Despite the importance of this group, the knowledge
Texto
Imagem
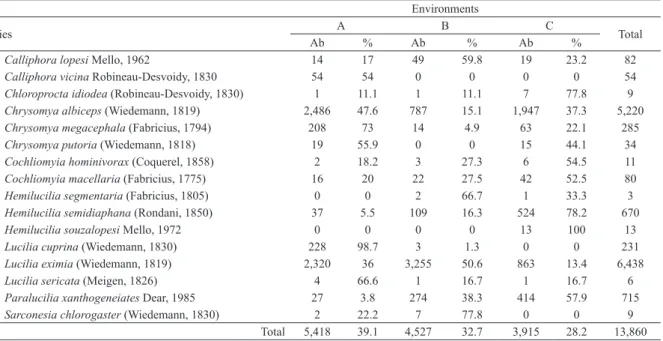
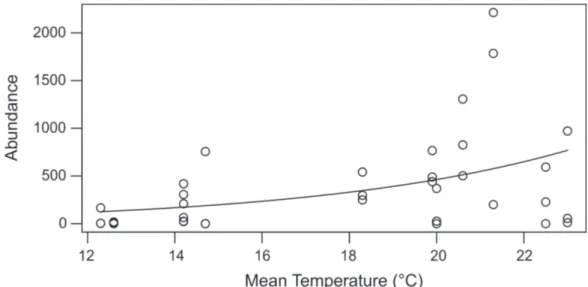
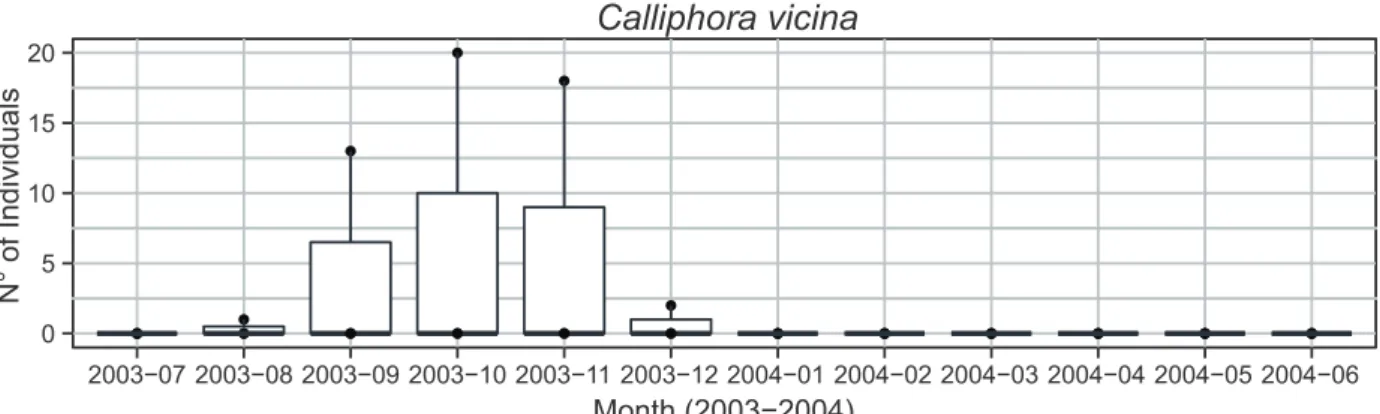
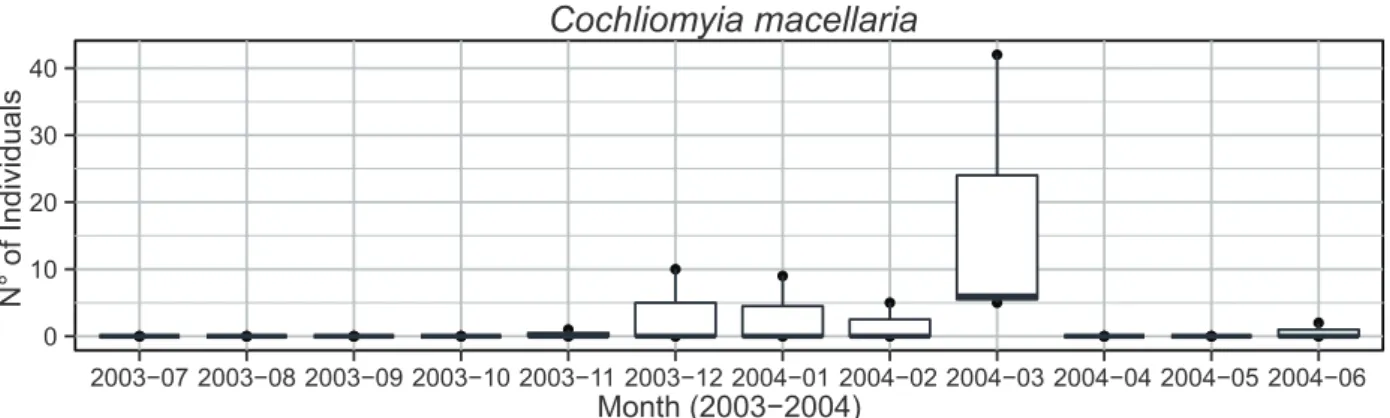

Documentos relacionados
i) A condutividade da matriz vítrea diminui com o aumento do tempo de tratamento térmico (Fig.. 241 pequena quantidade de cristais existentes na amostra já provoca um efeito
didático e resolva as listas de exercícios (disponíveis no Classroom) referentes às obras de Carlos Drummond de Andrade, João Guimarães Rosa, Machado de Assis,
Alguns ensaios desse tipo de modelos têm sido tentados, tendo conduzido lentamente à compreensão das alterações mentais (ou psicológicas) experienciadas pelos doentes
Na hepatite B, as enzimas hepáticas têm valores menores tanto para quem toma quanto para os que não tomam café comparados ao vírus C, porém os dados foram estatisticamente
O serviço de repositório é de fundamental importância, pois, além de prover facilidades para armazenamento e recuperação de artefatos aos demais serviços e ferramentas,
The fourth generation of sinkholes is connected with the older Đulin ponor-Medvedica cave system and collects the water which appears deeper in the cave as permanent
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
In an earlier work 关16兴, restricted to the zero surface tension limit, and which completely ignored the effects of viscous and magnetic stresses, we have found theoretical
