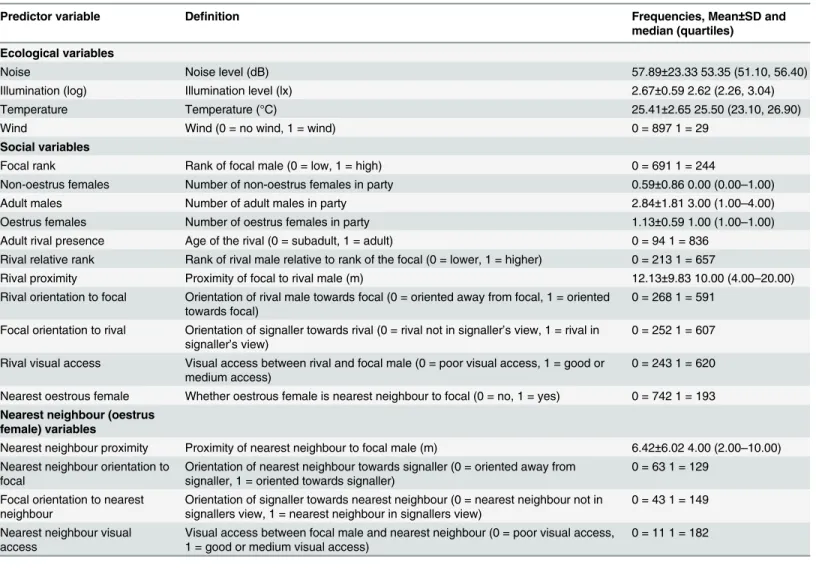
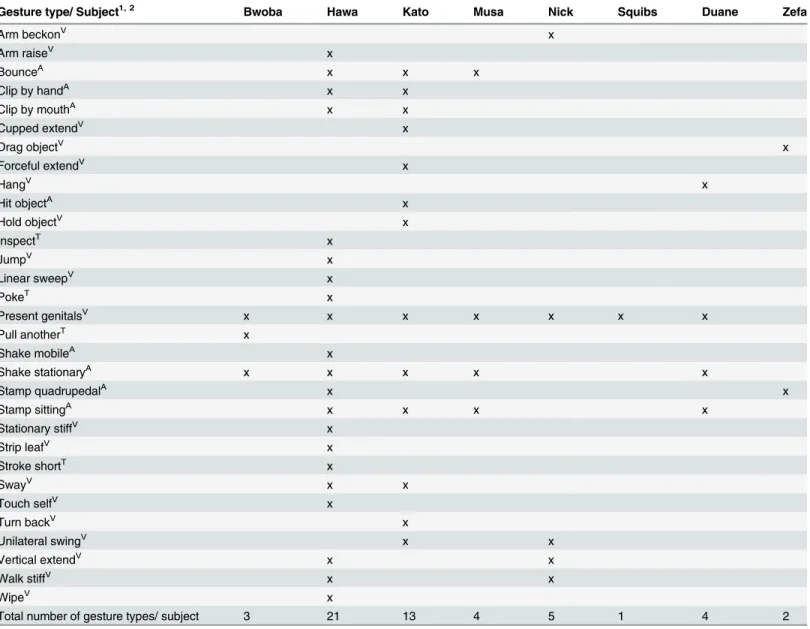
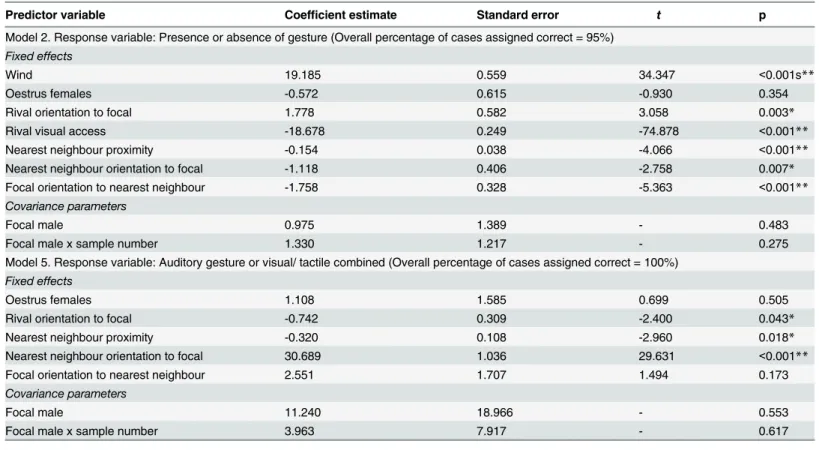
Gestural Communication and Mating Tactics in Wild Chimpanzees.
Texto
Imagem

Documentos relacionados
48,49 A study was designed to examine the hypothesis that minimally invasive combined endoscopic procedures were as good as or even better than surgical staging (mediastino- scopy)
São ainda estudadas outras marcas linguísticas dos actores: o «nós» equivalente à instituição que avalia a situação do menor (configurando o apagamento de um enunciador
When investigating the history of weight gain, young adult women with overweight in childhood and adolescence were more likely to have overweight in adulthood, according to the
When the chest X-rays were analyzed as to whether or not they presented the respective alterations, we found that the alveolar consolidation pattern was predominant and was present
When, in the familial cases, the probands and affected relatives were considered, t h e slight male predominance among the affected (24 males and 17 females) did not
When male worms were incubated with resorufin in this medium to which 2 mM calcium was added (and no EGTA), but which still lacked phosphate, the collecting ducts were labelled but
Não agregou prestígio; não tinha nada a ver com nosso leitor, que nunca soube do que se tratava aquela coluna; e não se encaixou na rotina de produção da Redação, regulada pela
From the 12 studies included (13 entries) in this meta-analysis, they all started from the same research assumption, in which the elderly with classifications according to the
