A wide hybrid zone of chromosome races of the
common shrew,
Sorex araneus
Linnaeus, 1758
(Mamma-lia), between the Dnieper and Berezina Rivers (Belarus)
Yu.M. Borisov, E.V. Cherepanova, V.N. Orlov
Severtsov Institute of Problems of Ecology and Evolution, Russian Academy of Sci-ences, Leninskii pr., 33, Moscow 119071, Russia
E-mails: boris@sevin.ru, e-cherepan@mail.ru, orlovvic@yandex.ru
Abstract. Karyological study of 75 specimens of the common shrew, Sorex araneus
Linnaeus, 1758, from 8 localities in the Berezina River basin (eastern Belarus) was carried out. A wide hybrid zone (not less than 100 km) between the northern West Dvina chromosome race (XX / XYY, af, bc, gm, hk, ip, jl, no, qr, tu) and the southern Turov race (XX / XYY, af, bc, g, h/k, i, jl, m, n, o, p, q, r, tu) was revealed in this region. Frequencies of fused-unfused arms comprising four diagnostic metacentrics of the West Dvina race (g/m, h/k, n/o, q/r) were calculated in all capture sites. Tak-ing into consideration the absence of metacentric ip in specimens from six northern localities, the Borisov (Bs) race (XX / XYY, af, bc, g/m, h/k, i, jl, n/o, p, q/r, tu) (Orlov, Borisov, 2009) was distinguished in these sites. Common shrews from two southern localities on the right and left banks of the Berezina River (Berezino vicin-ity) were referred to the Turov race. The presence of four metacentrics descended from the West Dvina race in the Bs race testifi es to the hypothesis expressed earlier that the polymorphic populations of the S. araneus between the Dnieper and Berezina Rivers originated as a result of the West Dvina race spreading from the north and of hybridization between this race and local populations with acrocentric chromosomes.
Key words: common shrew, Sorex araneus, chromosome race, hybrid zone.
I
NTRODUCTIONThe northern parts of Eastern Europe were covered with ice during the Last Glacial Maximum (24-17 kyr BP) (Markova, van Kolfschoten, 2008). The region covered with the ice sheet was subsequently recolonized by the populations from refugia near the glacier frontier or in the area of the ice sheet (Orlov et al., 2007). The populations of the common shrew that spread after the latest glaciation from refugia formed the recent chromosome races. By defi nition, a chromosome race
of S. araneus is a group of geographically contiguous populations that share the same set of metacentrics and acrocentrics by descent (Hausser et al., 1994).
It is known that the chromosome races characterized by several metacentrics of incomplete monobrachial homology (e.g. hn, ik, mq, and pr of the Seliger race; hi, kr, no,
and pq of the Moscow race; hk, ip, no, and
glaciation (10,000 – 15,000 years) (Orlov et al., 2007; Borisov et al., 2008, 2009).
From the other hand, in contact zones of “acrocentric” chromosome races and “metacentric” chromosome races, which were formed as a result of centric fusions, the penetration of metacentric chromosomes into “acrocentric” populations and vice versa is observed for dozens of kilometers (Searle, Wojcik, 1998; Jones, Searle, 2003; Zima et al., 2003; Fredga, 2007). Moreover, for many
millennia after the latest glaciation some local differentiated populations sometimes arose in such wide transitional zones. Such populations differ by chromosome rearrangements and can be classifi ed as different chromosome races (Fredga, 2007).
In 2007 the karyotypes of common shrews were studied in six localities in the Upper Dnieper River and Berezina River basins. We presented a preliminary description of new chromosome race Borisov (Bs) which
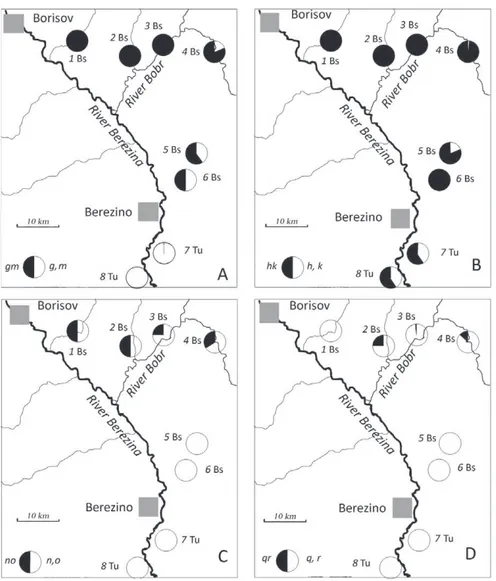
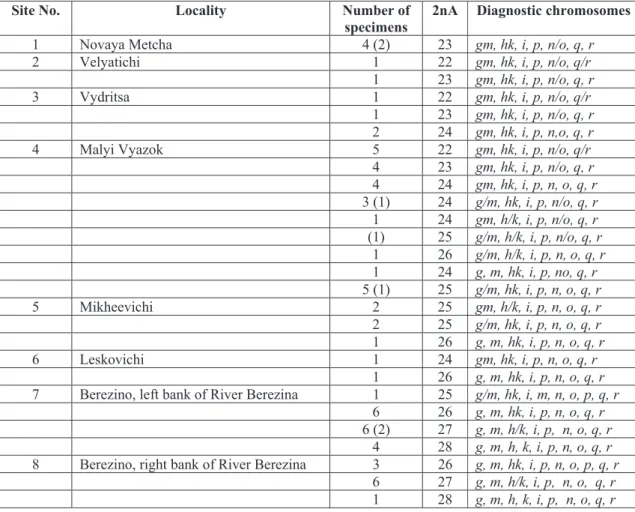
Fig. 1. Frequency of fused (black) and unfused (white) state of chromosome arms: g and m (A), h and k (B),
appeared as a result of hybridization between the monomorphic West Dvina race and local populations with ten pairs of acrocentrics (Orlov, Borisov, 2009).
In 2008 and 2009 we collected additional material in the Berezina River basin and in this transitional zone we discovered popula-tions with various karyotypes. In this paper, the frequency distribution of metacentric and acrocentric variants of the 4 diagnostic chro-mosomes in common shrews from this terri-tory is described.
M
ATERIAL ANDMETHODSCommon shrews were caught with live traps in the years 2008-2009. All the individu-als were karyotyped, and every chromosome arm was identifi ed by G-banding. The trypsin-Giemsa technique of Seabright (1971) was applied. DAPI-banding of chromosomes ac-cording to Minina et al. (2007) was performed for the Turov race. In total, karyotypes of 75 common shrews from 8 populations have been studied. The results of the present study are summarized in the Table, together with the
Table 1. Capture sites and diagnostic chromosomes of the common shrews of races Borisov (sites 1-6) and Turov (sites 7-8) in Belarus (Minsk Prov.). Heterozygosity of a chromosome pair is indicated with a slash (/).
* - Karyotypes reported in paper of Orlov and Borisov (2009) are given in parentheses. Site No. Locality Number of
specimens
2nA Diagnostic chromosomes
1 Novaya Metcha 4 (2) 23 gm, hk, i, p, n/o, q, r
2 Velyatichi 1 22 gm, hk, i, p, n/o, q/r
1 23 gm, hk, i, p, n/o, q, r
3 Vydritsa 1 22 gm, hk, i, p, n/o, q/r
1 23 gm, hk, i, p, n/o, q, r
2 24 gm, hk, i, p, n,o, q, r
4 Malyi Vyazok 5 22 gm, hk, i, p, n/o, q/r
4 23 gm, hk, i, p, n/o, q, r
4 24 gm, hk, i, p, n, o, q, r
3 (1) 24 g/m, hk, i, p, n/o, q, r
1 24 gm, h/k, i, p, n/o, q, r
(1) 25 g/m, h/k, i, p, n/o, q, r
1 26 g/m, h/k, i, p, n, o, q, r
1 24 g, m, hk, i, p, no, q, r
5 (1) 25 g/m, hk, i, p, n, o, q, r
5 Mikheevichi 2 25 gm, h/k, i, p, n, o, q, r
2 25 g/m, hk, i, p, n, o, q, r
1 26 g, m, hk, i, p, n, o, q, r
6 Leskovichi 1 24 gm, hk, i, p, n, o, q, r
1 26 g, m, hk, i, p, n, o, q, r
7 Berezino, left bank of River Berezina 1 25 g/m, hk, i, m, n, o, p, q, r
6 26 g, m, hk, i, p, n, o, q, r
6 (2) 27 g, m, h/k, i, p, n, o, q, r
4 28 g, m, h, k, i, p, n, o, q, r
8 Berezino, right bank of River Berezina 3 26 g, m, hk, i, p, n, o, p, q, r
6 27 g, m, h/k, i, p, n, o, q, r
diagnostic fusions g/m, h/k, n/o, and q/r, chromosome polymorphism was revealed at all 8 localities; the polymorphism and the involved chromosome arms varied. For example, at four localities in the north of the region (1-4 in Table) 9 different karyotypes were present in 39 specimens, 2nA= 22-25. At four localities in the south of the region (5-8 in Table) 6 different karyotypes were revealed in 36 specimens, 2nA= 23-28. To visualize the complicated situation we constructed pie charts showing the frequencies of the fused and unfused states of the individual chromosome arms (Fig. 1, A-D). At seven localities on the left bank of the Berezina River the frequency of fused arms g and m decreased from 100 to 2.6 per cent towards the south (Fig. 1, A). The frequency of fusion h/k reached 100 % in six northern sites and was equal to 50% in two sites in the Berezino vicinity (Fig. 1, B). Arms n, o, q, and r were not fused in 4 localities, while metacentric variant of these arms occurred in 4 northern localities (Fig. 1, C-D).
Taking into consideration exclusively acrocentric chromosomes i and p in common shrews from 6 northern localities (1-6 in Table) and in previously reported specimens from the vicinity of Orsha (Orlov, Borisov, 2009) we referred these samples to preliminarily described Borisov race (Orlov, Borisov, 2009). Here is a more precise defi nition of this new race.
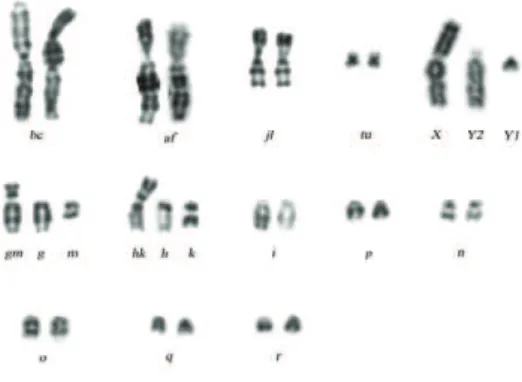
Borisov race (Bs). XX / XYY, af, bc, g/m, h/k, i, jl, n/o, p, q/r, tu (Fig. 2).
Description: Orlov, Borisov, 2009.
Type locality: Borisov Distr., Minsk Prov., Belarus. N 54°14’ E 28°31’
Distribution: East Belarus between the Dnieper and Berezina rivers including the confl uence of the Bobr and Berezina rivers; the distribution area stretches from Orsha in the northeast westward to Borisov and to Berezino in the southeast.
data (7 specimens from 3 sites) from previous-ly published studies (Orlov, Borisov, 2009).
R
ESULTSANDDISCUSSIONThe West Dvina chromosome race was described by Bulatova et al. (2002) in the vicinity of the Zapadnaya Dvina railway station (the Upper West Dvina basin). This race has only been known in the Upper West Dvina basin (Bulatova et al., 2002; Orlov, Borisov, 2007). We were the fi rst in fi nding this chromosome race in the Dnieper River basin (Orlov, Borisov, 2009). The monomorphic karyotype of the West Dvina race, which includes sex chromosomes and nine pairs of metacentric autosomes (XX/XY1Y2, af, bc, gm, hk, ip, jl, no, qr, and tu), is known from the Valdai populations. Five centric fusions serve as diagnostic ones for this race (gm, hk, ip, no, and qr). The common shrews from the Upper Dnieper River also have a monomorphic karyotype. However, the Orsha Hills populations proved to be polymorphic for three fusions (g/m, i/p, and q/r) (Orlov, Borisov, 2009).
Analyzing the specimens from the Berezina River basin we noted the exclusively unfused variant of the i and p arms. As for
Common shrews from two southern localities (Berezino vicinity) with 9 pairs of acrocentric chromosomes should be referred to the Turov race (Mishta et al., 2000) (XX / XYY, af, bc, g, h/k, i, jl, m, n, o, p, q, r, tu) described from Turov Distr. in southern Belarus; the Turov race extends from the Pripyat River (the left bank) to the right bank of River Berezina.
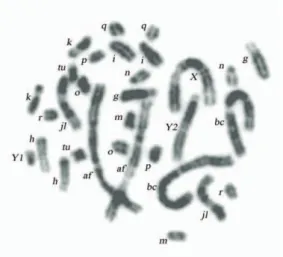
The G-banded karyotype of the new Borisov race is shown in Fig 2, and the G-banded karyotype and DAPI-banded chromosomes (inverted image) of the Turov race – in Figs 3-4.
The frequency of metacentrics descended from the West Dvina race distinctly decreases over a distance of 150 km from the northeast (Kardymovo, Smolensk region) to the southwest (Berezino), which testifi es to the hybrid character of this polymorphism (Orlov, Borisov, 2009; this paper). Apparently, the West Dvina race spread from the north into the area occupied by local populations with acrocentric chromosomes and thus formed a hybrid zone traced in territory 100 km length in southwest direction from Orsha to Berezino.
Common shrew populations with ten pairs of acrocentric chromosomes are known in the south of Europe; for example, the Cordon
chromosome race in the Western Alps and the Pelister race in Macedonia (Zima et al., 1996). As judged by the current distribution of acrocentric chromosomes in common shrew populations in Belarus (Mishta et al., 2000; Orlov, Borisov, 2009), a local chromosome race with acrocentric karyotype was distributed more widely in Glacial epoch than now. Five animals with 10 acrocentric pairs were found to the south of Berezino (this paper). Besides, a specimen with 9 acrocentric pairs and h/k
fusion was found near Turov and one animal with h/i was found near Bobruisk (Orlov, Borisov, 2009). So we suppose that common shrews with acrocentrics may be revealed in the southeast part of Belarus between rivers Pripyat and Dnieper.
During the coldest epoch of the Late Pleistocene the territory of present-day Belarus was under relatively favorable climatic conditions. Periglacial tundra-forest-steppes were predominant there (Markova, van Kolfschoten, 2008). Therefore hybridization of the West Dvina chromosome race and local
Fig. 3. G-banded karyotype of the Turov race, male, g, h, i, k, m, n, o, p, q, r.
“acrocentric” race could have started in the Late Pleistocene.
The most striking peculiarity of the common shrew populations in the Berezina basin is an unequal introgression of 4 metacentrics of the West Dvina race into populations of the Borisov race. So, meta-centric ip was not revealed in all samples studied (this paper) and in Orsha vicinity (Orlov, Borisov, 2009); the absence of metacentric
ip is a diagnostic feature in defi nition of the Borisov race. As for metacentric hk, it was found in all studied samples including representatives of the Borisov and the Turov races. Metacentrics no and qr were not found in two southern capture sites of the Borisov race. The hk combination is widespread in eleven chromosome races from the basins of rivers Wisla, Bug, Neman, and West Dvina, including the West Dvina and Lepel races, which representatives were found in Belarus close to the localities studied here (Mishta et al., 2000; Bulatova et al., 2000; Orlov, Borisov, 2009). This metacentric chromosome has probably introgressed into populations with acrocentric chromosomes earlier than other metacentrics. As we have already mentioned above, the Borisov race appeared as a result of hybridization between the West Dvina and the Turov races. However, the introgression of metacentric hk into populations in the southern part of the Berezina basin, including the Turov race, may be associated not only with the West Dvina race. The Turov race is supposed to be a result of hk introgression from the Lepel race into local populations with acrocentric chromosomes (Orlov, Borisov, 2009).
The origin of the polymorphic race Bori-sov through hybridization between the West Dvina and the Turov races shows that all poly-morphic populations discovered in Belarus could appear as a result of introgression of metacentrics from “metacentric” races into
local “acrocentric” populations at the end of glacial epoch. The presence of acrocentric karyotypes with a single fusion h/i (Bobruisk race) close to the studied territory (Berezino) (Orlov, Borisov, 2009) indicates that different “metacentric” races could contribute to chro-mosomal polymorphism of common shrews in the Berezina basin.
A
CKNOWLEDGEMENTSWe are grateful to A.P. Kashtal’ian, A.A. Kovaleva, A.M. Springer, Z.Z. Borisova, S.A. Dorofeev, and V.A. Bakharev for the assistance during the expedition in Belarus. This study was supported by the Russian Foundation for Basic Research (RFBR grant 09-04-00530a and 09-04-01390a), Program of the Presidium of the Russian Academy of Sciences “Biodiversity,” and Federal Target Program “Research and Scientifi c Educational Specialists of Innovative Russia” (2009-1.1-141-063-021).
R
EFERENCESBorisov Yu.M., Kovaleva A.A., Irkhin S.Y., Orlov V.N. 2009. Zones of contact and joint occurrence of three chromosomal races of the common shrew
Sorex araneus L. (Mammalia) in the southern Valdai Hills // Dokl. Akad. Nauk. 428(2): 275-277. (In Russian, with English translation in Dokl. Biol. Sci. 428: 437-439).
Borisov Yu.M., Kozlovsky A.I., Balakirev A.E., Demidova T.B., Irchin S.Yu., Malygin V.M., Okulova N.M., Potapov S.G., Shchipanov A.N., Orlov V.N. 2008. Contact zones between chromosome races of the common shrew Sorex araneus L. (Insectivora, Mammalia) at the margins of the Veps Stage of the Valdai ice sheet // Sibirskii Ekol. Zh. 15(5) : 763-771. (in Russian, with English translation in Contemp. Probl. Ecol. 1: 583-589).
Bulatova N.Sh., Nadzhafova R.S., Krapivko T.P. 2002. Intraspecifi c phylogenetic relationships in
Genet. 38(1): 64-69.
Bulatova N., Searle J.B., Bystrakova N., Nadjafova R., Shchipanov N., Orlov V. 2000. The diversity of chromosome races in Sorex araneus from European Russia // Acta Theriol. 45(l): 33-46.
Fredga K., 2007. Reconstruction of the postglacial colonization of Sorex araneus into northern Scandinavia based on karyotype studies, and the subdivision of the Abisko race into three // Russian J. Theriol. 6(1): 85-96.
Hausser J., Fedyk S., Fredga K., Searle J.B., Volobouev V., Wojcik J., Zima J. 1994. Defi nition and nomenclature of the chromosome races of Sorex araneus // Folia Zool. 43. Suppl. 1: 1-9.
Jones R.M., Searle J.B. 2003. Mapping the course of the Oxford-Hermitage chromosomal hybrid zone in the common shrew Sorex araneus – a GIS approach // Mammalia. 68: 193-200.
Markova A., van Kolfschoten T. (eds.) 2008. Evolution of the European Ecosystems during Pleistocene- Holocene Transition (24-8 kyr BP). Moscow. 556 p. (In Russian).
Minina J.I., Borodin P.M., Searle J.B., Volobouev V.T. Zhdanova N.S. 2007. Standard DAPI karyotype of the common shrew Sorex araneus L. (Soricidae, Eulipotyphla) // Russian J. Theriol. 6(1): 3-6.
Mishta A.V., Searle J.B., Wojcik J.M. 2000.
Karyotypic variation of the common shrew Sorex araneus in Belarus, Estonia, Latvia, Lithuania and Ukraine // Acta Theriol. 45. Suppl. 1: 47-58.
Orlov V.N., Borisov Yu.M. 2007. Chromosome races of the common shrew Sorex araneus L. (Mammalia: Insectivora) from the south part of Valdai Heights //
Comp. Cytogenet.1(2): 101-106.
Orlov V.N., Borisov Yu.M. 2009. Phylogenetic relationships of the common shrew (Sorex araneus
L., Insectivora) populations in Belarus according to chromosome evidence // Zool. Zh. 88(12): 1506-1514. (In Russian).
Orlov V.N., Kozlovskii A.I., Balakirev A.E., Borisov Yu.M. 2007. Endemic chromosome races of the common shrew Sorex araneus L. (Insectivora, Mammalia) and the possible preservation of refugia in the Late Valdai glaciation area // Dokl. Acad. Nauk.
416(6): 1-4. (In Russian, with English translation in
Dokl. Biol. Sci. 416: 396-399).
Seabright M. 1971. A rapid banding technique for human chromosomes // Lancet. 11: 971-972.
Searle J.B., Wójcik J.M., 1998. Chromosomal evolution: the case of Sorex araneus, (pp. 219-268) // Eds J.M. Wójcik, M. Wolsan. Evolution of shrews. Bialowieza, Poland. 458 p.
Zima J., Fedyk S., Fredga K., Hausser J., Mishta A., Searle J. B.,Volobouev V.T., Wojcik J. M. 1996.
The list of the chromosome races of the common shrew (Sorex araneus) // Hereditas. 125: 97-107.
Zima J., Slivkova L., Tomaskova L. 2003. New data on karyotypic variation in the common shrew, Sorex araneus, from the Czech Republic: an extension of the range of the Laska race // Mammalia. 68: 209-215.
Received May 28, 2010
Accepted by V.G. Kuznetsova, October 31, 2010.