Une analyse critique de la littérature montre que la variabilité environnementale peut générer des différences d’histoire de vie entre cohortes de naissance, ou effets de cohorte, par son influence sur la qualité des individus et sur l’intensité de la compétition intraspécifique au sein des espèces d’une cohorte et entre cohortes. Nos résultats, intégrés dans le contexte général de l’écologie des populations, soulignent l’importance de considérer la structure de la population et l’hétérogénéité du cycle biologique pour établir des prédictions fiables sur le sort des petites populations et mettre en œuvre des mesures de gestion efficaces.
INTRODUCTION GÉNÉRALE Des populations structurées
Structure de population et histoire de vie
La dépendance négative à la densité résulte d'une compétition intraspécifique pour l'accès à des ressources limitées telles que la nourriture ou l'espace (Begon et al., 2006). En revanche, il existe peu d’exemples empiriques d’effets démographiques Allee (voir l’analyse de Gregory et al., 2010).
Structure de population, système d’appariement et sélection sexuelle La sélection sexuelle résulte de la compétition entre individus pour l’accès aux partenaires
Structure de la population, système d'accouplement et sélection sexuelle La sélection sexuelle résulte de la compétition entre individus pour l'accès aux partenaires. Deux théories se contredisent concernant l'influence du sex-ratio opérationnel et de la densité sur le système d'accouplement et l'intensité de la sélection sexuelle.
Conséquences démographiques de la structure de population
Conséquences démographiques de la densité de population et de la structure par âge et par sexe La densité de population, la structure par âge et la structure par sexe peuvent fortement influencer sa dynamique (par exemple Edeline et al., 2010, Coulson et al., 2001). En outre, l'intensité de la sélection sexuelle pourrait menacer la persistance des populations (voir l'analyse de Kokko et Rankin, 2006 et les références citées).
CONTEXTE DE L’ÉTUDE
De plus, le système d'accouplement peut être important pour la persistance de la population, car les espèces polygames sont généralement moins sensibles aux effets Allee que les espèces monogames (par exemple, Legendre et al., 1999, Bessa-Gomez et al., 2004). Enfin, en comparant les modèles de population qui incluent et n'incluent pas la structure par âge, Coulson et al. 2003) ont montré l'importance de considérer les caractéristiques du cycle de vie pour établir des prédictions fiables sur la dynamique et la persistance des populations structurées par classes d'âge.
Structure de population et histoire de vie chez le lézard vivipare
Après une présentation du modèle d’étude, nous décrirons dans un premier temps les effets observés de la structure de la population sur l’histoire de vie au sens large. Deuxièmement, nous nous concentrerons sur l’influence de la structure de la population sur le système d’accouplement et sur l’intensité de la sélection sexuelle.
Le lézard vivipare comme organisme modèle
Chez les jeunes femelles et les femelles âgées, la stratégie polyandre est favorisée (Richard et al., 2005), tandis que les femelles d'âge moyen sont plutôt monoandres. En effet, lors de l’accouplement, les mâles tiennent fermement les femelles en les mordant au niveau de l’abdomen (Fitze et al., 2005).
INFLUENCES DE LA STRUCTURE DE POPULATION SUR L’HISTOIRE DE VIE DU LÉZARD VIVIPARE
Les implications de nos travaux ne se limitent donc pas simplement au cas du fonctionnement des populations de lézards, mais s'inscrivent dans le cadre plus général de l'écologie des populations structurées. De plus, la capacité de manipuler les populations dans des conditions semi-naturelles offre une opportunité unique de relier de manière réaliste la structure de la population, le cycle biologique, la dynamique des populations et le risque d'extinction (par exemple Cote et al., 2007, Le Galliard et al. , 2005d, Lecomte et al. ., 2004).
Structuration en cohortes, environnement biotique et interactions sociales
À l’inverse, les effets de l’environnement biotique précédent sur l’intensité de la compétition intraspécifique restent largement inexplorés (mais voir Beckerman et al., 2003). Ces résultats montrent que les effets directs de l'environnement et ses effets indirects sur les interactions sociales le peuvent.
Densité dépendance de l’histoire de vie
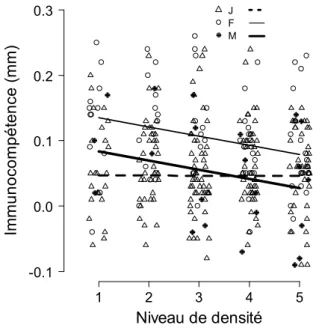
À l'échelle intraspécifique, la dépendance négative à la densité du cycle biologique peut être causée par (1) des interactions trophiques de type interférence et compétition via l'exploitation de la ressource et (2) des interactions sociales entre congénères (Edeline et al., 2010 et références citées). ). ). Effets sur le cycle de vie de l'interaction entre la densité de population et le rapport de masculinité des adultes.
Effets d’histoire de vie de l’interaction entre la densité de population et le sexe ratio adulte
Ainsi, nous nous attendons à ce que la survie et la croissance en taille des mâles et en taille à la naissance des juvéniles soient indépendantes de la densité des adultes et du sex-ratio. Nous nous attendons également à ce que le sex-ratio adulte ait un effet négatif sur la survie des femelles adultes et subadultes.
L’importance du sexe, de l’âge et de la famille dans les réponses individuelles à la structure de population
L'importance du sexe, de l'âge et de la famille dans les réponses individuelles à la structure de la population. Dans les deux cas, nous avons mis en évidence l’existence d’une variation entre les ménages dans la sensibilité du taux de croissance à l’environnement.
INFLUENCES DE LA STRUCTURE DE POPULATION SUR LE SYSTÈME D’APPARIEMENT ET L’INTENSITÉ DE LA SÉLECTION
EFFETS DE LA STRUCTURE DE LA POPULATION SUR LE SYSTÈME D'ADAPTATION ET L'INTENSITÉ DE LA SÉLECTION.
SEXUELLE CHEZ LE LÉZARD VIVIPARE
Ces résultats suggèrent que les stratégies d'accouplement et l'intensité de la sélection sexuelle seraient plus sensibles au nombre relatif de mâles présents dans la population. Ainsi, la gamme de densités étudiées n'était peut-être pas suffisante pour détecter les effets dépendants de la densité sur le système d'accouplement et l'intensité de la sélection sexuelle.
CONSÉQUENCES POUR LA DYNAMIQUE DES POPULATIONS DE LÉZARDS VIVIPARES
Pour étudier l'influence de la dépendance à la densité sur l'histoire de vie sur la dynamique déterministe des populations, nous avons effectué des analyses d'élasticité du taux de croissance λ des populations (modèle déterministe indépendant de la densité) et de la densité d'équilibre Neq (modèle déterministe dépendant de la densité). Une description complète des modèles est présentée au chapitre 2 pour l'influence de la structuration des populations en cohortes et au chapitre 5 pour l'influence de la densité de population sur la dynamique des populations de lézards.
Conséquences pour la dynamique déterministe des populations
Conséquences pour le risque d’extinction des populations
Notre expérience, combinée à l'analyse de la viabilité des populations, montre l'importance d'une dépendance négative à la densité dans les petites populations (Schoener et al., 2003). Ces réponses se traduisent par une aggravation du biais du sex-ratio chez les adultes et une forte augmentation du risque d'extinction de la population (Le Galliard et al., 2005d).
CONCLUSION
Cela devrait fortement influencer l’intensité de la compétition intra- et inter-cohortes et donc les trajectoires de vie et la dynamique des populations (Chapitre 1). Premièrement, quels sont les mécanismes proximaux de la réponse du cycle biologique à la structure de la population.
Maternal effects on offspring locomotion: Influence of density and corticosterone elevation in the lizard Lacerta vivipara. Male territoriality mediates density and sex ratio effects on oviposition in the zebrafish, Danio rerio.
ANNEXES
Caractérisation de la réponse au stress
Parallèlement, les animaux du groupe témoin ont été gardés dans des terrariums individuels. Les données de l'expérience de densité ont été analysées par des séances d'échantillonnage utilisant des modèles mixtes linéaires (Pinheiro & Bates, 2000).
Expérience Densité de population × sexe ratio adulte Etude réalisée par Marianne Mugabo & Jean-François Le Galliard
La croissance corporelle, la survie, la capacité de reproduction et l'immunocompétence ont été estimées de la même manière que pour l'expérience 2 (voir chapitre 5). Les analyses génétiques nécessaires aux attributions de paternité et à l'analyse du système d'appariement sont en cours (collaboration avec C. Acasuso, P. Federici et M. Richard).
Analyses de paternités
Les attributions de paternité ont été effectuées à l'aide du logiciel Cervus 3.0, comprenant 579 juvéniles viables, 40 juvéniles non viables, 30 œufs avortés et 50 œufs jaunes. Dans le cadre de l'expérience 4, 718 œufs ont été produits, dont 555 juvéniles viables, 54 juvéniles non viables, 51 œufs avortés et 58 œufs jaunes.
CHAPITRES
Effets cohortes dans des populations structurées et conséquences démographiques conséquences démographiques
Mécanismes des effets cohortes et leurs conséquences pour la dynamique de populations
RÉSUMÉ
Mechanisms of cohort effects and their consequences for population dynamics
ABSTRACT
Effets de l’environnement nutritif précoce sur l’histoire de vie dans des populations
Effets immédiats et décalés d’une privation de nourriture précoce sur l’histoire de vie chez un lézard non-longévif
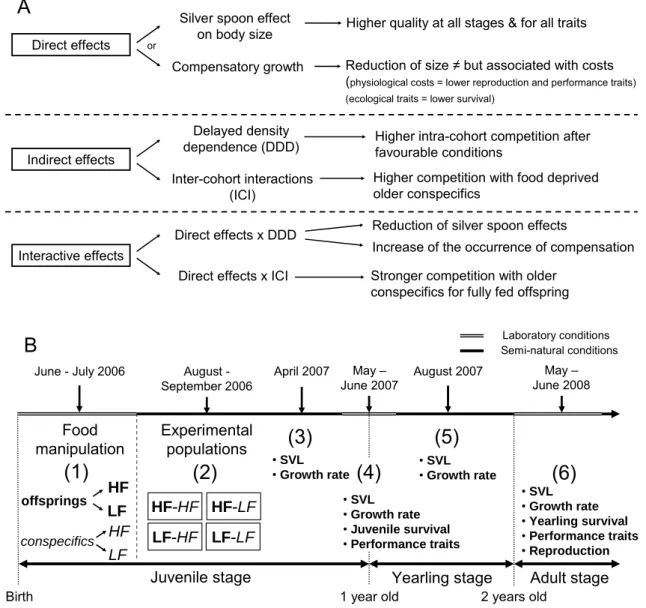
Effets immédiats et retardés d'une privation précoce de nourriture sur l'histoire de vie d'un lézard qui ne vit pas longtemps. La disponibilité alimentaire peut affecter les trajectoires de vie à travers trois mécanismes non exclusifs : (i) des effets immédiats et différés sur la qualité individuelle et des effets indirects différés sur les interactions sociales (ii) au sein des groupes et (iii) entre les cohortes.
Immediate and delayed life history effects caused by food deprivation early in life in a short-lived lizard
Food deprivation had direct immediate negative effects on growth, but positive long-term effects on immune competence. Food deprivation also had indirect lagged effects on growth, body size, early survival and reproduction, mediated by an interaction between its direct effects on individual quality and its lagged effects on the intensity of social interactions between cohorts, combined with the dependence on body size .
INTRODUCTION
Indeed, little is known about how the direct effects of early life food availability and the indirect effects mediated by social interactions combine to shape life trajectories (Beckerman et al., 2003). Here we report a study of the immediate and delayed effects of food availability on changes in lifespan in the common lizard (Zootoca vivipara Jacquin 1787), a short-lived reptile (3–4 years of life expectancy) with a plastic life history (Marquis et al., 2008).
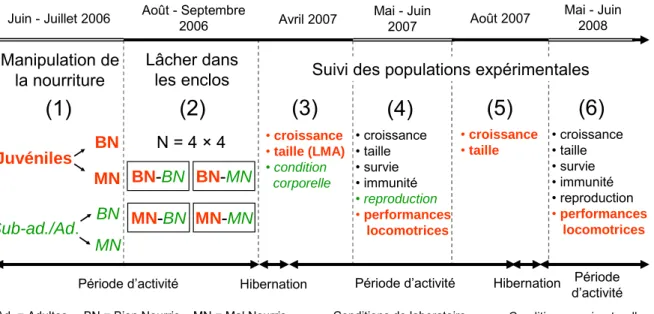
Measurements of life history traits
We generated populations of lizards by mixing offspring from the same food treatment group with yearlings and adults from a food-deprived or a fully fed treatment group following a fully crossed factorial design (Figure 1B). Four outdoor enclosures were randomly selected for each treatment group and initial populations had similar age and sex structure (4–5 adult females, 3 adult males, 4–6 year old females, 4–5 year old males, 28–34 juveniles, juvenile sex ratio = 51% of men ± 2 s.e.).
Measurements of performance traits
Statistical analyses
We therefore tested separately for the effects of treatments and the effects of body size on maximum sprint speed and endurance. Models included fixed effects of offspring food treatment, conspecific food treatment, sex and their first-order interactions, as well as additive effects of birth date and geographic origin and a random enclosure effect.
RESULTS
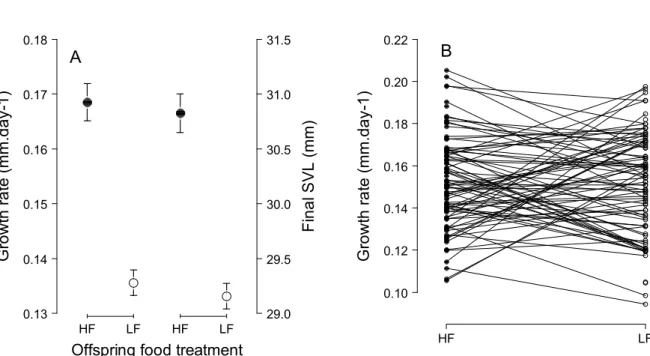
Immediate effects
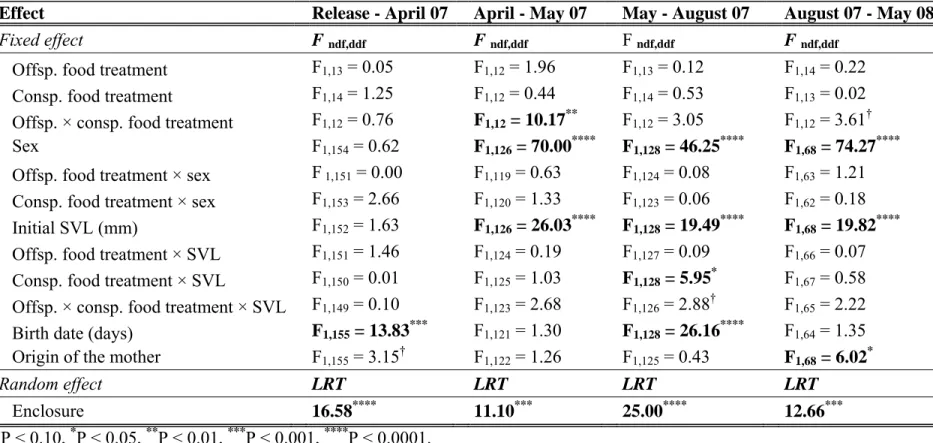
Analysis of growth rate of lizards (mm.day-1) during their first two years of life according to offspring food treatment, consp. food treatment, sex, initial SVL, birth date and maternal origin. The F and P values were obtained by backward selection from the full models and LRT values were obtained for the minimally adequate selected model.
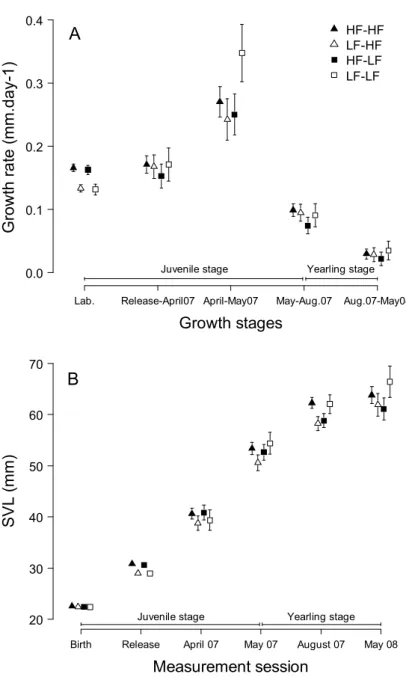
Delayed effects on body size
Differences in SVL between HF and LF lizards in April 2007 were of the same order of magnitude as after the laboratory manipulation (Fig. 3B), but were only marginally significant due to greater inter-individual variation (Table S1). Cohort density did not explain differences in growth rates, but partially explained differences in SVL.
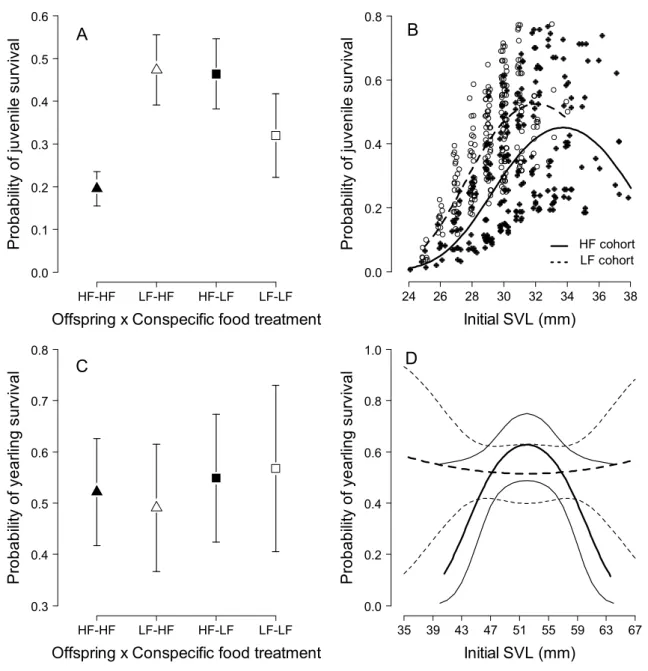
Delayed effects on survival
From May 2007 onwards, SVL was affected by an interaction between offspring and specific food treatment (Table S1). Post-hoc tests revealed that LF lizards grew to be larger than HF lizards when maintained with LF conspecifics, but grew to be smaller when maintained with HF conspecifics (Figure 3B, Table S1).
Delayed effects on reproductive performances
For a given fecundity, females from the HF offspring treatment group produced offspring on average 1 to 2 mm larger than those produced by LF females when kept with HF conspecifics, but the opposite was observed when they were maintained with conspecific LF (Figure 5B). However, if differences in fecundity are compensated after age 2, for example due to growth compensation, the λ of a population of HF offspring maintained with HF conspecifics would be significantly less than the λ of other treatment groups (Figure S2B).
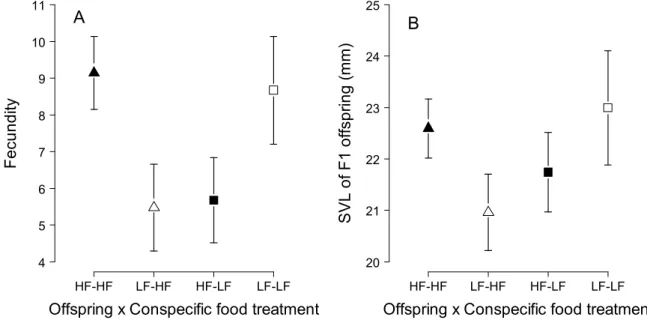
Delayed effects on performances traits
DISCUSSION
Immediate effects of food availability
In addition, unlike a previous study involving a smaller number of households (Le Galliard et al., 2005), we found that the sensitivity of growth rate to food availability varied significantly between households. Maternal (reviewed in Mousseau & Fox, 1998) and paternal (Galloway, 2001) effects may contribute to differences in family ranking across environments.
Long lasting effects of food availability on life-history traits
Here, fully fed lizards benefited from a silver spoon effect on female body size and reproduction when housed with fully fed yearlings and adults. Thus, the low survival of juveniles of fully fed offspring cannot be explained by selection against larger individuals.
Long-lasting effects of food availability on performance traits
Cohort interactions may therefore constrain the evolution of rapid growth rates early in life and should be considered in future studies of growth strategies in this and other species with structured populations. 2008, Cote et al., 2010). However, we found no evidence of direct survival benefits for higher immunocompetence in this study.
Conclusion
AKNOWLEGMENTS
Effect of maternal and paternal environments on seed characters in the herb Campanula americana (Campanulaceae). 1988) On the uses of lifetime reproductive success data. Experimental tests of intercohort competition for food and cover in the intertidal scallop (Oligocottus maculosus Girard).
SUPPORTING INFORMATION
Effets de l’environnement nutritif tardif sur l’histoire de vie dans des populations structurées en cohortes
Effets directs et sociaux de la disponibilité en nourriture tard dans la vie sur la variation d’histoire de vie chez un lézard non-longévif
Effets directs et sociaux de la disponibilité alimentaire en fin de vie sur la variation du cycle biologique d'un lézard qui ne vit pas longtemps. Ces populations expérimentales ont ensuite été suivies pendant deux ans, révélant les effets complexes de la nutrition sur les trajectoires de vie.
Direct and socially-mediated effects of food availability late in life on life- history variation in a short-lived lizard
In a manipulated cohort of juveniles, we previously demonstrated that food deprivation in the laboratory had direct immediate negative effects on structural growth but positive long-term effects on immune competence (Mugabo et al., 2010). At the same time, we manipulated the daily food consumption of 460 young lizards in the laboratory within 50 days of their birth (detailed procedures are described in Mugabo et al., 2010).
Measurements of life-history traits and immunocompetence
These models included fixed effects of (i) main food treatment interacted with pup food treatment to account for pup manipulation, (ii) second-order interactions with sex to test for sex differences in sensitivity to food treatment, (iii) initial age class ( year or adult) and first-order interactions with food treatment (iv), as well as additive effects of date at the start of the manipulation and geographic origin. Models included fixed effects of (i) major food treatment, juvenile food treatment, sex, initial age class and their first-order interactions, (ii) additive effects of initial SVL centered on its grand mean (linear and quadratic effects) to estimate selection on body size and (iii) additive effects of geographic origin and date at the start of the manipulation.
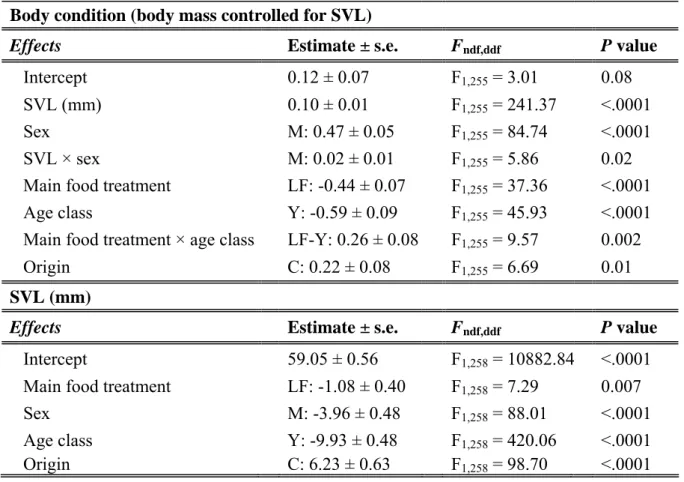
Immediate effects on morphology
Delayed effects on life history and immunocompetence Delayed effects on morphology
Lagged effects of main food treatment and juvenile food treatment on the probability of annual survival of females and males during (A) the first and (B) the second year of the experiment. Intergenerational effects of main food treatment and juvenile food treatment on (A) SVL at birth of offspring born in 2007 and 2008 and (B) on the annual growth rate and (C) annual survival probability of offspring born in 2007 during their first year of life .
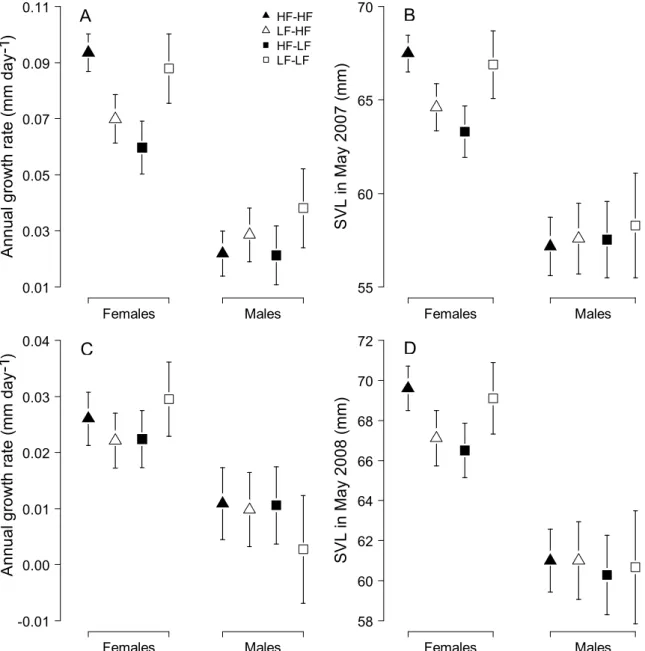
Immediate effects of food availability on morphology
Body size is also an important determinant of mating success in male common lizards (Fitze & Le Galliard, 2008). Therefore, male and female common lizards may experience similar selection for an optimal body condition and body size (Schwarzkopf, 1994), thus explaining their parallel flexibility with food availability.
Direct delayed effects of food availability on life history
This is surprising because female common lizards expend a lot of energy in vitellogenesis and pregnancy, and body size is important for their fertility, whereas male energy investment in gamete production is less important (Avery, 1975, Nagy, 1983). Socially mediated delayed effects of food availability on life history In the first year of a field experiment, annual growth rates and body size of females and.
Socially mediated delayed effects of food availability on life history During the first year of the field experiment, annual growth rates and body size of females and
When housed with food-deprived conspecifics (i.e., the LF-LF treatment group), both food-deprived aged females (this study) and food-deprived juveniles (Mugabo et al., 2010) exhibited excessive compensatory growth. This result differs from what was observed in the manipulated group of juveniles (Mugabo et al., 2010).
Direct delayed effects of food availability on immunocompetence In sharp contrast with life-history traits, immunocompetence was affected by lasting effects of
In juveniles, we previously hypothesized that competition with fully fed yearlings and adults prevented food-deprived juveniles from compensating for their limited growth early in life (Mugabo et al., 2010). Here we suggest that fully fed females were able to maintain a higher growth rate than food deprived females, even after the food manipulation, due to their higher competitive ability than juveniles (Lecomte et al., 1994).
Intergenerational effects of food availability
In fact, a food shortage in developing individuals can alter the makeup of the immune system, leading to a higher investment in immunity throughout life compared to individuals who experienced early good conditions (Birkhead et al., 1999, Tschirren et al., 2009). In adults and non-developing individuals, a decrease in energy resources is expected to inhibit immune responses (Martin et al., 2007).
ACKNOWLEDGEMENTS
Densité de population et interactions trophiques proies-prédateurs
Découvrir les effets de la taille des prédateurs et de la densité des proies sur la consommation de proies chez un lézard.
Dénouer les effets de la taille du prédateur et de la densité de proies sur la consommation de proies chez un lézard
Cependant, le meilleur modèle de réponse fonctionnelle a reçu un soutien modéré par rapport aux modèles allométriques plus simples basés uniquement sur la masse et le sexe des prédateurs. Les modèles de réponse fonctionnelle qui ignorent les variations individuelles sont susceptibles de déformer les interactions trophiques.
Disentangling the effects of predator body size and prey density on prey consumption in a lizard
In most species, variation in body size affects both foraging ability and predation risk (Aljetlawi et al., 2004, Tripet & Perrin, 1994, Werner & Hall, 1988). To incorporate this size dependence, feeding rates can be determined using allometric functions that incorporate predator and/or prey body size effects with simple power laws (e.g., Kooijman, 2000, Vucic- Pestic et al., 2010).

Lizard populations: enclosures
Prey populations
Estimation of feeding rates
Data analysis: feeding rates
These three basic functional response equations were modified to include more complex attack rates that include the effects of predator body mass and sex, and duration of solar irradiance. The total wi was calculated as the cumulative weight of all models including a particular variable or type of functional response (eg, Holling type I response) that is similar to the weight of the variable wj.
Data analysis: impact on invertebrate communities
We used the ADONIS procedures in the VEGAN package in R.2.10.0 (R Development Core Team 2009) with 999 permutations. Therefore, we entered stream proximity, a proxy for moisture, as an additional regression variable to explain invertebrate biomass and as a block in the ADONIS procedure.
Natural feeding rates
Models including M- and S-dependent attack rates were strongly supported (total wi > 0.99 and total wi = 0.98, respectively). In contrast to the results from the allometric group, functional response models including the I effect were supported (overall wi = 0.79).
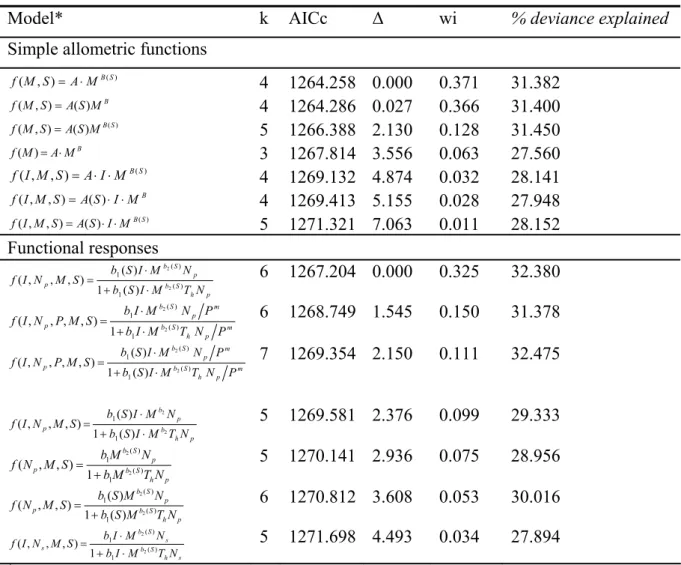
Impacts of lizard density on invertebrate populations
Because both sets of models used the same database, we can also compare them to determine the overall simplest model. The two main allometric functions including S and M ( Tables 2 and 3 ) were identified as the only models supported when combining the two sets of functions.
Intrinsic factors influencing feeding rates
Extrinsic factors influencing feeding rates and functional response functions
However, unlike previous studies (Miller et al., 2006, Schenk et al., 2005), we found only limited support for an effect of predator density. The common lizard is a non-territorial species with overlapping home ranges, and conspecific interference during predation may be rare (Massot et al., 1992).
Most parsimonious feeding rate functions
Densité dépendance de l’histoire de vie et risque d’extinction d’extinction
Histoire de vie densité-dépendante et la dynamique des petites populations
La dynamique de la population dépend dans une large mesure de la force et de la forme de la dépendance à la densité. Nous avons créé un gradient de densité faible à élevé dans de petites populations isolées de lézards vivipares (Zootoca vivipara) et étudié la forme de la dépendance de la densité au cours du cycle biologique sur un cycle d'un an.
Density-dependent life history and the dynamics of small populations
Unfortunately, the lack of reliable life history data often prevents making accurate demographic predictions about small populations regulated by density dependence. We created a gradient from low to high densities in small isolated populations of common lizards (Zootoca vivipara) and investigated the shape of density dependence of life history during an annual cycle.
RÉSUME