Effect of water chemistry on the planktonic communities and relationships among food web components across a freshwater ecotone
Texto
Imagem
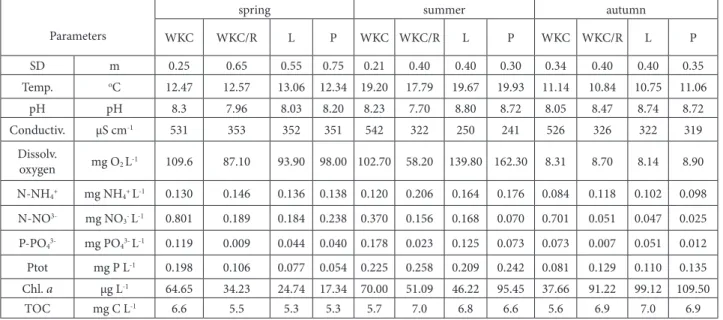
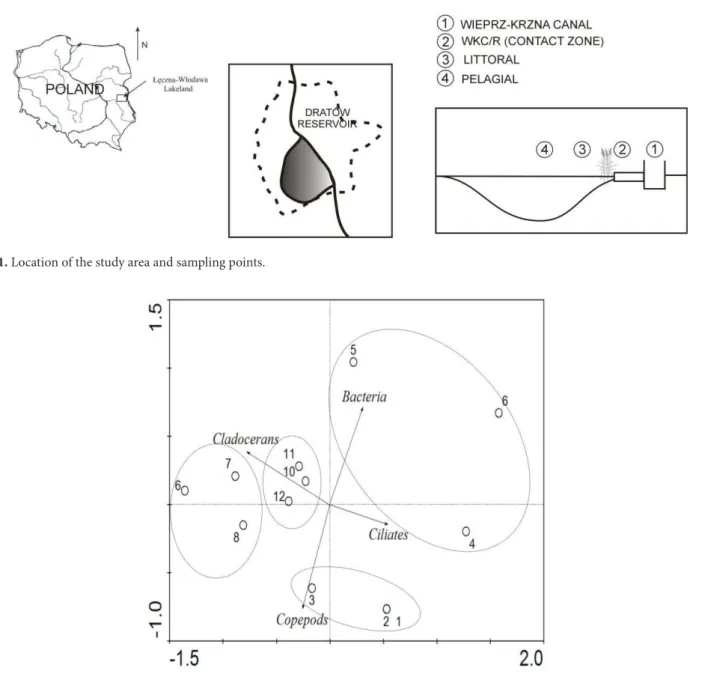
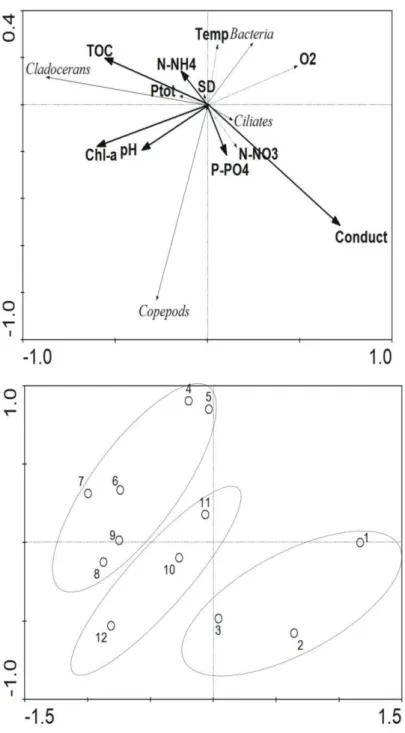
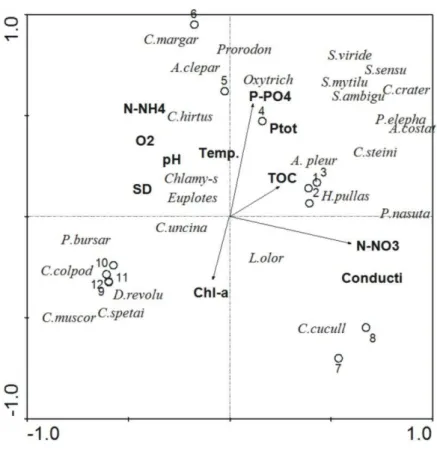

Documentos relacionados
The structural zones, shadowed zones or zones who suffer reflections were not considered as they are of difficult analysis using this scale.. The possibly
Para a realização de ensaios de medição do isolamento sonoro a sons de percussão entre compartimentos, de acordo com a NP EN ISO 16283-2, é necessária a existência de dois
1: scheme showing the distribution of different extracellular components in the three zones (external or peripheral, medial or intermediate and central or periovular) of
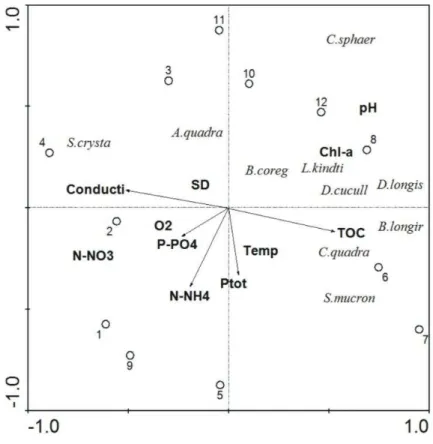
are to 1) document the trophic position of species of Chirostoma in relation to other fish species and food web components in Lake Chapala based on stable isotope data,
(2013) proposed the following procedure for selecting variables as input data for the Fuzzy C-Means algorithm. 1) Variables with spatial dependence and no significance at
A: BSE image of a single zoned crystal [reproduced from Vlach and Gualda (2007) with Elsevier permission]; clear zones are ferriallanite (clearest) and allanite; relatively
Como se pode verificar na figura 6, o conceito de 1PL refere-se às empresas cujas funções logísticas são totalmente verticalizadas, empresas que executam as suas próprias funções de
Y éstas han sido todas las actividades presentadas y trabajadas en el taller supramencionado; por algunas tuvimos que pasar “en volandas” debido a la escasez de tiempo destinado a
