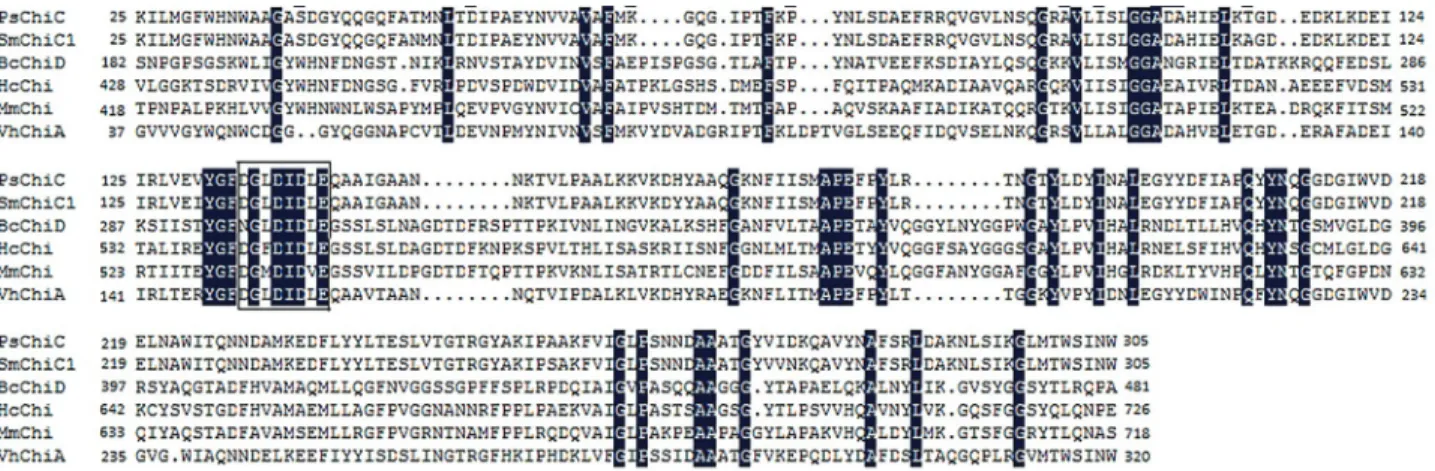
The chitinase C gene PsChiC from Pseudomonas sp. and its synergistic
Texto
Imagem


Documentos relacionados
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
Infelizmente, a entrevista com o Professor Remo Entelman não pode ser realizada em face de seu falecimento poucos meses antes da entrevista; conseguindo realizar, destarte,
O presente trabalho tem como objetivo fazer um estudo de ações de grupos e semigrupos inversos sobre bicategorias de C*-álgebras com diferentes formas de morfismos, como
Também Santo Agostinho, nas Confissões, ao fazer «avant-la-lieítre», a luminosa fenomenologia do pecado, concluía que ele era fruto de um perverso amor a Deus, ÍNa verdade, todo o
CSJ-3 were investigated in batch fermentation, and the maximum yield of acetic acid reached up to 0.49 % during 30 h cultivation under the optimum growth condition,
589 However, the continuity of the narrow Finisterra Block is 590 not obvious because its boundary with the Iberian Terrane 591 is marked by a lithospheric shear zone (the PTFSZ) and
