Diffusivity in Searching Processes
E. P. Raposo1*, F. Bartumeus2, M. G. E. da Luz3, P. J. Ribeiro-Neto1, T. A. Souza1,4, G. M. Viswanathan5,6
1Laborato´rio de Fı´sica Teo´rica e Computacional, Departamento de Fı´sica, Universidade Federal de Pernambuco, Recife, Brazil,2Centre d’Estudis Avanc¸ats de Blanes, CEAB-CSIC, Blanes, Spain,3Departamento de Fı´sica, Universidade Federal do Parana´, Curitiba, Brazil,4Center for Polymer Studies and Department of Physics, Boston University, Boston, Massachusetts, United States of America,5Departamento de Fı´sica Teo´rica e Experimental, Universidade Federal do Rio Grande do Norte, Natal, Brazil, 6Instituto de Fı´sica, Universidade Federal de Alagoas, Maceio´, Brazil
Abstract
Theoretical and empirical investigations of search strategies typically have failed to distinguish the distinct roles played by density versus patchiness of resources. It is well known that motility and diffusivity of organisms often increase in environments with low density of resources, but thus far there has been little progress in understanding the specific role of landscape heterogeneity and disorder on random, non-oriented motility. Here we address the general question of how the landscape heterogeneity affects the efficiency of encounter interactions under global constant density of scarce resources. We unveil the key mechanism coupling the landscape structure with optimal search diffusivity. In particular, our main result leads to an empirically testable prediction: enhanced diffusivity (including superdiffusive searches), with shift in the diffusion exponent, favors the success of target encounters in heterogeneous landscapes.
Citation:Raposo EP, Bartumeus F, da Luz MGE, Ribeiro-Neto PJ, Souza TA, et al. (2011) How Landscape Heterogeneity Frames Optimal Diffusivity in Searching Processes. PLoS Comput Biol 7(11): e1002233. doi:10.1371/journal.pcbi.1002233
Editor:Mercedes Pascual, University of Michigan and Howard Hughes Med. Inst., United States of America ReceivedJune 1, 2011;AcceptedSeptember 2, 2011;PublishedNovember 3, 2011
Copyright:ß2011 Raposo et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding:This work was supported by CNPq, CAPES, F. Arauca´ria, FAPEAL, FACEPE (Brazilian agencies). FB acknowledges the Spanish Ministry of Science and Innovation (Grants RyC09-449-06-01, BFU2010-22337). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing Interests:The authors have declared that no competing interests exist. * E-mail: [email protected]
Introduction
The random search problem has lately received a great deal of attention [1,2]. This is partly due to its broad interdisciplinary range of applications, which include, e.g., enhanced diffusion of regulatory proteins while ‘‘searching’’ for specific DNA spots [3,4] and the finding of binding sites on transmembrane proteins by neurotrans-mitters in the brain [5]. Recently, this problem has also found interesting connections with human mobility and related topics [6–9]. A classical context in which the random search problem has been applied in the last four decades is animal foraging [1,2,10– 27], with the searcher (i.e. forager) typically represented by an animal species in quest of target sites (prey, food, other individuals, shelter, etc.) in a search landscape.
Among the most studied random walk models proposed as plausible search strategies, we cite correlated random walks [12,28,29], Le´vy flights and walks [13–17,19,20,24,25,27,30–39], intermittent walks [40–46], and composite Brownian walks [47,48]. In particular, Le´vy random searchers, with probability distribution of step lengthsp(‘)*‘{m, for1vmƒ3, have successfully explained [34] the emergence of optimal searches in landscapes with randomly and scarcely distributed target sites. On the other hand, when resources are plentiful Le´vy strategies are unnecessary [34], and efficient Brownian optimal searches may arise with, e.g., a Poisson-like exponential distribution p(‘) [24,25]. Le´vy flights and walks have been also shown to be relevant in several other contexts [1], such as in proteins searching for specific DNA sites [49], in which the optimal Le´vy mechanism emerges directly from the underlying physics of the problem (polymer scaling theory in three dimensions).
In the regime of low density of resources of the random search problem, two limiting situations have been extensively considered [34]: (i) non-destructive searches, in which the searcher always departs from a position at the vicinity of the last target found with unrestricted revisits; and (ii) destructive searches, in which, once found, the target becomes inaccessible to future visits, so that the starting point of the searcher is, on average, faraway from all targets. In the former case the maximum efficiency is achieved [34] for mopt,nd&2 (a ‘‘compromise’’ superdiffusive solution), whereas in the lattermopt,d?1(ballistic motion). It is important to observe, nevertheless, that by varying the searcher’s starting point [44,48] or the degree of target revisitability or temporal regeneration [50,51], intermediate values of the optimal Le´vy exponent arise,1vmopt*v2.
It is also interesting to comment on the effect of an energy cost function on the efficiency of search strategies. Indeed, as reported in [50,51], the range ofm-values associated with search paths in which the net energy gain (the balance between the energy income due to the finding of targets and the energy cost of the search process itself) remains always positive is actually limited. In such a case, low values ofmgiving rise to very large search jumps might not be acceptable, since they imply a high energy cost, with intermediate values ofmopt
emerging as the best strategy. In addition, we also refer to the study reported in [52] in which exact results for the first passage time and leapover statistics of Le´vy flights are presented. In this case, the targets might not be always detected, being thus overshoot by jumps whose length distribution displays infinite variance.
A particularly important one is to understand the coupling mechanism between landscape spatiotemporal dynamics and efficient search motility, when resources are scarce and environ-mental information is limited. In this sense, the pervasiveness of different animal search strategies is expected to strongly depend on a few but essential features of actual landscapes. For instance, targets distributions in realistic search processes usually present heterogeneous properties through time and space, such as diverse degrees of temporal regeneration and spatial aggregation [26,53,54]. Although the effect of (global) resource density on animal foraging behavior is well documented [25,26,37,42,55], much less is known about how spatiotemporal landscape heterogeneity dynamics affects the target revisitability and/or searcher-to-targets distances, both known to be key properties to optimize perception-limited searches [44,48,50,51]. Thus, a mechanistic understanding of how and which landscape features are related to search efficiency should be a relevant step towards a comprehensive view of animal foraging behavior.
Here we address the question of how the landscape heterogeneity influences the encounter success and search efficiency under conditions of constant (global) density of scarce resources. We develop a random search model in which diverse degrees of inhomogeneities are considered by introducing fluctuations in the starting distances to target sites. We thus ask what happens to the optimal search strategy in an heterogeneous landscape, as the searcher’s initial distances to the targets fluctuate along the search. We answer to this query qualitatively for the general case and quantitatively for Le´vy random searches in particular, in the constant density regime of scarce resources. In patchy or aggregated landscapes, we find that enhanced diffusivity (including superdiffusive strategies) favors the encounter of targets and the success of foraging. Eventually, for strong enough fluctuations in the starting distances to nearby targets a crossover to ballistic strategies might emerge.
These predictions are empirically testable through feasible experiments which investigate the dynamics (e.g. diffusion exponent) of foraging organisms in specially designed low-density environments of controlled heterogeneity.
Materials and Methods
Distributions of starting positions: General considerations We consider a random search model in which diverse degrees of landscape heterogeneity are taken into account by introducing
fluctuations in the starting distances to target sites in a one-dimensional (1D) search space, with absorbing boundaries separated by the distancel. Every time an encounter occurs the search resets and restarts over again. Thus, the overall search trajectory can be viewed as the concatenated sum of partial paths between consecutive encounters. The targets’ positions are fixed – targets are in fact the boundaries of the system. Fluctuations in the starting distances to the targets are introduced by sampling the searcher’s departing position after each encounter from a probability density function (pdf) p(x0) of initial positions x0. Importantly,p(x0) also implies a distribution of starting (a)sym-metry conditions regarding the relative distances between the searcher and the boundary targets.
This approach allows the typification of landscapes that, on average, depress or boost the presence of nearby targets in the search process. Diverse degrees of landscape heterogeneity can thus be achieved through suitable choices ofp(x0).
For example, a pdf providing a distribution of nearly symmetric conditions can be assigned to a landscape with a high degree of homogeneity in the spatial arrangement of targets. In this sense, the mentioned destructive search represents the fully symmetric limiting situation, with the searcher’s starting location always equidistant from all boundary targets. On the other hand, a distributionp(x0)which generates a set of asymmetric conditions is related to a patchy or aggregated landscape. Indeed, in a patchy landscape it is likely that a search process starts with an asymmetric situation in which the distances to the nearest and farthest targets are very dissimilar. Analogously, the non-destructive search corresponds to the highest asymmetric case, in which at every starting search the distance to the closest (farthest) target is minimum (maximum). Finally, a pdfp(x0)giving rise to an heterogeneous set of initial conditions (combining symmetric and asymmetric situations) can be associated with heterogeneous landscapes of structure in between the homogeneous and patchy cases.
More specifically, the limiting case corresponding to the mentioned destructive search can be described by the pdf with fully symmetric initial condition,
p(x0)~d(x0{l=2), ð1Þ where d(x0) denotes Dirac d-function. This means that every destructive search starts exactly at half distance from the boundary targets. In this context, it is possible to introduce fluctuations inx0 by considering, e.g., a Poisson-like pdf [56] exponentially decaying with the distance to the point at the center of the search space, x0~l=2:
p(x0)~Aexp½{(l=2{x0)=a, ð2Þ wherervƒx0ƒl=2, withrvthe ‘‘radius of vision’’ of the searcher (see below),Athe normalization constant, andp(x0)~p(l{x0)
due to the symmetry of the search space.
On the other hand, the highest asymmetric non-destructive limiting case is represented by
p(x0)~d(x0{rv), ð3Þ so that every search starts from the point of minimum distance in which the nearest target is undetectable, x0~rv. Similarly, fluctuations in x0 regarding this case can be introduced by considering a Poisson-like pdf decreasing with respect to the point x0~rv:
Author Summary
p(x0)~Bexp½{(x0{rv)=a, ð4Þ where rvƒx0ƒl=2, B is a normalization constant, and
p(x0)~p(l{x0). In Eqs. (2) and (4), the parameter a controls the range and magnitude of the fluctuations. Actually, the smaller the value of a, the less disperse are the fluctuations around x0~l=2andx0~rv in Eqs. (2) and (4), respectively.
Random search model in 1D
When looking for boundary target sites in a 1D interval, the searcher’s step lengths‘are taken from ageneralpdfp(‘). At each step the probabilities to move to the right or to the left are equal. We define the ‘‘radius of vision’’rvas the distance below which a target becomes detectable by the searcher. Thus, if the targets are located at the boundary positionsx~0andx~l, the search keeps on as long as the walker’s position lies in the rangervƒxƒl{rv. Here we are interested in searches in environments scarce in targets, i.e. forl&rv. In this case, leaving the present position to look randomly for targets should occur much more frequently than simply detecting a site in the close vicinity, a regime favored when targets are plentiful.
Suppose initially that, as a target is found, the search always restarts from the same positionx0in the intervalrvƒx0ƒl{rv. As discussed, the highest asymmetric (non-destructive) and fully symmetric (destructive) cases correspond respectively to setting x0~rv (orx0~l{rv, due to symmetry) andx0~l=2. After the encounter of a statistically large number of targets, the efficiency of the search,g, is evaluated [34] as the ratio of the number of sites found to the total distance traversed by the searcher. Since this distance is equal to the product of the number of encounters and the average distance traveled between consecutive findings, SLTp(x0), theng(x0)~1=SLTp.
Consider now that, instead of always departing from the same location after an encounter, the searcher can restart from any initial positionx0in the rangervƒx0ƒl{rv, chosen from a pdf
p(x0). The fluctuating values of x0 imply a distribution ofSLTp values. Since searches starting atx0are statistically indistinguish-able from searches starting atl{x0(in both cases the closest and farthest targets are at distances x0 and l{x0 from the starting location), the symmetry of the search space regarding the position x~l=2impliesSLTp(x0)~SLTp(l{x0). The average efficiency thus becomes
g~1=SLTp~1= 2 ðl=2
rv
SLTp(x0)p(x0)dx0 !
, ð5Þ
wherep(x0)~p(l{x0)due to the above mentioned symmetry. To study the effect of fluctuations in the starting distances of a searcher, we note that the exact average distanceSLTp(x0)in Eq. (5) can be formally expressed [57,58] as
SLTp(x0)~½(I{L)
{1Sj‘jT(x0), ð6Þ
where the integral operatorLacts as follows:
½LSj‘jT(x0)(x)~
ðl{rv
rv
p(x{x0)Sj‘jT(x0)dx0, ð7Þ
andI and Sj‘jT(x0)are, respectively, the unity operator and the average length of a single step starting atx0. Specifically, we can write for a general pdfp(‘)
Sj‘jT(x0)~(x0{rv)
ð{(x0{rv)
{?
p(‘)d‘z
ð{rv
{(x0{rv) j‘jp(‘)d‘
z
ðl{rv{x0
rv
j‘jp(‘)d‘z(l{rv{x
0 )
ð?
l{rv{x0
p(‘)d‘:
ð8Þ
The second and third integrals above represent steps to the left and to the right which are not truncated by the encounter of a target site at the boundaries; the first and last ones concern steps truncated by the detection of the targets at x~0 and x~l, respectively (what actually happens atx~rvandx~l{rv, due to the searcher’s ‘‘radius of vision’’).
Despite the formal aspect of Eq. (6), the numerical calculation of SLTp(x0) with a given p(‘) can be performed by discretizing [57,58] the search interval rvƒxƒl{rv, i.e. x~jdx, with j integer anddx%l. In this procedure, integrals are approximated
by summations, and so on.
In the next section, we use this model to study the role of landscape heterogeneity on the search efficiency and diffusivity. The presented analysis is qualitative for the general case and quantitative for Le´vy random searches.
Results
Efficient search strategies with a general pdf of step lengths
Consider, first, the limiting case with no fluctuation in the starting distances. The underlying mechanisms of efficient searches with asymmetric and symmetric initial conditions are fundamen-tally distinct. In the fully symmetric (destructive) case(x0~l=2)
the closest sites are located at equal initial distancesl=2&rvfrom the searcher in the low-density regime. Thus, for a general distribution of step lengths p(‘) characterized by a set of parametersfkig, the one(fkigs)that leads to the largest efficiency
gmust present the fastest possible diffusivity in order to reach these faraway targets. For example, in the case of the single-parameter power-law pdfp(‘)*‘{m, gis maximized with ballistic strategy [34]:k1, s~mopt,s?1.
In contrast, in the highest asymmetric (non-destructive) situation
(x0~rvorx0~l{rv)the most efficient search must compromise between performing large steps to access the farthest site and sweeping in detail at the vicinity of the closest site. In the parameter space, this solution, related to a set fkigas, displays intermediate diffusivity between normal (Brownian) and the fastest possible one, assigned to the setfkigs. In the same example, this implies [34]k1, as~mopt,as&2, in contrast with Brownian diffusion resulting frommw3(see Figs. 1 and 2).
When the starting positions are not fixed, heterogeneous landscapes with stronger fluctuations in the distances to nearby targets lead to optimal search strategies with faster dynamics (enhanced diffusivity). The arguments giving rise to this general conclusion are as follows.
On one hand, sampling starting positions around x0~l=2
corresponds to introduce fluctuations in the initial distances to the faraway boundary targets in the low-density regime, as discussed. In this case, we expect that starting positions far away from x0~l=2 are chosen with smaller probabilities. This implies a decreasing pdfp(x0) from l=2 torv, such as found in Eq. (2). Consequently, both p(x0) and SLTp(x0) increase monotonically from rv to l=2(Fig. 3). The most relevant contribution to the productSLTp(x0)p(x0)in Eq. (5) thus comes from positions near
SLTp and SLTp(x0~l=2), indicating that searches with fully symmetric (fixed) initial condition and those comprising fluctua-tions in the faraway targets present similar optimal dynamics, related to the setfkigs, namely ballistic, if supported byp(‘).
On the other hand, in the asymmetric case fluctuations in the starting distances to the nearby boundary target can be introduced by a decreasing pdf p(x0) from rv to l=2, such as in Eq. (4). Therefore, asSLTp(x0)increases andp(x0)diminishes, the initial position associated with the most relevant contribution to SLTp(x0)p(x0) in Eq. (5) crosses over to somewhere in between
x0~rvand x0~l=2. Indeed, the slowerp(x0)decays, the larger such position becomes. As a consequence, the asymmetric optimal setfkigasin the absence of fluctuations might give away the role of the most efficient search strategy to some other intermediate compromising solutionfkigint, which is closer to the symmetric set fkigs in the parameter space and, therefore, presents enhanced dynamics (e.g., a larger diffusion exponent). Eventually, for some proper choice ofp(x0)encompassing strong fluctuations with large weight near x0~l=2, the justification for such compromising solution might even fade away, so that fkigint?fkigs, with strategies of fastest possible diffusivity becoming optimal. In this uttermost case fluctuations lose their local character, and a crossover from superdiffusive to ballistic search behavior may take place.
We observe that the above rationale should also apply, at least qualitatively, to searches in higher-dimensional spaces. In this situation, as the search path can be approximated by a sequence of nearly rectilinear moves, the general qualitative features of 1D random searches usually hold true in higher dimensions [34,39]. Nevertheless, the finding of targets in 2D and 3D occurs with considerably lower probability, since the extra spatial directions yield a larger exploration space, resulting in lower encounter rates and search efficiencies. The impact of target spatial fluctuations on high-dimensional search strategies should also reduce [39]. We can thus conclude that, beyond representing the realistic exploration space of some animal species [27], the 1D analysis presented here is also useful in establishing upper limits for the influence of landscape heterogeneities in random searches. Therefore, the understanding of animal foraging behavior in 2D and 3D, as well as other practical realizations of the random search problem, might also benefit from the present results.
We next apply the above arguments, valid for a general pdfp(‘), to the particular case of Le´vy random searchers.
Le´vy searches in heterogeneous landscapes
We now specifically consider a random searcher with step lengths chosen from the pdf
p(‘)~(m {1)‘m{1
0 2
1
j‘jm, j‘j§‘0, ð9Þ
andp(‘)~0otherwise, with‘0representing a lower cutoff length. We assign a ‘‘negative step length’’ (j‘j~{‘) if the searcher moves to the left and take‘0~rv for simplicity. Equation (9) for
1vmƒ3corresponds to the long-range asymptotical limit of Le´vy
a-stable distributions with index a~m{1, characterized by the Figure 1. Robustness of the ballistic optimal search strategy
with respect to fluctuations in the distances to faraway target
sites.In the case of Le´vy random searchers, forl~103andr
v~1, the
average search efficiencyg, Eq. (5), is always highest form?1(ballistic
dynamics), for any value of the parameter a of the Poissonian fluctuations around the maximum allowed distance,x0~l=2, Eq. (2). Cases with uniform and without any (d-function) fluctuation are also shown. Solid lines are a visual guide.
doi:10.1371/journal.pcbi.1002233.g001
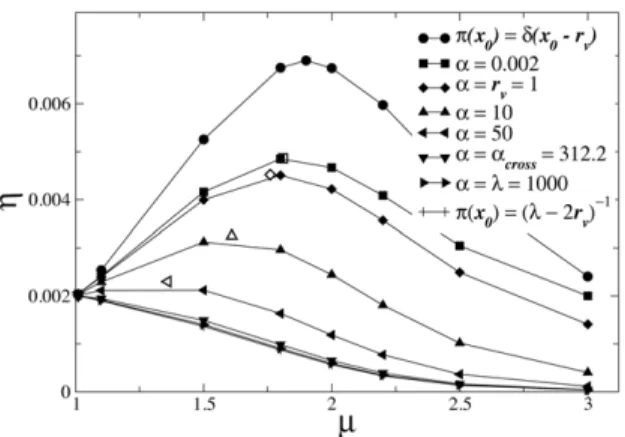
Figure 2. Shift in the optimal search strategy towards an enhanced superdiffusive dynamical regime, as landscapes with
distinct degrees of heterogeneity are considered. For Le´vy
random searchers, usingl~103andr
v~1(solid symbols), the average
search efficiency g, Eq. (5), is maximized for smallermopt(a) (faster diffusivity) in the case of wider (larger-a) Poissonian fluctuations in the distances to nearby target sites, Eq. (4). Cases with uniform and without any (d-function) fluctuation are also shown (solid lines are a visual guide). Empty symbols locate the maximum g obtained from the conditionf(m~mopt,a)~LSLTp=Lmjm~mopt~0. For strong enough
fluc-tuations, withawacross&312:2, a crossover to ballistic dynamics (m?1) emerges.
doi:10.1371/journal.pcbi.1002233.g002
Figure 3. Nice adjustment of the average distance SLTp
traversed between consecutive findings by a Le´vy random
searcher starting at position x0. Results obtained by numerical
discretization of Eq. (6) (solid lines) and multiple regression (symbols), forl~103andr
v~1.
existence of rare, large steps alternating between sequences of many short-length jumps [13,14,16]. As its second moment diverges the central limit theorem does not hold, and anomalous (superdiffusive) dynamics governed by the generalized central limit theorem takes place. Indeed, Le´vy random walks and flights are related to a Hurst exponent [13,14] Hw1=2, with ballistic dynamics in the casem?1, whereas diffusive behavior (H~1=2) emerges formw3. Formƒ1pdf (9) is not normalizable andm~2
corresponds to the Cauchy distribution.
The search path eventually comprises truncated steps due to the encounter of targets, so that the power-law decay of Eq. (9) cannot extend all the way to infinity, thus implying an effective truncated Le´vy distribution [59]. In spite of this, in the regime l&rv the search should retain the most relevant properties of a non-truncated Le´vy walk to a considerable extent. Indeed, the ratiorof the number of truncated steps to the non-truncated ones, essentially equal to the inverse of the average number of steps performed between consecutive targets, is given by r*(l=rv)(1{m)=2 and r*(l=rv)1{m, for l&rv, in the highest asymmetric (non-destructive) and fully symmetric (destructive) cases, respectively [34,57,58]. Thus, except for m?1 ballistic walks, one has that r%1 if l&rv. Further, the justification for truncated distributions also arises naturally in the context of animal foraging since directional persistence due to scanning is likely to be broken at the finding of targets [19]. Indeed, infinitely long rectilinear paths are not allowed for searching organisms.
By inserting Eq. (9) into Eqs. (6) and (7), we numerically calculateSLTp(x0)through the discretization of the search space (see previous section). Results are displayed in solid lines in Fig. 3. Notice first the presence of the symmetrySLTp(x0)~SLTp(l{x0) discussed above. In the absence of fluctuations in the initial distances, the existence of a maximum efficiency with an intermediate exponentmopt,as&2(see Fig. 2) for searches starting at fixedx0~rv(highest asymmetric condition) can be understood as follows: strategies withm?1might access the farthest target at x~lin a ballistic way after a small number of very large steps, implying a large SLTp(x0~rv) and low efficiency; in contrast, searches withm?3tend on average to find the closest site atx~0 after a great number of small steps, also giving rise to a large SLTp(x0~rv); the efficient compromise between these two trends, leading to the lowestSLTp(x0~rv)and maximumg, is therefore represented by a strategy with an intermediate value,mopt,as&2.
In the presence of fluctuations in the starting distances, the integral (5) must be evaluated. Although the explicit expression for SLTp(x0), Eq. (6), is not known up to the present, a multiple regression can be successfully performed,
SLTp(x0)~ XNx
i~0
X Nm
j~0
aijxi0mj, ð10Þ
as indicated by the nice adjustment shown in Fig. 3, obtained with Nx~10andNm~8. Thus, the integral (5) can be done using Eqs. (2), (4) and (10), with results displayed in Figs. 1 and 2 for several values of the parametera.
By considering fluctuations in the starting distances to faraway targets through Eq. (2), we notice in Fig. 1 that the efficiency is qualitatively similar to that of the fully symmetric condition, Eq. (1), in agreement with the general arguments of the previous section. Indeed, in both cases the maximum efficiency is achieved as m?1. For 1vmv3 the presence of fluctuations only slightly improve the efficiency. These results indicate that ballistic strategies remain robust to fluctuations in the distribution of faraway targets.
On the other hand, fluctuations in the starting distances to nearby targets, Eq. (4), are shown in Fig. 2 to decrease considerably the search efficiency, in comparison to the highest asymmetric case, Eq. (3). In this regime, since stronger fluctuations increase the weight of starting positions far from the target at x~0, the compromising optimal Le´vy strategy displays enhanced superdiffusion, observed in the location of the maximum efficiency in Fig. 2, which shifts frommopt&2, for the delta pdf and Eq. (4) with smalla, towardsmopt?1, for largera(slower decayingp(x0)). Indeed, both the pdf of Eq. (4) with a vanishingaand Eq. (3) are very acute atx0~rv. It is also worth noticing that a lowermoptis related to a larger Hurst exponent [1,13,14], and therefore to a larger diffusion exponent, as argued in the previous section.
As even larger values of aare considered, fluctuations in the starting distances to the nearby target become non-local, and Eq. (4) approaches thea??limiting case of the uniform distribution,
p(x0)~(l{2rv)
{1 (see Fig. 2). In this situation, search paths
departing from distinctx0are equally weighted in Eq. (5), so that the dominant contribution to the integral (and to the average efficiencygas well) comes from search walks starting at positions nearx0~l=2. Since for these walks the most efficient strategy is ballistic, a crossover from superdiffusive to ballistic optimal searches emerges, induced by such strong fluctuations. Conse-quently, the efficiency curves for very large a (Fig. 2) are remarkably similar to that of the fully symmetric case (Fig. 1).
We can quantify this crossover shift in mopt by defining a functionmopt(a) that identifies the location in them-axis of the maximum in the efficiencyg, for each curve in Fig. 2 with fixeda. As discussed, eventually a compromising solution withmopt(a)w1
cannot be achieved, and an efficiency functiongmonotonically decreasing with increasingmarises forawacross. In this sense, the valueacrossfor which such crossover occurs marks the onset of a regime dominated by ballistic optimal search strategies.
The value of mopt for each a can be determined from the condition f(m~mopt,a)~LSLTp=Lmjm~mopt~0, so that, by con-sidering Eqs. (4), (5) and (10),
f(m,a)~2AX
Nx
i~0
X Nm
j~0 aijjmj
{1
Xi
k~0 i!akz1
(i{k)! e {arvri{k
v {e {al=2 l
2
i{k!
" #
( )
,
ð11Þ
with A~f2a½exp({rv=a){exp({l=(2a))g {1
. Solutions are displayed in Fig. 4 and also in Fig. 2 as empty symbols, locating the maximum of each efficiency curve. In addition, the crossover value can be determined throughf(m?1z,a~across)~0. In the case of pdf (4), we obtain (Fig. 4)across&312:2forl~103 and rv~1(regimel&rv).
We also note that the scale-dependent interplay between the target density and the range of fluctuations implies a value ofacross which is a function ofl. For instance, a largerl(i.e., a lower target density) leads to a largeracross and a broader regime in which superdiffusive Le´vy searchers are optimal. Nevertheless, the above qualitative picture should still hold as long as low target densities are considered.
Moreover, since ballistic strategies lose efficiency in higher dimensional spaces [44], it might be possible that in 2D and 3D the crossover to ballistic dynamics becomes considerably limited. In spite of this, enhanced superdiffusive searches, with
From these results we conclude that, in the presence of Poissonian-distributed fluctuating starting distances with
aƒacross, Le´vy search strategies with faster (enhanced) super-diffusive properties, i.e. 1vmoptvm opt,as&2, represent optimal compromising solutions. In this sense, as local fluctuations in nearby targets give rise to landscape heterogeneity, Le´vy searches with enhanced superdiffusive dynamics actually maximize the search efficiency in aggregate and patchy environments. On the other hand, for strong enough fluctuations with awacross, a crossover to the ballistic strategy emerges in order to access efficiently the faraway region where targets are distributed. These findings are in full agreement with the general considerations discussed in the previous section.
At last, to further test the robustness of these results we have also considered the power-law distribution of starting positions,
p(x0)~Cx{n
0 , with rvƒx0ƒl=2, p(x0)~p(l{x0), and C as the normalization constant. Differently from distributions (2) and (4), the long tail in this pdf confers self-affine scale-invariant properties over a long spatial range in the low-density regime,
l&rv. The evidence of scale-free distributions of targets has been reported in the context of animal foraging, e.g. in [24]. In the present analysis we have essentially verified all the general features previously discussed. In particular, all strategies with
nvncross&0:35 are ballistic, with compromising superdiffusive solutions arising forn§ncross.
Discussion
The effect of limited resources on animal motility is well documented in ecology. Scarcity coming from resource competi-tion is known to induce higher dispersal rates [60,61] and larger home ranges [62,63]. Habitat fragmentation also reshapes dispersal kernels, often increasing dispersal distances [64]. In the context of foraging behavior, the role of (global) resource density has been considerably investigated, with strong evidence pointing to shifts from Brownian to superdiffusive search strategies as animals move from high to low productive areas. Examples range from microorganisms [37] to large marine predators [25,26,55]. In
contrast, much less is known about the influence of heterogeneity in the resource distribution on the foraging success.
Most theoretical efforts relying on core random search theory have by far provided only a limited approach to the issue of optimal searches, since they mostly assume oversimplified landscapes [2,40]. Nonetheless, a few simulation studies have addressed the effect of environmental heterogeneity, including target motion, on encounter success for different searcher types [19,24,39,65,66]. These works give support to the hypothesis that search processes are linked to target distributions and dynamics, thus agreeing with our results in that the optimal strategy can actually change, e.g. from superdiffusive to ballistic motion, depending on the landscape heterogeneity. In a more recent example, it was shown [65] that Le´vy optimal foragers can be evolutionarily optimal in heterogeneous environments, for suitable details of the simulations and definition of efficiency. Our work advances on this topic by pinpointing a very general mechanism which seems essential to understand previous simulation results [19,24,39,65].
By comprehensively describing the key mechanism coupling landscape dynamics and search diffusivity, we have shown that statistical fluctuations in the set of initial search conditions play a crucial role for determining which strategy is optimal. The presence of such fluctuations sets a clear basis for the non-universality of search patterns, and shows that enhanced diffusivity (including superdiffuse strategies) favors random encounter success in patchy and aggregated landscapes. As a consequence, the foraging conditions in which Le´vy strategies appear as optimal are much broader than previously suggested [40,44–46].
In dynamic and complex landscapes with scarcity of resources neither ballistic nor Le´vy strategies should be considered as universal (see, e.g., [45,46]), since realistic fluctuations in the targets distribution may induce switches between these two regimes. This observation has been confirmed by recent empirical results [25,27], showing that foragers in the wild do not exhibit movement patterns that can be approximated, at all times, by Le´vy, ballistic or exponential models. Nevertheless, the relevant finding is that in the low-density regime superdiffusive Le´vy strategies remain as the optimal solution in a broad range of heterogeneous landscape conditions, with the optimal exponent
1vmopt*v2 dependent on specific environment properties. Crossovers between superdiffusive and ballistic strategies may also emerge depending on whether strong target spatial fluctuations are local or not, and if they depress or boost the presence of nearby targets. For instance, recent data on a species of jellyfish have reported [27] on Le´vy flight foraging strategies with optimal index as low asmopt~1:18. Moreover, studies on marine predators have also found [24] small values as mopt~1:63. Such rather fast, enhanced superdiffusion (with respect to mopt&2) suggests the occurrence of foraging activity in a highly dynamic and heterogeneous landscape, as it is clearly the case for marine prey landscapes [25,26,67].
In the present work, the question of how the landscape heterogeneity affects the search efficiency in encounter interactions is addressed under conditions of constant global density of scarce resources. In such conditions we predict that efficient strategies with larger diffusion exponents (including superdiffusive ones) should arise, as heterogeneous environments with wider distribu-tions of starting distances between the foraging organism and the nearby targets are considered. Similarly to what occurs in homogeneous landscapes [42], we do not expect density fluctuations in the scarcity regime to modify optimal Le´vy solutionsper se, but only to the extent that fluctuations in density modify the initial searcher-to-targets distances. In other words, Figure 4. Determination of the optimal search strategy of Le´vy
random searchers with Poissonian fluctuations in the distances
to nearby targets, Eq. (4). The condition f(m~mopt,a)~
LSLTp=Lmjm~mopt~0, forl~10
3andr
v~1, provides the optimal Le´vy
exponent, mopt, associated with the strategy of maximum average efficiency. Inset: since strategies with mƒ1 are not allowed (non-normalizable pdf of step lengths), the highest efficiency is always obtained form?1as fluctuations withawacross&312:2are considered, marking the onset of a regime dominated by ballistic optimal search dynamics.
provided that the asymmetry in the searcher-to-targets distances is maintained as density changes, optimal Le´vy strategies should result insensitive to target density fluctuations. This means that for a Le´vy searcher is less important to have advanced knowledge of the density than of the relative positions of the targets. Clearly, robustness to changes in environmental parameters (i.e. density) should be considered as an advantage in non-informed optimal search solutions [42].
If we acknowledge the presence of selective pressures respon-sible for the evolution and maintenance of non-oriented motility in organisms [68], our results lead to a neat empirically testable prediction: patchy and heterogeneous landscapes should promote the emergence of enhanced diffusivity and compromising optimal Le´vy strategies. Even though the empirical inference of large scale movement patterns from heterogeneity properties of the landscape is a difficult task [26], specifically designed and controlled large scale experiments are feasible in the laboratory [68–71] and even in the field [54].
We hope the present study might shed light on unsettled issues related to the efficiency and associated dynamics of organisms performing random searches. Besides the well documented depen-dence of search efficiency on resource density [25,26,34,37,55], our results suggest another relevant aspect of non-universal random search behavior: landscape heterogeneity frames optimal diffusivity.
Acknowledgments
We thank A. S. Ferreira and H. A. Arau´jo for fruitful discussions.
Author Contributions
Conceived and designed the experiments: EPR FB MGEdL GMV. Analyzed the data: EPR FB MGEdL GMV. Contributed reagents/ materials/analysis tools: EPR FB MGEdL PJRN TAS GMV. Wrote the paper: EPR FB MGEdL GMV.
References
1. Viswanathan GM, da Luz MGE, Raposo EP, Stanley HE (2011) The physics of foraging: an introduction to random searches and biological encounters. Cambridge: Cambridge University Press.
2. da Luz MGE, Grosberg A, Raposo EP, Viswanathan GM, eds. (2009) The randomsearch problem: trends and perspectives. J Phys A 42, no. 43. Bristol: IOPscience.
3. Lomholt MA, van den Broek B, Kalisch S-MJ, Wuite GLJ, Metzler R (2009) Facilitated diffusion with DNA coiling. Proc Natl Acad Sci U S A 106: 8204–8208.
4. van den Broek B, Lomholt MA, Kalisch S-MJ, Metzler R, Wuite GLJ (2008) How DNA coiling enhances target localization by proteins. Proc Natl Acad Sci U S A 105: 15738–15742.
5. Mandell AJ, Selz KA, Shlesinger MF (1997) Mode matches and their locations in the hydrophobic free energy sequences of peptide ligands and their receptor eigenfunctions. Proc Natl Acad Sci U S A 94: 13576–13581.
6. Hufnagel L, Brockmann D, Geisel T (2004) Forecast and control of epidemics in a globalized world. Proc Natl Acad Sci U S A 101: 15124–15129.
7. Brockmann D, Hufnagel L, Geisel T (2006) The scaling laws of human travel. Nature 439: 462–465.
8. Shlesinger MF (2006) Random walks: follow the money. Nature Phys 2: 69–70. 9. Gonza´lez MC, Hidalgo CA, Baraba´si A-L (2008) Understanding individual
human mobility patterns. Nature 453: 779–782.
10. Stephens DW, Krebs JR (1987) Foraging theory. Princeton: Princeton University Press.
11. Berg HC (1993) Random walks in biology. Princeton: Princeton University Press.
12. Turchin P (1998) Quantitative analysis of movement: measuring and modelling population redistribution in animal and plants. Sunderland: Sinauer Associates Inc.
13. Shlesinger MF, Klafter J (1986) Le´vy walks versus Le´vy flights. In: Stanley HE, Ostrowsky N, eds. On growth and form. Dordrecht: Nijhoff. pp 279–283. 14. Shlesinger MF, Zaslavsky G, Frisch U, eds. (1995) Le´vy flights and related topics
in physics. Berlin: Springer.
15. Levandowsky M, Klafter J, White BS (1988) Swimming behavior and chemosensory responses in the protistan microzooplankton as a function of the hydrodynamic regime. Bull Marine Sci 43: 758–763.
16. Raposo EP, Buldyrev SV, da Luz MGE, Viswanathan GM, Stanley HE (2009) Le´vy flights and random searches. J Phys A 42: 434003.
17. Viswanathan GM, Raposo EP, da Luz MGE (2008) Le´vy flights and superdiffusion in random search: the biological encounters context. Phys Life Rev 5: 133–162.
18. Petrovskii S, Mashanova A, Jansen VAA (2011) Variation in individual walking behavior creates the impression of a Le´vy flight. Proc Natl Acad Sci U S A 108: 8704–8707.
19. Bartumeus F, Levin S (2008) Fractal reorientation clocks: linking animal behavior to statistical patterns of search. Proc Natl Acad Sci U S A 105: 19072–19077.
20. Bartumeus F, Catalan J (2009) Optimal search behavior and classic foraging theory. J Phys A 42: 434002.
21. Stocker R, Seymour JR, Samadani A, Hunt DE, Polz M (2008) Rapid chemotactic response enables marine bacteria to exploit ephemeral microscale nutrient patches. Proc Natl Acad Sci U S A 105: 4209–4214.
22. Seymour JR, Marcos, Stocker R (2008) Resource patch formation and exploitation throughout the marine microbial food web. Am Nat 173: E15–E29. 23. Oshanin G, Vasilyev O, Krapivsky PL, Klafter J (2009) Survival of an evasive
prey. Proc Natl Acad Sci U S A 109: 13696–13671.
24. Sims DW, Southall EJ, Humphries NE, Hays GC, Bradshaw CJA, et al. (2008) Scaling laws of marine predator search behaviour. Nature 451: 1098–1102. 25. Humphries NE, Queiroz N, Dyer JRM, Pade NG, Musyl MK, et al. (2010)
Environmental context explains Le´vy and Brownian movement patterns of marine predators. Nature 465: 1066–1069.
26. Sims DW, Witt MJ, Richardson AJ, Southall EJ, Metcalfe JD (2006) Encounter success of freeranging marine predator movements across a dynamics prey landscape. Proc R Soc B 273: 1095–1201.
27. Hays GC, Bastian T, Doyle TK, Fossette S, Gleiss AC, et al. (2011) High activity and Le´vy searches: jellyfish can search the water column like fish. E-pub ahead of print. doi: 10.1098/rspb.2011.0978.
28. Zollner PA, Lima SL (1999) Search strategies for landscape-level interpatch movements. Ecology 80: 1019–1030.
29. Bartumeus F, Catalan J, Viswanathan GM, Raposo EP, da Luz MGE (2008) The role of turning angle distributions in animal search strategies. J Theor Bio 252: 43–55.
30. Shlesinger MF (2006) Search research. Nature 443: 281–282. 31. Shlesinger MF (2009) Random searching. J Phys A 42: 434001.
32. Metzler R, Koren T, van den Broek B, Wuite GJL, Lomholt MA (2009) And did he search for you, and could not find you? J Phys A 42: 434005.
33. Viswanathan GM, Afanasyev V, Buldyrev SV, Murphy EJ, Prince PA, et al. (1996) Le´vy flight search patterns of wandering albatrosses. Nature 381: 413–415.
34. Viswanathan GM, Buldyrev SV, Havlin S, da Luz MGE, Raposo EP, et al. (1999) Optimizing the success of random searches. Nature 401: 911–914. 35. da Luz MGE, Buldyrev SV, Havlin S, Raposo EP, Stanley HE, et al. (2001)
Improvements in the statistical approach to random Le´vy flight searches. Physica A 295: 89–92.
36. Edwards AM, Phillips RA, Watkins NW, Freeman MP, Murphy EJ, et al. (2007) Revisiting Le´vy flight search patterns of wandering albatrosses, bumblebees and deer. Nature 449: 1044–1048.
37. Bartumeus F, Peters F, Pueyo S, Marrase´ C, Catalan J (2003) Helical Le´vy walks: adjusting searching statistics to resource availability in microzooplankton. Proc Natl Acad Sci U S A 100: 12771–12775.
38. Bartumeus F (2009) Behavioral intermittence, Le´vy patterns, and randomness in animal movement. Oikos 118: 488–494.
39. Bartumeus F, Fernandez P, da Luz MGE, Catalan J, Sole RV, et al. (2008) Superdiffusion and encounter rates in diluted, low dimensional worlds. Eur Phys J Special Topics 157: 157–166.
40. Be´nichou O, Loverdo C, Moreau M, Voituriez R (2011) Intermittent search strategies. Rev Mod Phys 83: 81–129.
41. Be´nichou O, Coppey M, Moreau M, Suet P-H, Voituriez R (2005) A stochastic model for intermittent search strategies. J Phys Cond Matter 17: S4275–S4286. 42. Lomholt MA, Koren T, Metzler R, Klafter J (2008) Le´vy strategies in intermitent search processes are advantageous. Proc Natl Acad Sci U S A 105: 11055–11059.
43. Reynolds AM (2006) On the intermitent behavior of foraging animals. Europhys Lett 75: 517–520.
44. Reynolds AM, Bartumeus F (2009) Optimising the success of random destructive searches: Le´vy walks can outperform ballistic motions. J Theor Biol 260: 98–103. 45. Plank MJ, James A (2008) Optimal foraging: Le´vy pattern or process? J R Soc
Interface 5: 1077–1086.
46. James A, Planck MJ, Brown R (2008) Optimizing the encounter rate in biological interactions: ballistic versus Le´vy versus Brownian strategies. Phys Rev E 78: 051128.
48. Reynolds AM (2010) Balancing the competing demands of harvesting and safety from predation: Le´vy walk searches outperform composite Brownian walk searches but only when foraging under the risk of predation. Physica A 389: 4740–4746.
49. Lomholt MA, Ambjo¨rnsson T, Metzler R (2005) Optimal target search on a fast-folding polymer chain with volume exchange. Phys Rev Lett 95: 260603. 50. Raposo EP, Buldyrev SV, da Luz MGE, Santos MC, Stanley HE, et al. (2003)
Dynamical robustness of Le´vy search strategies. Phys Rev Lett 91: 240601. 51. Santos MC, Raposo EP, Viswanathan GM, da Luz MGE (2004) Optimal
random searches of revisitable targets: crossover from superdiffusive to ballistic random walks. Europhys Lett 67: 734–740.
52. Koren T, Lomholt MA, Chechkin AV, Klafter J, Metzler R (2007) Leapover lengths and first passage time statistics for Le´vy flights. Phys Rev Lett 99: 160602.
53. Johnson CJ, Parker KL, Heard DC (2001) Foraging across a variable landscape: behavioral decisions made by woodland caribou at multiple spatial scales. Oecologia 127: 590–602.
54. McIntyre NE, Wiens JA (1999) Interactions between landscape structure and animal behavior: the roles of heterogeneously distributed resources and food deprivation on movement patterns. Landsc Ecol 14: 437–447.
55. Bartumeus F, Giuggioli L, Louzao M, Bretagnolle V, Oro D, et al. (2010) Fishery discards impact on seabird movement patterns at regional scales. Curr Bio 20: 1–8.
56. Kardar M (2007) Statistical physics of particles. Cambridge: Cambridge University Press. 37 p.
57. Buldyrev SV, Havlin S, Kazakov AY, da Luz MGE, Raposo EP, et al. (2001) Average time spent by Le´vy flights and walks on an interval with absorbing boundaries. Phys Rev E 64: 041108.
58. Buldyrev SV, Gitterman M, Havlin S, Kazakov AY, da Luz MGE, et al. (2001) Properties of Le´vy flights on an interval with absorbing boundaries. Physica A 302: 148–161.
59. Mantegna RN, Stanley HE (1994) Stochastic process with ultraslow convergence to a Gaussian: the truncated Le´vy flight. Phys Rev Lett 73: 2946–2949.
60. Byers JE (2000) Effects of body size and resource availability on dispersal in a native and a nonnative estuarine snail. J Exp Mar Bio Ecol 248: 133–150. 61. Sutherland WJ, Gill JA, Norris K (2002) Density-dependent dispersal in animals:
concepts, evidence, mechanisms, and consequences. In: Bullock JM, Kenward RE, Hails RS, eds. Dispersal ecology. Oxford: Blackwell Publishing. pp 129–162.
62. South A (1999) Extrapolating from individual movement behaviour to population spacing patterns in a ranging mammal. Ecol Model 117: 343–360. 63. Borger L, Dalziel BD, Fryxell JM (2008) Are there general mechanisms of
animal home range behaviour? A review and prospects for future research. Ecol Lett 11: 637–650.
64. Van Houtan KS, Pimm SL, Halley JM, Bierregaard Jr. RO, Lovejoy TE (2007) Dispersal of Amazonian birds in continuous and fragmented forest. Ecol Lett 10: 219–229.
65. Preston MD, Pitchford JW, Wood AJ (2010) Evolutionary optimality in stochastic search problems. J R Soc Interface 7: 1301–1310.
66. Pasternak Z, Bartumeus F, Grasso FW (2009) Le´vy-taxis: a novel search strategy for finding odor plumes in turbulent flow-dominated environments. J Phys A 42: 434010.
67. Makris NC, Ratilal P, Symonds DT, Jagannathan S, Lee S, et al. (2006) Fish population and behavior revealed by instantaneous continental shelf-scale imaging. Science 311: 660–663.
68. Wei Y, Wang X, Liu J, Nememan I, Singh AH, et al. (2011) The population dynamics of bacteria in physically structured habitats and the adaptive virtue of random motility. Proc Natl Acad Sci U S A 108: 4047–4052.
69. Giuffre C, Hinow P, Vogel R, Ahmed T, Stocker R, et al. (2011) The ciliate Paramecium shows higher motility in non-uniform chemical landscapes. PLoS ONE 6: e15274.
70. Seymour JR, Simo´ R, Ahmed T, Stocker R (2010) Chemoattraction to dimethylsulfoniopropionate throughout the marine microbial food web. Science 329: 342–345.