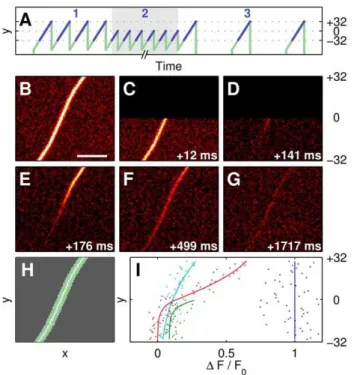
Direct measurement of diffusion in olfactory cilia using a modified FRAP approach.
Texto
Imagem

Documentos relacionados
Por fim, no quarto capítulo apresentamos a caracterização da população em estudo, a sua formação; o professor em ação em seu planejamento com o conteúdo, suas atividades
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
didático e resolva as listas de exercícios (disponíveis no Classroom) referentes às obras de Carlos Drummond de Andrade, João Guimarães Rosa, Machado de Assis,
[r]
Ao Dr Oliver Duenisch pelos contatos feitos e orientação de língua estrangeira Ao Dr Agenor Maccari pela ajuda na viabilização da área do experimento de campo Ao Dr Rudi Arno
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
Managers involved residents in the process of creating the new image of the city of Porto: It is clear that the participation of a resident designer in Porto gave a
(1984) analisando povoamentos de Eucalyptus saligna, aos 8 e 9 anos de idade, respectivamente, verificaram que em média 85% da biomassa aérea encontrava- se no fuste (madeira
