Je remercie évidemment tous les membres de l'équipe GQMP, présents et passés, ainsi que la plateforme PAPPSO de m'avoir permis de me sentir intégrée et valorisée dès mon arrivée, ainsi que d'être toujours disponible lorsque j'avais besoin d'aide. aide ou conseil. Je tiens également à remercier tous les membres, actuels et anciens, de la ferme du Moulon, et tout particulièrement Dominique de Vienne de m'avoir accueilli dans son unité.

La sécheresse chez les plantes
La sécheresse : définitions et risques liés au changement climatique
- Définition de la sécheresse
- Changement climatique et risques de sécheresse
- Stratégies mises en place face à la sécheresse
- L’efficacité de l’utilisation de l’eau
Chaque rectangle correspond à une stratégie mise en œuvre par les plantes pour faire face à la limitation hydrique : (1) la stratégie d'évitement implique une maturation précoce avant que le déficit hydrique ne devienne trop important ; (2) la stratégie d'évitement implique la conservation de l'eau dans les tissus pendant des périodes relativement courtes de stress sévère afin d'éviter des dommages irréversibles ; (3) la tolérance consiste à maintenir les fonctions principales de la plante malgré un déficit hydrique. Les plantes tolérantes conservent leurs fonctions (croissance foliaire et reproductrice, nombre d'organes, transpiration et photosynthèse) malgré le manque d'eau.

Les mécanismes physiologiques impliqués dans la réponse à la sécheresse
- La fermeture stomatique
- La modification de la conductivité hydraulique
- L’ajustement osmotique
- La réduction de la croissance des parties aériennes
Les mécanismes impliqués dans la régulation de la fermeture et de l'ouverture des stomates sont représentés sur la cellule de garde située respectivement à gauche et à droite. La modification de la conductivité hydraulique des tissus qui se produit au début d'un déficit hydrique correspond à la modulation du débit d'eau.

Les mécanismes moléculaires impliqués dans la réponse à la sécheresse
- La perception du signal
- La transduction du signal…
- Les glycérolipides
- Le calcium
- Les espèces réactives de l’oxygène…
- Les protéines kinases
- Les protéines phosphatases
- Les facteurs de transcription
- Les facteurs de transcription dépendants de l’ABA…
- Les facteurs de transcription indépendants de l’ABA
- Les fonctions des gènes induits par la sécheresse
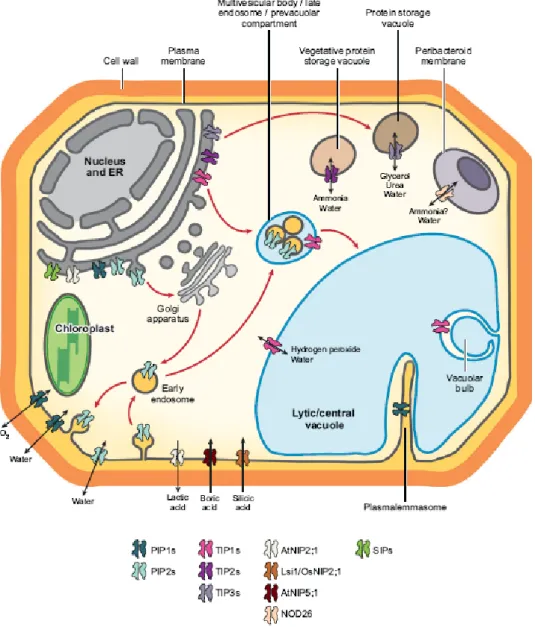
- Les aquaporines
- Les protéases
- La détoxification
- Les protéines HSP
- Les protéines LEA
Les membres de la sous-mille SnRK3 correspondent à des proteins SCaBP ("SOS3-like calcium sensor binding protein")/CIPK ("CBL(cobalamin)-interacting protein kinases") (Luan et al., 2009). We previously showed that water deficit caused a decrease in β-O-4 linked monolignols in DZ of maize leaf (Vincent et al., 2005).

La sensibilité du maïs à la sécheresse
- La sensibilité du maïs à la sécheresse au stade reproducteur
- La sensibilité du maïs à la sécheresse lors du développement végétatif…
L’amélioration de la réponse à la sécheresse
La transgénèse
- La transgénèse : définition et méthodologies
- La transgénèse comme outil de connaissance de la fonction des gènes…
- La transgénèse comme outil pour l’amélioration de la tolérance à la sécheresse
Il est important de le noter dans le contexte de l’amélioration de la tolérance à la sécheresse, car ce processus complexe repose sur des réactions impliquant de nombreux gènes. De nombreux travaux sont actuellement en cours dans les entreprises semencières pour améliorer la tolérance du maïs à la sécheresse grâce à la transgenèse.
La sélection génétique « classique »
- La production d’hydrides plus tolérants à la sécheresse
- Les ressources génétiques
Les programmes de sélection classiques, mis en œuvre par de nombreuses sociétés semencières et instituts de recherche nationaux et internationaux, étudient la variabilité génétique naturelle du maïs pour sélectionner les génotypes les plus favorables en termes de tolérance à la sécheresse. Cependant, les ressources génétiques naturelles sont encore sous-utilisées : seulement 5 % des ressources génétiques disponibles sont utilisées dans les hybrides de maïs commerciaux (Hoisington et al., 1999).
La sélection assistée par marqueurs
- La SAM : définition et exemples d’application
- Un modèle écophysiologique stable pour identifier des QTL de réponse à la
- Identification des régions chromosomiques conférant le phénotype
- Le clonage positionnel
- La stratégie gènes et protéines candidats
Le paramètre c correspond à la pente de la réponse du LER au potentiel hydrique du sol (D). L’utilisation de la génomique fonctionnelle a donc permis de nombreuses avancées dans l’analyse et l’identification des QTL.

Présentation du travail de thèse
Objectif
There is also evidence that maize silking and leaf growth under water deficit have common genetic determinisms (Welcker et al., 2007). Interestingly, four collocations were observed between ASI trait under water deficit (ASIdeficit) and LERdeficit (Welcker et al., 2007). The candidate gene ZmMYB31 belongs to subgroup 4 of the R2R3-MYB transcription factor family (Fornale et al., 2006).
Proteins were solubilized in UKS buffer ( Mechin et al., 2007a ) and quantified using the 2-D Quant Kit (GE Healthcare Life Sciences). Fatty acid methyl esters were obtained by transfection and analyzed essentially as described ( Domergue et al., 2010 ). In the case of rice we adopted the nomenclature published during the time course of this study (Philippe et al., 2010).
For plants, responses to water deficit involve both ABA-dependent and ABA-independent pathways ( Nakashima et al., 2009 ). Agrobacterium-mediated transformation of the inbred maize line A188 was based on a published protocol (Ishida et al., 1996). Proteins were solubilized in UKS buffer ( Mechin et al., 2007 ) and quantified using the 2-D Quant Kit (GE Healthcare Life Sciences).

Publication 1. Transcriptomic and proteomic profiling of bulked recombinant maize
Supplément expérimental : production et caractérisation de plantes de maïs
- Production des souches d’agrobactérie utilisées pour la transformation du maïs et
- Sélection des lignées transgéniques primaires
- Caractérisation des lignées transgéniques T1
- Analyse de cinétiques de dessèchement des plantes T1…
- Abondance relative du transgène dans les plantes T1
- Conclusion
Pour chaque gène d'intérêt, Biogemma a entrepris la transformation, la détermination du nombre de copies intactes insérées dans les transformants primaires (T0), la production de grains T1 et l'évaluation de la cinétique de séchage des plantes. transgéniques. J'ai moi-même évalué l'absence d'arrêt prématuré de la transcription du transgène dans les plantes T0, participé aux échantillons de feuilles nécessaires pour évaluer l'expression du transgène dans les plantes T1, puis analysé l'abondance relative du transgène dans les feuilles des plantes T1. Les transformants primaires (T0) ont été sélectionnés selon quatre critères : (1) nombre de copies insérées (PCR quantitative sur ADN génomique) ; (2) intégrité de l'ADN-T inséré (PCR sur ADN génomique) ; (3) absence de terminaison prématurée de la transcription du transgène dans les feuilles (RT-PCR utilisant une amorce sens située au niveau de l'intron de l'ubiquitine 3 du riz et une amorce antisens située au niveau du terminateur AtSac66) ; (4) nombre de grains récoltés T1.
Les plantes transgéniques de première génération (T1 ; croisement du transformant primaire avec la lignée parentale A188), hémizygotes pour le transgène, ont ensuite été caractérisées à deux niveaux : (1) évaluation de la tolérance au déficit hydrique par cinétique de dessiccation ; (2) détermination de l'abondance relative de chaque transcrit d'intérêt dans la feuille par qRT-PCR. Sur la base du taux de séchage et du RCW après 8 h, 12 événements semblaient intéressants pour des analyses plus approfondies : huit événements ZmMYB31 (un de la souche T01892 et sept de la souche T01893) et quatre événements ZmFatA de la souche T01908. Sur la base des données d'expression obtenues par RT-PCR sur des plantes T0 (résultats non présentés) et des données de cinétique de séchage obtenues sur des plantes T1, 29 événements ont été soumis à une nouvelle culture dans des conditions irriguées pour déterminer l'abondance relative du transgène présent dans leurs feuilles. en phase végétative.
Des plantes transgéniques dérégulées pour les gènes candidats ZmMYB31 et ZmFatA ont été obtenues en transformant la lignée de maïs A188 avec une cassette d'expression contenant chaque gène d'intérêt sous le contrôle du promoteur constitutif de l'ubiquitine 3 du riz.

Discussion générale
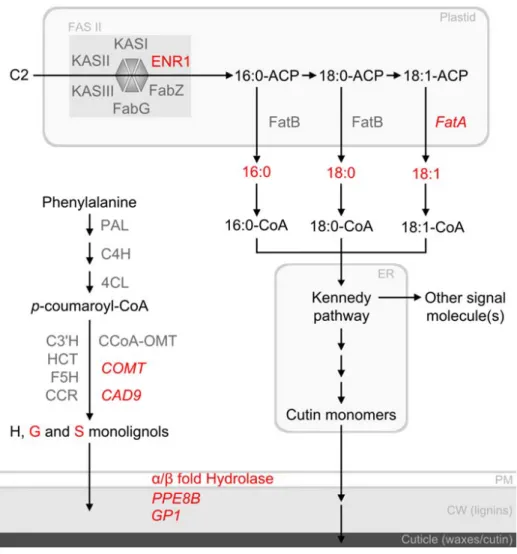
Modèle schématique des voies métaboliques potentiellement régulées par la protéine ZmMYB31 lors de la croissance des feuilles et de la soie dans des conditions de déficit hydrique. Des données récentes de la littérature montrent que les protéines AtMYB30 et AtMYB41 influencent respectivement l'expression des gènes AtFatB et AtFatA (Raffaele et al., 2008 ; Comineli et al., 2008). Il est possible que les acides gras formés soient métabolisés en d'autres composés non mesurés, tels que les monomères de cutine, un composant de la cuticule (Pollard et al., 2008).
Récemment, il a été démontré que l'augmentation de la cutine permet une réduction de la perméabilité épidermique et une diminution de la transpiration (Pollard et al., 2008 ; Kosma et al., 2009). Ainsi, en régulant l'expression du gène ZmFatA, la protéine ZmMYB31 pourrait influencer à la fois la quantité de protéine candidate ZmENR1 et la teneur en dérivés lipidiques capables de réduire la transpiration des feuilles dans des conditions de déficit hydrique (Fig. 6). Comme la protéine AtMYB41, la protéine ZmMYB31 pourrait également affecter divers composants de la membrane plasmique et de la paroi cellulaire (Fig. 6).
Il a été démontré que les changements dans la structure de la pectine affectent le développement des plantes et la réponse au stress (Pelloux et al., 2007).
Perspectives
88 JAZ libère alors le facteur de transcription MYC2, qui induit l'expression de gènes dépendants du jasmonate (Staswick, 2008). Il est possible que la forte expression du gène ZmJAZ2 dans la population favorable offre une protection contre le déficit hydrique. En effet, il a été démontré qu’un retard dans la sénescence des feuilles induit une tolérance au déficit hydrique chez Nicotania tabaccum cv.
Une alternative serait d'étudier l'expression du gène AtFatA chez le mutant mod1 codant pour un ENR dans A. Pour s'assurer qu'un des autres gènes candidats identifiés n'est pas impliqué dans la régulation du locus situé dans la région cible, on pourrait envisager : (1) la production d’autres plantes transgéniques qui surexpriment ou sous-expriment ces gènes ; (2) rechercher des mutants dans les collections ; (3) réaliser une analyse génétique d'association pour lier un polymorphisme dans un ou plusieurs de ces gènes à un changement phénotypique associé à la tolérance au déficit hydrique du maïs. Enfin, une façon d'étudier la fonction de gènes dont la fonction exacte reste inconnue ou qui ont une fonction inconnue pourrait être de compléter les mutants d'A. thaliana avec des ADNc de maïs dont le phénotype présente un changement caractéristique du paramètre cible.
C ARACTERISATION FONCTIONNELLE DE LA PROTEINE CANDIDATE
Implication de la protéine ZmASR1 dans la régulation de la biosynthèse des acides
- Publication 2 : The Zea mays abscisic acid-, stress- and ripening-induced protein
- Supplément méthodologique
These associations were confirmed by the observation that transgenic maize lines misexpressing ZmASR1 showed significant changes in leaf senescence under mild water deficit conditions in the field (Jeanneau et al., 2002). Overexpression of the tomato ASR1 gene (SlASR1) in tobacco plants also resulted in a reduced rate of water loss (Kalifa et al., 2004a). The amino acid positions identified as the A and B regions of the bipartite nuclear localization signal of the LLA23 protein (Wang et al., 2005) are underlined.
MZ00000739 (ZmSPL14) probably encoded a nuclear SQUAMOSA (SQUA) promoter-binding protein (SBP)-box transcription factor of the plant-specific subfamily IIa that is most closely related to AtSPL14 from Arabidopsis (Stone et al., 2005; Guo et al. , 2008. ). Consequently, the data support the idea that the Poaceae intermediate ancestor with 12 protochromosomes (Murat et al., 2010) already had 7 ASR genes. It is noteworthy that maize and rice (Philippe et al., 2010) ASR genes belonging to Poaceae subclades I-3 (ZmASR4 and OsASR6) and II-2 (ZmASR7-1 and OsASR1) were all up-regulated by water deficit in mature leaves.
À cet égard, une observation notable est que Canas et al. 2010) ont récemment montré qu'augmenter l'expression de l'enzyme P5CS pouvait avoir un effet bénéfique sur le rendement. La digestion par transfert de gel 2D et l'analyse LC-MS/MS ont été réalisées comme décrit ( Page et al., 2010 ). http://www.thegpm.org/TANDEM/). Sur les 25 gènes identifiés, 13 (quatre transcrits et neuf protéines) codent pour des cibles potentielles ou avérées des thiorédoxines (Trx) selon Montrichard et al.

Suppléments expérimentaux et méthodologiques
- Etude de l’impact de la sur-expression du gène ZmASR1 sur la régulation redox de
- Introduction
- Mise au point de conditions expérimentales permettant de visualiser les
- Influence de la protéine ZmASR1 sur la régulation redox des protéines
- Conclusion
- Mise en évidence de modifications post-traductionnelles par phosphorylation chez
- Introduction
- Développement d’outils biochimiques pour étudier les MPTs des protéines
- Identification de sites de phosphorylations in vitro chez la protéine
- Mise en évidence de phosphorylations in vivo chez les protéines ZmASR.162
- Conclusion
- Matériel et méthodes
- Matériel végétal et conditions de cultures
- Protéines recombinantes et anticorps
- Extraction et dosage des protéines en condition native…
- Analyses par électrophorèse monodimensionnelle
- Western blot
- Immunoprécipitation…
- Essai de phosphorylation in vitro
- Digestion enzymatique des protéines en solution
- Diméthylation des peptides sur colonnes
- Enrichissement en phosphoprotéines
- Enrichissement en phosphopeptides
- Analyse des peptides par spectrométrie de masse
- Visualisation de l’état redox des protéines
Discussion générale et perspectives
- Analyse fonctionnelle de la protéine ZmASR1 par phénotypage au champ et
- Effet de la protéine ZmASR1 sur le rendement au champ
- Identification de gènes dont l’expression est affectée par la sur-expression du
- Perspectives
- Etude des mécanismes de régulation des gènes ZmASR
- Analyse phylogénétique de la famille ZmASR
- Mise en évidence d’une régulation transcriptionnelle et post-transcriptionelle
- Mise en évidence d’une phosphorylation des protéines ZmASR
- Perspectives
Brevet d’invention : obtention de plantes ayant une tolérance améliorée à un
Brevet d’invention : obtention de plantes ayant une tolérance améliorée à un
Identification des sites de phosphorylation des protéines ZmASR2 et