Je suis très reconnaissante que vous m'ayez formé avec un grand intérêt aux métiers de chercheur et d'enseignant. Je remercie sincèrement Annie et Robert de toujours me soutenir et de m'aimer comme leur propre fille.
La voie de protéolyse ciblée Ubiquitine-Protéasome 26S 30
Recherche de la fonction de TaGW2 dans le grain de blé tendre
Production de lignées de surexpression de TaGW2-A 125
Recherche des partenaires protéiques de TaGW2 par Y2H cytoplasmique 137
Recherche des partenaires protéiques de TaGW2 par Y2H nucléaire 140
44 Figure 45 : GW2 est un régulateur négatif de la largeur et du poids des grains. 114 Figure 77 : Alignement des séquences primaires des protéines homologues TaGW2-A, -B et –D ; et TaGW2-A dans le blé tendre et GW2 dans le riz.
Liste des Tableaux
LISTE DES ABBREVIATIONS
LOD: logarithm of probability NES: Core output signal NLS: Core location signal NoLS: Core location signal NS: Nul(le)s-ségrégant(s).
SYNTHESE BIBLIOGRAPHIQUE
Le blé
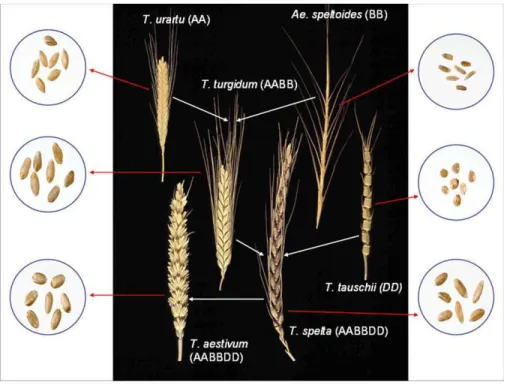
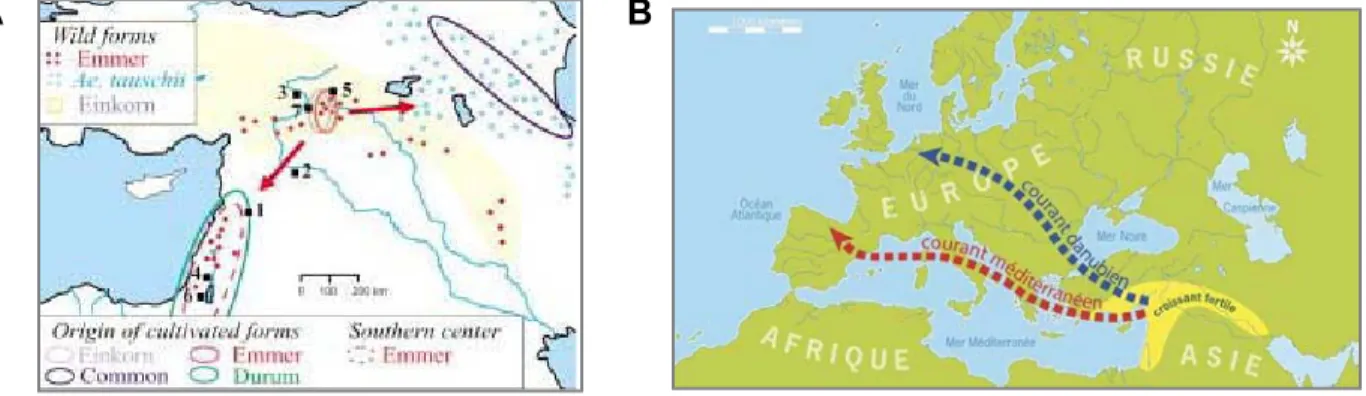
- Origines du blé tendre
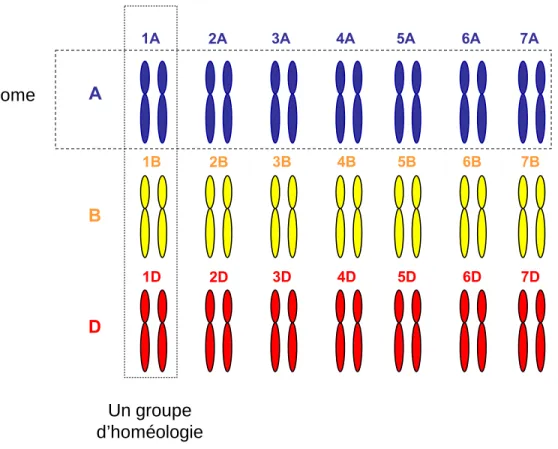
- Anatomie du blé
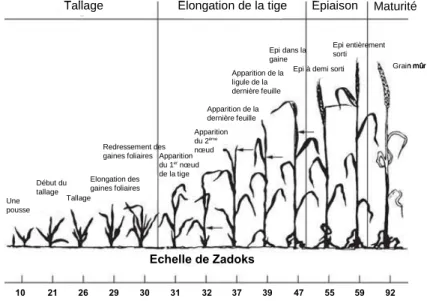
- Développement du blé
Le blé est originaire du sud-est de la Turquie, dans la région de Diyarbakir (Figure 4, A). Le système aérien du blé est constitué d'un certain nombre d'unités biologiques appelées capsules, toutes issues d'une zone située à la base de la plante, le plateau de culture.
2 Le grain de blé
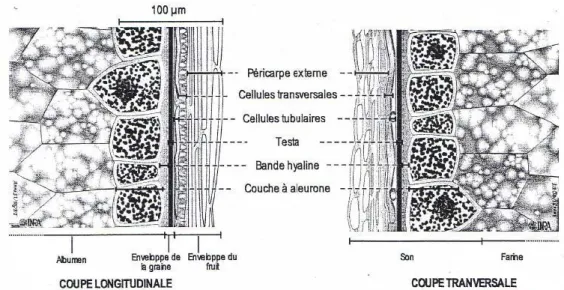
Structure du grain de blé
On retrouve de l'intérieur du grain vers l'extérieur : la couche nucléaire ou bande hyaline, la teste, les cellules tubulaires, les cellules transverses et le péricarpe externe. Les cellules tubulaires et transversales du péricarpe interne sont respectivement parallèles et perpendiculaires à l'axe longitudinal du grain.
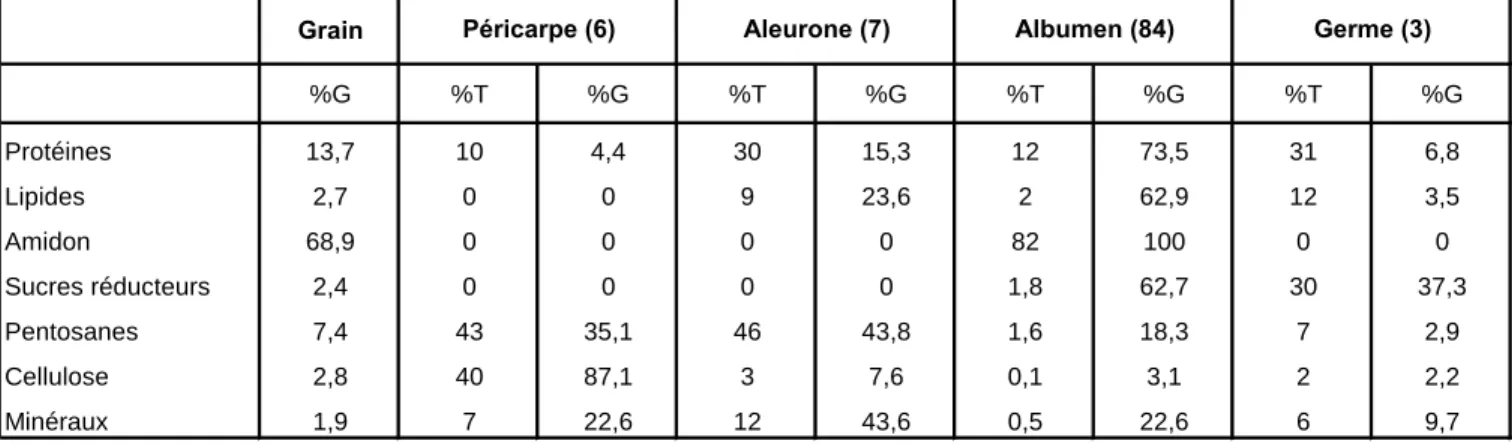
Composition du grain de blé
Granules de type B. Granules de type A. Granules de type C. Granules de type B. Granules de type A. Granules de type C. Les cellules périphériques de l'endosperme d'amidon contiennent uniquement des granules de type A, tandis que les cellules prismatiques et centrales de l'albumine d'amidon en contiennent trois types. de granulés (A, B et C).

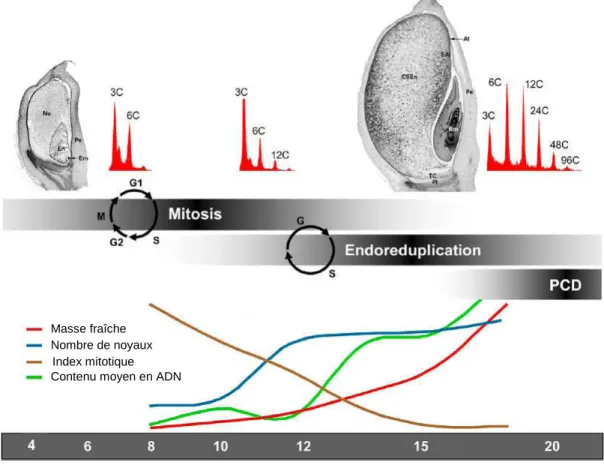
Développement du grain de blé
Une modulation de l'activité des CDK (cyclin-dependent kinases) semble particulièrement importante pour cette transition (Sabelli, 2012). Le rapport acide abscissique/auxine (ABA/Aux) affecte le remplissage des réserves de l'endosperme (Yang et al, 2001a ; Xu et al, 2007).
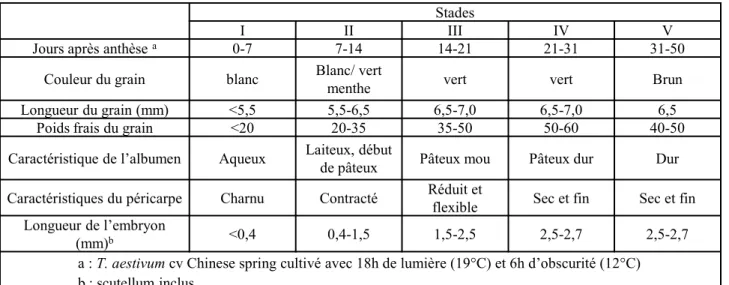
Le statut hydrique du grain au cours de son développement
La mort cellulaire programmée faciliterait l'hydrolyse des nutriments pour leur future absorption par l'embryon lors de la germination (Nguyen et al, 2007). Schnyder et Baum (1992) ont suggéré que la teneur en eau détermine le volume de croissance des grains pendant la phase de division cellulaire de l'endosperme. Lors de la phase de remplissage, l'état de l'eau et la dynamique d'accumulation de matière sèche sont étroitement liés (Schnyder et Baum, 1992 ; Calderini et Reynolds, 2000).
La fin de l'accumulation d'eau dans la protéine coïncide principalement avec la fin de l'accumulation de matière sèche (Schnyder et Baum, 1992).
L’effet des stress abiotiques sur le développement du grain
De plus, l'arrêt de la division cellulaire intervient à peu près en même temps que l'arrêt de cette accumulation rapide d'eau dans le grain (Nadaud et al, 2010). De plus, la taille finale des cellules serait partiellement affectée par l'accumulation d'eau (Egli, 1998). La réduction des activités métaboliques pendant la phase de maturation semble être particulièrement causée par cette perte massive d'eau (Young et Gallie, 2000).
Plusieurs études montrent que le stress hydrique réduit la taille des grains, en raccourcissant la période de remplissage des grains, mais pas le taux d'accumulation des réserves (Brooks et al, 1982 ; Nicolas et al, 1984 ; Kobata et al, 1992 ; Altenbach et al, 2003).
3 Le rendement en grains
Exemples de QTL de rendement en grains identifiés chez le blé
Dans le blé, de nombreux QTL contrôlant le rendement en grains et ses composants ont été identifiés génétiquement (McCartney et al, 2005 ; Kuchel et al, 2007 ; Gegas et al, 2010 ; Tableau 3). Bien que le nombre de QTL identifiés pour les composants de rendement soit très important, très peu de gènes contrôlant le rendement ont été clonés dans le blé tendre et aucun gène dans un GMP ou une taille de grain de blé contrôlé par QTL n'a été trouvé. ont été clonés et caractérisés fonctionnellement à ce jour (Yan et al Fu et al, 2009). En revanche, chez le riz, plusieurs gènes contrôlant les composants du rendement ont été isolés et étudiés fonctionnellement (Xing et Zhang, 2010).
Représentation schématique de l'effet des gènes GS3, GIF1, GW5 et GW2 sur le contrôle de la taille des grains de riz (Xing et Zhang, 2010).
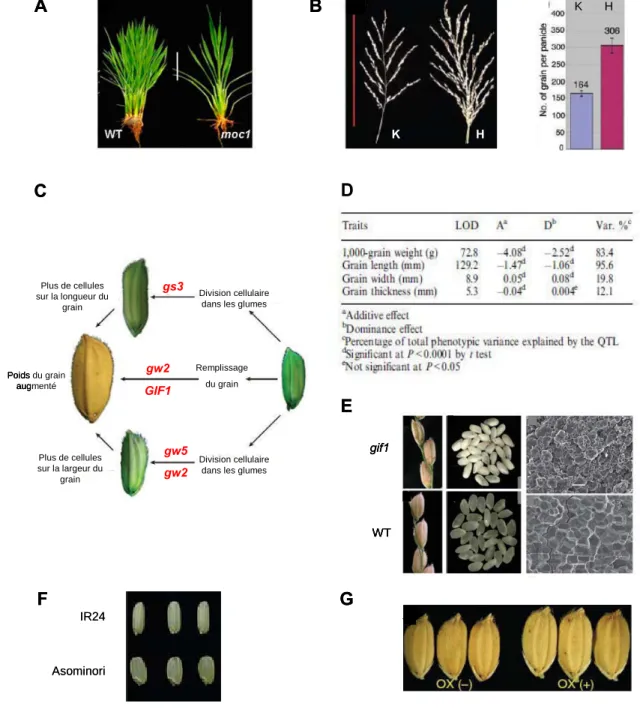
Exemples de gènes de composantes du rendement en grains isolés et caractérisés chez le riz
Les grains des variétés possédant l'allèle mutant ont des grains plus lourds en raison de l'augmentation du nombre de cellules dans les glumes qui permet indirectement une augmentation de la largeur des grains. Cependant, les auteurs ont montré que la protéine GW5 pouvait interagir physiquement avec les chaînes de polyubiquitine ; et suggèrent que GW5, comme GW2, est impliqué dans la régulation de la division cellulaire au cours du développement des grains de riz via la voie de protéolyse ciblée par l'ubiquitine-protéasome 26S. Récemment, le gène GS5 (Grain Size 5) a été identifié comme un QTL majeur pour la largeur, le poids et le remplissage des grains sur le chromosome 5 du riz (Li et al, 2011).
Le gène GS5 code pour une sérine carboxypeptidase et est un régulateur positif de la taille des granules car une augmentation de l'expression de GS5 est positivement corrélée à la largeur des granules (Figure 27, G).
4 L’ubiquitination
- L’ubiquitine
- La réaction enzymatique d’ubiquitination
- Les divers types d’ubiquitination
- Les enzymes déubiquitinases (DUBs)
- Les protéines Ubiquitin-like (UBLs)
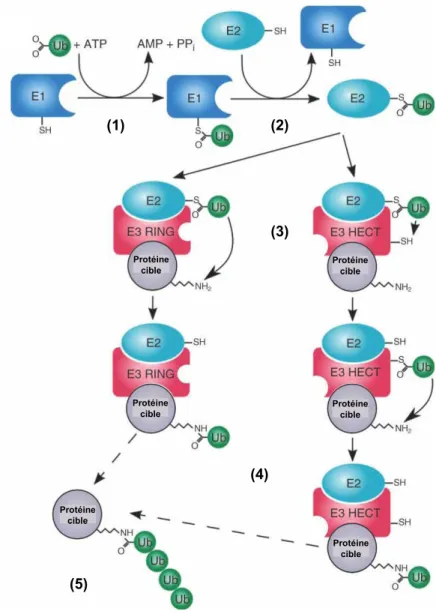
Après une réaction ATP-dépendante, l'enzyme E1 active l'ubiquitine (1), puis l'ubiquitine activée est transférée à l'enzyme E2 (2). Le produit final de cette réaction est la protéine substrat marquée avec un monomère d’ubiquitine. Plusieurs cycles répétés de cette réaction conduisent à l'attachement d'une chaîne d'ubiquitine (polyubiquitination) à la protéine cible (5).
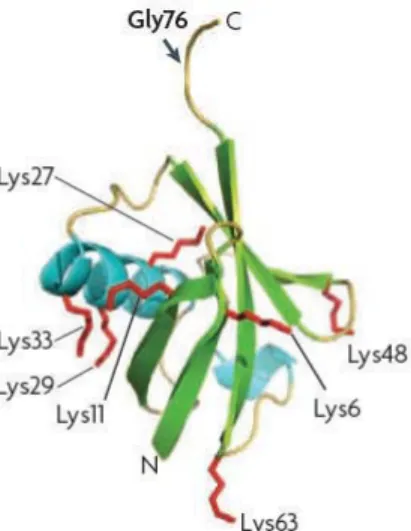
La réaction commence par la formation ATP-dépendante d'une liaison thioester entre la cystéine de l'enzyme E1 et la glycine 76 de l'ubiquitine.
5 La voie de protéolyse ciblée Ubiquitine-Protéasome 26S
Le protéasome 26S
20S, ou CP (core particule), est un complexe de 720 kDa composé de 28 sous-unités disposées en 4 anneaux heptamères empilés : 2 anneaux de sous-unités α aux extrémités de. Les sites protéolytiques actifs dans la chambre interne du CP sont les thréonines N-terminales des sous-unités β1, β2 et β5. Les sous-unités RPT utilisent l'hydrolyse de l'ATP pour faciliter le déploiement des polypeptides ciblés et donner accès à la chambre interne 20S (Hartmann-Petersen et al, 2003).
Les peptides clivés quittent le 20S par l'ouverture du cylindre grâce à l'énergie produite par l'hydrolyse de l'ATP, catalysée par les sous-unités ATPase de la base 19S.
Les fonctions de l’UPS
Il assure une dégradation indépendante de l'ubiquitine des protéines oxydées par des espèces d'oxygène activées et jouerait un rôle important dans les mécanismes de défense contre les potyvirus TMV et LMV en raison de son activité siARN (Kurepa et Smalle, 2008).
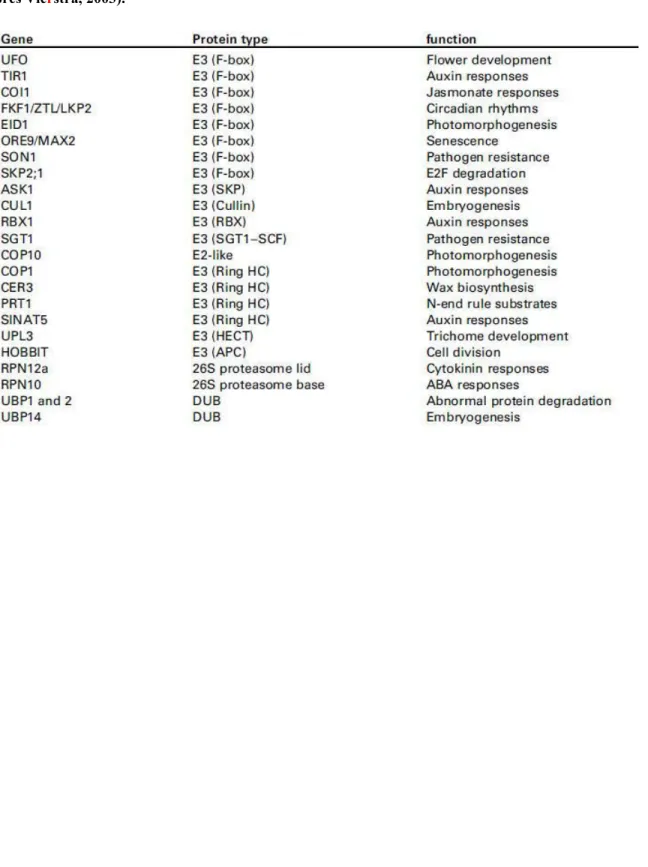
6 Les enzymes E3
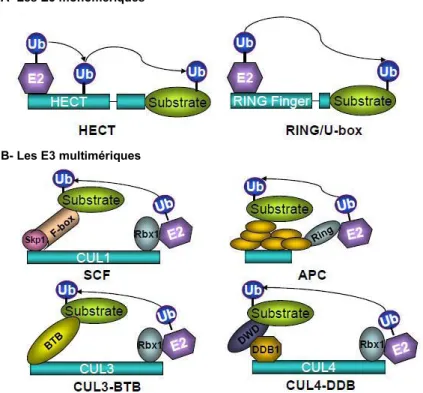
- Les E3 HECT
- Les E3 RING
- Les E3 U-box
- Les CRLs
Ils possèdent à la fois le domaine RING, requis pour l'interaction avec l'enzyme E2, et le(s) domaine(s) requis pour la reconnaissance des protéines substrats. Les phytochromes (phyA et phyB) et les cryptochromes (cry1 et cry2) sont les principaux photorécepteurs, qui perçoivent un large spectre de lumière visible. COP1 est un intégrateur et un répresseur central de la voie de signalisation de la croissance de la lumière visible chez les plantes (Figure 37).
La liaison de la protéine CAND1 à la sous-unité Cullin empêche alors la reformation du complexe (Petroski et Deshaies, 2005 ; Vierstra, 2009).
7 Les E3 et le rendement en grain chez les céréales
Les protéines E3 RING OsSDIR1, OsDIS1 et TdRF1
Le gène OsDIS1 a été identifié comme un régulateur négatif de la tolérance à la sécheresse du riz (Ning et al, 2011). Les plants de riz surexprimant et sous-exprimant OsDIS1 présentent respectivement une tolérance réduite et accrue à la sécheresse. Le gène TdRF1 a été identifié pour la première fois comme un gène dont l'expression répondait au stress dû au froid et à la sécheresse (Mastrangelo et al, 2005; De Leonardis et al, 2007).
Récemment, Guerra et ses collègues (2012) ont montré que la surexpression et la sous-expression de l'homologue de TdRF1 dans l'orge augmentaient et diminuaient la tolérance cellulaire à la sécheresse chez l'orge, respectivement.

Le complexe APC TAD1 ou APC TE
La suppression d'une adénine dans l'exon 4 (A316) génère un codon stop prématuré et donc une protéine GW2 tronquée. (C) Phénotypes des lignées isogéniques NIL (GW2) et variété FAZ1. Comparaison du grain (en haut, à gauche), de la hauteur de la plante (en haut, à droite) et de la talle (en bas, à gauche) entre la lignée isogénique NIL(GW2) et la variété FAZ1.
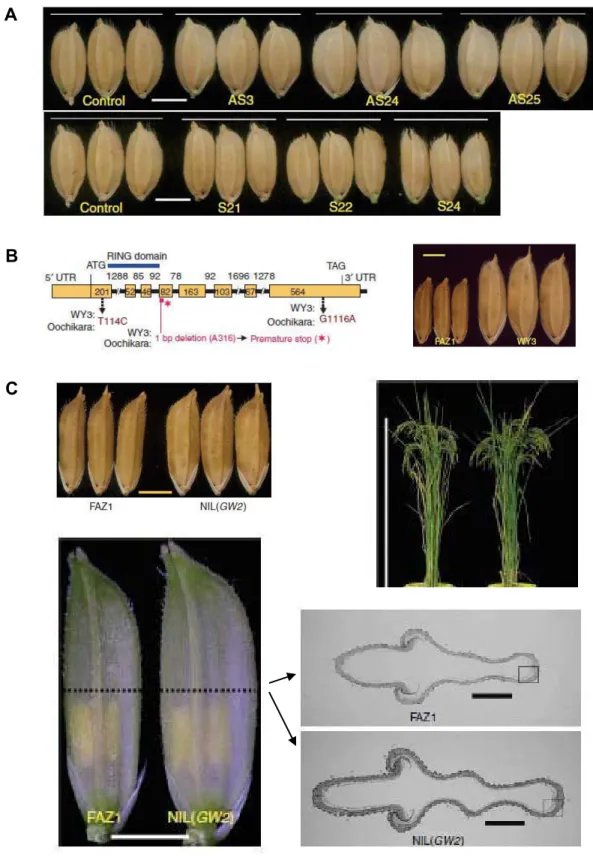
8 Le gène GW2
Les gènes homologues de GW2 chez le maïs, Zea mays
Une insertion-délétion de 8 bases et un SNP (transition C/T) dans le promoteur ainsi qu'un SNP (transition C/T) dans l'exon 8 de ZmGW2-CHR4 se sont également révélés associés à la largeur des grains dans 2 environnements. ZmGW2-CHR4 et ZmGW2-CHR5 sont exprimés de manière constitutive dans les épis, les grains de 0 à 20 jours après l'anthèse, l'inflorescence mâle, les enveloppes, les stigmates, les feuilles et les racines. Bien que très faible (R2=0,12), les auteurs suggèrent qu’il existe une corrélation négative entre le taux d’accumulation du transcrit ZmGW2-CHR4 et la largeur des grains, indiquant que le gène ZmGW2-CHR4 serait un régulateur négatif de la largeur et du rendement en grains tout aussi important. GW2 dans le riz (Figure 46, C).
Cependant, les auteurs mentionnent également le fait que malgré la corrélation entre la largeur des grains et le niveau d'expression de ZmGW2-CHR4, ainsi que l'association entre la largeur des grains et les polymorphismes identifiés dans ZmGW2-CHR4, aucun de ces polymorphismes n'affecte l'expression. niveau de ZmGW2-CHR4.
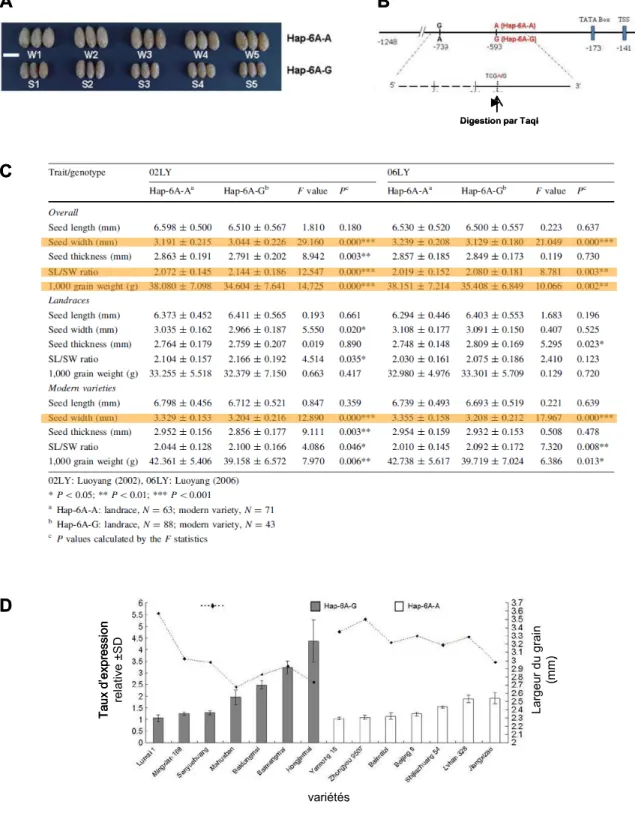
Les homologues de GW2 chez le blé tendre, Triticum aestivum
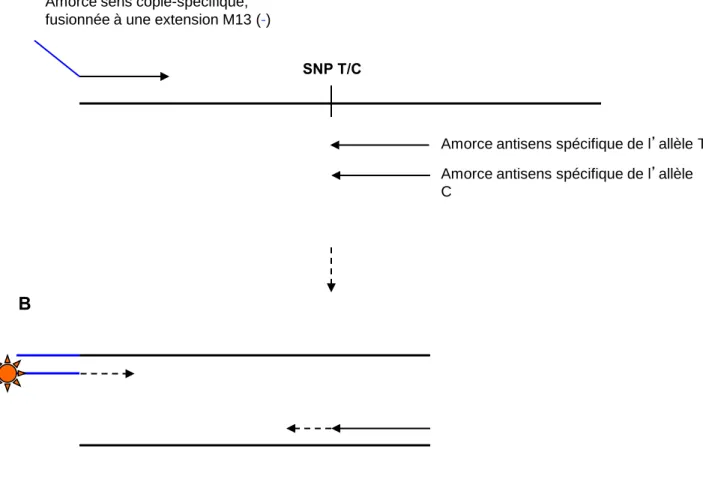
TSS, site d'initiation de la transcription. (C) Analyse d'association entre le marqueur SNP (-593) et les composants du rendement en grains, dans un panel de 265 accessions de blé tendre chinois, dans 2 environnements (Luoyang 2002 et 2006). (D) Relation entre le niveau d'expression relatif de TaGW2, toutes copies confondues, et la largeur des grains âgés de 10 DAA dans 7 cultivars portant l'allèle G et 7 cultivars portant l'allèle A du SNP (-593). L'analyse transcriptionnelle de TaGW2, toutes copies confondues, par qRT-PCR a été réalisée sur 10 grains anciens DAA dans 14 cultivars du panel d'association (7 cultivars avec l'allèle G pour le marqueur SNP à -593 et 7 cultivars avec l'allèle A ; Figure 47, D). A 10 DAA, les auteurs ont montré que le degré d'accumulation des transcrits de TaGW2, toutes copies confondues, était négativement corrélé à la largeur des grains (R2 = 0,36 et 0,43 chez les cultivars avec l'allèle G et l'allèle A, respectivement.
Les auteurs affirment que le niveau d'expression moyen de TaGW2 est plus élevé dans les variétés avec l'allèle G que dans les variétés avec l'allèle A, bien qu'un test t de Student sur les données fournies dans l'article ne montre aucune différence significative entre les taux d'accumulation de transcription. entre 2 groupes de variétés (p=0,12).
9 Objectifs du travail de thèse
Les haricots de la variété Wanmai 38 sont plus larges mais plus courts que les haricots de la variété Lankaodali. A l’issue de cette étude et contrairement aux travaux de Su et collègues (2011), les auteurs suggèrent que TaGW2-A affecterait négativement la taille et le poids final des grains du blé, par un mécanisme similaire à celui du GW2 dans le riz. .
MATERIEL ET METHODES
Il permet la fusion de fragments d'ADN à l'extrémité C-terminale et dans le cadre de lecture de l'assemblage NubG-HA. La taille de la séquence sélectionnée pour la cassette ARNi : Une taille minimale de la séquence insérée est requise pour une inactivation efficace de l’expression des gènes. La construction de la cassette RNAi-TaGW2 et l'obtention des lignées transgéniques T0 par transformation d'embryons immatures de blé tendre à l'aide de biolistes ont été réalisées par la plateforme de validation fonctionnelle UMR.
Les plantes transformées ont été génotypées pour la présence de la cassette par PCR sur ADN génomique, en utilisant le couple d'amorces PDK-intron permettant l'amplification de l'intron de 387 pb du gène PDK du transgène. Le volume des grains a été estimé par la méthode du déplacement d'eau à l'aide d'un pycnomètre. Tous les grains restant de l'épi ont été comptés, regroupés, pesés et moulus en farine complète à l'aide d'un moulin (moulin Cyclotec 6800).