ContentslistsavailableatScienceDirect
Carbohydrate
Polymers
jo u r n al h om ep a g e :w w w . e l s e v i e r . c o m / l o c a t e / c a r b p o l
Sulfated
polysaccharide
from
the
marine
algae
Hypnea
musciformis
inhibits
TNBS-induced
intestinal
damage
in
rats
Tarcisio
V.
Brito
a,
Francisco
C.N.
Barros
b,e,
Renan
O.
Silva
c,
Genilson
J.
Dias
Júnior
a,
José
Simião
C.
Júnior
a,
Álvaro
X.
Franco
c,
Pedro
M.G.
Soares
c,d,
Luciano
S.
Chaves
b,
Clara
M.W.S.
Abreu
f,
Regina
C.M.
de
Paula
f,
Marcellus
H.L.P.
Souza
c,
Ana
Lúcia
P.
Freitas
b,
André
Luiz
R.
Barbosa
a,∗aLAFFEX—LaboratoryofExperimentalPhysiopharmacology,BiotechnologyandBiodiversityCenterResearch(BIOTEC),FederalUniversityofPiauí,
Parnaíba,64202-020PI,Brazil
bLaboratoryofProteinsandCarbohydratesofMarineAlgae,DepartmentofBiochemistryandMolecularBiology,FederalUniversityofCeará,Fortaleza,
60455-760CE,Brazil
cLEFFAG—LaboratoryofPhysiopharmacologyStudyofGastrointestinalTract,FederalUniversityofCeará,Fortaleza,CE,Brazil
dDepartmentofMorphology,MedicalSchool,FederalUniversityofCeara,RuaDelmirodeFariass/n,RodolfoTeofilo,Fortaleza,CECEP60416-030,Brazil eFederalInstituteofEducation,ScienceandTechnologyofCeará,JuazeirodoNorte,CE,Brazil
fLaboratoryofPolymers,DepartmentofOrganicandInorganicChemistry,FederalUniversityofCeará,Fortaleza,60455-760CE,Brazil
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received15February2016
Receivedinrevisedform21April2016 Accepted11June2016
Availableonline14June2016
Keywords:
Sulfatedpolysaccharide Marinealgae Colitis
Inflammatorydisease
a
b
s
t
r
a
c
t
Sulfatedpolysaccharidesextractedfromseaweedhaveimportantpharmacologicalproperties.Thus,the aimofthisstudywastocharacterizethesulfatedpolysaccharide(PLS)fromthealgaeHypneamusciformis
andevaluateitsprotectiveeffectincolitisinducedbytrinitrobenzenesulfonicacidinrats.Thesulfated polysaccharidepossessahighmolecularmass(1.24×105gmol−1)andiscomposedofa-carrageenan,as
depictedbyFT-IRandNMRspectroscopicdata.PLSwasadministeredorally(10,30,and60mg/kg,p.o.)for threedays,startingbeforeTNBS(trinitrobenzenesulfonicacid)instillation(day1).Theratswerekilledon daythree,theportionofdistalcolon(5cm)wasexcisedandevaluatedmacroscopicscoresandwetweight. Then,samplesoftheintestinalwereusedforhistologicalevaluationandquantificationofglutathione, malonyldialdehydeacid,myeloperoxidase,nitrate/nitriteandcytokines.OurresultsdemonstratethatPLS reducedthecolitisandallanalyzedbiochemicalparameters.Thus,weconcludedthatthePLSextracted fromthemarinealgaeH.musciformisreducedthecolitisinanimalmodelandmayhaveanimportant promisingapplicationintheinflammatoryboweldiseases.
©2016ElsevierLtd.Allrightsreserved.
1. Introduction
Marinealgaeareknownforproducinglargeamountsofsulfated polysaccharides,asacomponentoftheircellwalls,widelyusedin thefoodandpharmaceuticalindustrybecauseoftheir rheologi-calpropertiesasgellingandthickeningagents(Barrosetal.,2013; Kravchenkoetal.,2014).Thesepolymerspresentinredalgaeare galactansconsistingofgalactoseormodifiedgalactoseunits clas-sifiedasagaransandcarrageenansbasedonthestereochemistry ofthe4-linkedresidue,whichbelongstotheseries“L”inagarans
∗Correspondingauthorat:BIOTEC/LAFFEX/UFPI,Av.SãoSebastião,n◦2819,
Par-naíba,CEP64202-020PI,Brazil.
E-mailaddress:[email protected](A.L.R.Barbosa).
and“D”incarrageenans(Cosenza,Navarro,Fissore,Rojas,&Stortz, 2014;Manivasagan&Oh,2016;Usov,1998).
Carrageenansareafamilyofsulfatedgalactansformedbya lin-earchainofalternatingresiduesof3-linked-d-galactopyranose
(G-units)and4-linked␣-d-galactopyranose(D-units)or4-linked
3,6-anhydro-␣-d-galactopyranose(DA-units)(Cosenzaetal.,2014;
VandeVelde,Knutsen,Usov,Rollema,&Cerezo,2002).The let-ter codes usedherebyare basedonnomenclatureproposedby
Knutsen, Myslabodski,Larsen,and Usov(1994).The most com-monly used carrageenansare normallyclassified as , and
formsaccordingtotheirsulfationpatternsandtheexistenceof 3,6-anhydro-␣-d-galactopyranose(3,6-AG,symbolizedasDA)(Liang,
Mao,Peng,&Tang,2014;Recaldeetal.,2016).
Marinealgaehave alsoproventoberichsources of biologi-callyactivemolecules,includingtheirpolysaccharides(Ioannou& Roussis,2009;Wang,Ooi,&Ang,2008).Inthiscontext,H.
T.V.Britoetal./CarbohydratePolymers151(2016)957–964
formisisaredalgaethatproducesseveralsulfatedpolysaccharides (PLS)withbiomedicalpotential.Recently,ourresearchgrouphas shownthatthePLSextractedfromHypneamusciformishas mod-ulated theacute inflammatory response mediated primordially byneutrophilmigration (Britoet al., 2013), hascaused protec-tionofgastricmucosaagainstaggressivestimuli,suchasethanol (Damascenoetal.,2013)hasproducedanti-diarrhealactivityin acute,inflammatory,andsecretorydiarrheamodels(Sousaetal., 2016), but untilnowanystudyhasdemonstratedtheaction of thePLSfromHypneaagainstintestinalinjuryintheTNBS-induced colitis.
Inflammatoryboweldiseases(IBD) arechronicinflammatory diseases characterized by inadequate activation of the intesti-nalandsystemicimmunesystemthatcausesadysregulationof mucosalimmunology (decreaseof immune response) and pro-gressiveinflammationonthecolonandgastrointestinaltract.In thiscontext,thetwomajorclinicallydefinedformsof inflamma-toryboweldiseasearetheulcerativecolitisandCrohn’sdisease (Hollander, 1988; Kaser, Zeissig, & Blumberg, 2010; Weber & Turner,2007).
Currently available treatments for IBD are effective only in ameliorating the disease symptoms while having many con-comitantdisadvantages.Antibiotics,one ofthecommonly used therapies,couldadverselychangetheenvironmentalconditionsof microfloraandtriggerresistance.Moreover,immunosuppressant andanti-inflammatorydrugs(suchascorticosteroids)havemany undesirablesideeffects(Sartor,2004).
KnowingthatPLSextractedfromHypneamusciformishave a potentialpharmacologicaleffectsagainstinflammatoryconditions (systemicinflammationanddiarrhea)andgastricdamage,theaim ofthepresentstudywastocharacterizethechemical structure ofthesulfatedpolysaccharideextractedfromHypneamusciformis
andinvestigatewhetherthisgalactancanmodulatetheintestinal damageduringTNBS-inducedexperimentalcolitisinrats.
2. Experimental
2.1. Algaesamples
ThemarineredalgaeH.musciformiswerecollectedfromthe AtlanticcoastatNortheastofBrazil(Flecheirasbeach,Trairí,Ceará, Brazil),atgeographicallocalization:03◦13’25”Sand39◦16’65”
W.AspecimenwasdepositedontheFicologicalHerbariumofthe SeaScienceInstituteatFederalUniversityofCeará–Fortaleza-CE, Brazil(ExsicateN◦2165).Thesampleswerecleanedofepiphytes,
washedwithdistilledwater,andstoredat−20◦C.
2.2. Extractionofsulfatedpolysaccharides
Thesulfatedpolysaccharides(PLS)wereextractedaspreviously described(Farias,Valente,Pereira,&Mourão,2000)withminor changes.Driedtissue(5g)wereflouredandsuspendedin250mLof 0.1Msodiumacetatebuffer(pH5.0)containing5mMEDTA,5mM cysteinewithsubsequentadditionof510mgofpapain(E.Merck). Themixturewasincubatedat60◦Cfor6h.Residuewasremoved
byfiltrationandcentrifugationat2725gfor30minat25◦C.ThePLS
wereprecipitatedbyadditionof16mLof10%cetylpyridinium chlo-ride(CPC,SigmaChemical)andincubationfor24h.Themixture wasthen centrifugedat 8000g for30minat 4◦C. The
polysac-charides in thepelletwere washed with500mLof 0.05% CPC, centrifugedatthesameconditionsandtheprecipitatesuspended in175mLofaNaCl2M:Ethanolsolution(100:15,v/v).The polysac-charideswereallowedtoprecipitatewith300mLethanolandthe pelletwaswashedtwicewith250mLethanol80%(v/v),oncewith
150mLofabsoluteethanolandthreetimeswithacetone,followed bydryingunderhotairflowat60◦C.
2.3. GeneralmethodsforthechemicalanalysisofPLS
Total sugar content was determined by the sulfuric acid-UV technique (Albalasmeh, Berhe, & Ghezzehei, 2013), using
d-galactoseasstandard.Nitrogenandcarboncontentwere
deter-mined by elemental microanalysis (Perkin Elmer CHN 2400). ProteincontentwascalculatedfromN%usingthecorrectionfactor of6.25,asproposedbyMarks,Buchsbaum,andSwain(1985).
2.4. Molarmassdistribution
To estimate the peak molar mass (Mpk) of PLS a High-Performance Size-Exclusion Chromatography(HPSEC) was con-duced in a Shimadzu system equippedwith a refractive index detector(Model RID-10A)at roomtemperature usingan ultra-hydrogellinearcolumn(7.8×300mm),flowrateof0.5mL/min,
witha0.5%polysaccharidesolutionand0.1MNaNO3 assolvent andeluant.Acalibrationcurvewasobtainedbyusingpullulans (ShodexDenko)ofdifferentmolecularweightsrangingfrom103to
106gmol−1.Theequationobtainedfromthiscalibrationplotwas:
LogMw =146,827−106,967Ve (1)
whereVeistheelutionvolumeinmL.Thelinearcorrelation coef-ficientwas0.99.
2.5. Infrared(FT-IR)andnuclearmagneticresonance(NMR) spectroscopy
The Fourier transform infrared spectra (FT-IR) of PLS were recorded witha Shimadzu IRspectrophotometer (model 8300) scanningbetween400and4000cm−1.Thesampleswereanalyzed
asKBrpellets.13Cand1HNMRspectraof2.5%(w/v)solutionsof
PLSinD2Owererecordedat353KonaFouriertransformBruker
AvanceDRX500spectrometerwithaninversemultinuclear gradi-entprobe-headequippedwithz-shieldedgradientcoils,andwith Silicon Graphics. Sodium 2,2-dimethylsilapentane-5-sulphonate (DSS)wasusedastheinternalstandard(0.00ppmfor1H).
2.6. Animals
MaleWistarrats(180–200g)werederivateoftheFederal Uni-versityofPiauí(UFPI).Theanimalswerehousedinplasticcageat 25±2◦Cundera12/12hlight/darkcycle.Allratsweredeprived
offoodfor18–20hpriortotheexperimentalprocedurebutwere allowedfreeaccesstowater.Experimentswereconductedin accor-dance withcurrent established principles for thecare and use ofresearchanimals(NationalInstitutesofHealthguidelines)and approvedbyEthicsCommitteeoftheFederalUniversityofPiauí (ProtocolN◦036/12).
2.7. Inductionofcolitis
Colitiswasinducedbyasingleintracolonicadministrationof 20mgof trinitricbenzenesulfonicacid(TNBS)dissolvedin50% ethanolsolutionintotheeachcolonofratatvolumeof800L.A
T.V.Britoetal./CarbohydratePolymers151(2016)957–964
2.8. Assessmentofcolitis
Theanimalsweredividedinsixgroups(n=5–7rats/group):a negativecontrolgroupthatreceivedintracolonicsaline(GroupI), thecontrolgroupreceivedonlyTNBS(GroupII)intothecolon,the groupsremainingwerepretreatedwithPLS(GroupIII,IVandV:10, 30and60mg/kg,p.o.)ordexamethasone(GroupVI:1mg/kg,s.c.) 1hbeforeofthecolitisinduction.Theanimalsweretreatedeach daywithPLSfractionordexamethasoneinthesecondandthird day.Theratswerekilledonthethirdday,1hafteroftreatment, abdomenswerethen openedandaftertheidentificationofthe intestine,theportionofdistalcolonwasexcised,washedwith0.9% salineandevaluatedthewetweightusingananalyticalbalance, afterthecolonwaspinnedontoawaxblockfortheevaluationof macroscopicscoresbymodifyingthecriteriapreviouslydescribed (Morrisetal.,1989).Then,samplesweretakenforbiochemistry assays.
2.9. Histologicalanalysis
Samplesintestinalwerefixedin10%formalinsolutionfor24h. Then,samplesweretransferredtoasolutionof70%alcohol.The materialwasthenembeddedinparaffinandsectioned;4-m-thick sectionsandstainedwithhematoxylin/eosinandevaluatedbyan experiencedpathologist(P.M.G.S).Histologicalcriteriaevaluated included:mucosalarchitectureloss(0–3),cellularinfiltrate(0–3), musclethickening(0–3),cryptabscess(0,absent;1,present),and gobletcelldepletion(0,absent;1,present).
2.10. Myeloperoxidase(MPO)activity
Briefly,thetissuewashomogenizedin1mLofpotassiumbuffer with0.5%ofhexadecitrimetilamônio(HTAB).Then, homogenate wascentrifuged at 40.000g for 10min at 4◦C. MPOactivity in
the resuspended pellet was assayed by measuring the change inabsorbanceat450nmusingo-dianisidinedihydrochlorideand 1%hydrogenperoxide(Bradley,Priebat,Christensen,&Rothstein, 1982).TheresultswerereportedastheMPOunitspermgoftissue.
2.11. TNF-˛andIL-1ˇlevels
Level of IL-1 and TNF-␣ were evaluated using sandwich Enzyme-Linked Immunoabsorbent Assay (ELISA). ELISA kits for IL-1andTNF-␣werefromtheNationalInstitutefor Biological StandardsandControl(PottersBar,UK).Theresultswereexpressed aspicogramspermilliliter.
2.12. Glutathione(GSH)levels
Samples were homogenized in cold 0.02M EDTA solution (1mL/100mg oftissue).Aliquots(400L)oftissue homogenate weremixedwith320Lofdistilledwaterand80Lof50%(w/v) trichloroaceticacidinglasstubesandcentrifugedat3000rpmfor 15min.Next,400Lofeachsupernatantwasmixedwith800L ofTrisbuffer(0.4M,pH8.9),and20Lof0.01M 5,5-dithio-bis(2-nitrobenzoicacid).Aftershakingthepreparation,absorbancewas measured at 412nm on spectrophotometer (Sedlak & Lindsay, 1968).TheresultsareexpressedasmicrogramsofGSHpergramof tissue.
2.13. Malondialdehyde(MDA)concentration
ToquantifythelevelsofMDAcolonsampleproceededas fol-lows:sampleofthecolonwerehomogenizedwithcold1.15%KCl toprepare10%homogenates,immediatelyafter,250L ofeach homogenatewasaddedto1.5mLof1%H3PO4and0.5mLof0.6%
tert-butylalcohol(aqueoussolution).Then,thismixturewasstirred and heatedin a boilingwaterbath for45min.Thepreparation wasthencooledimmediatelyin anicewaterbath,followedby theadditionof2mLofn-butanol.Thismixturewasshakenand thebutanol layerwasseparated bycentrifugation at1200g for 10min.Absorbancewasdeterminedtobe520and535nm,andthe inferencebetweenthe2determinationswascalculated(Mihara& Uchiyama,1978).MDAconcentrationsareexpressedasmillimoles pergramoftissue.
2.14. NO3−/NO2−levels
Homogenateofintestinaltissueoftheanimalswasincubated ina microplatewithnitratereductase(0.016U/well)for12hto convertNO3−toNO2−.Nitricoxideproductionwasdetermined
bymeasuringnitriteconcentrations in anELISAplatereaderat 540nmusingtheGriessmethod(Greenetal.,1982).Resultswere expressed asmicromoles of nitrite using the internal standard curve.
2.15. Statisticalanalysis
Data were described as either means±SEM or median, as
appropriate.AnalysisofVariance(ANOVA),followedby Student-Newman-Keuls test, was usedto compare means;p<0.05 was definedasstatisticallysignificant.Thehistopathological parame-terswereanalyzedusingtheKruskal–Wallisnonparametrictest, followedbyamultiplecomparisonsDunn’stest.p<0.05was con-sideredstatisticallysignificant.
3. Resultsanddiscussion
3.1. ChemicalanalysisofHypneamusciformispolysaccharide
The Hypnea musciformis polysaccharide (PLS) was obtained by aqueous extraction under digestion by protease, yielding 31.8% of recovery from the algae dry weight. From a solu-tion of 1.0mgmL−1 of PLS (dissolved in distilled water)it was
detected 0.97±0.03mgmL−1 of total soluble carbohydrate by
the UV/sulfuric acid method (Albalasmeh et al., 2013). The homogeneityofthepolysaccharidewasconfirmedbyelemental microanalyses,whichshowedanitrogencontentof0.16±0.03%,
correspondingto1.00±0.18%of proteinresidues.These results
indicatesthattheprocedureusedtoextractedandisolatePLSwas suitabletoyieldacompoundfreeofundesiredmoleculeswhich couldinterfereinthesubsequentexperiments.
The molarmass distributionof Hypnea musciformis polysac-charide(PLS)wasestimatedbyHigh-PerformanceSize-Exclusion Chromatography. The HPSEC chromatogram of PLS (Fig. 1A) showeda singleand narrow peakat 8.97mL, correspondingto a peakmolarmass (Mpk)of 1.24×105gmol−1.High molecular
masses, such as the one described here, are common in car-rageenans (Campo,Kawano,da Silva,&Carvalho, 2009; Pomin, 2010). Cosenza et al. (2014) and Alves et al. (2012) evalu-ated the molar mass of Hypnea musciformis from Natal coast (Brazil).The molarmass obtainedfromAlveset al.(2012) was 1.47×105gmol−1 while Cosenza et al. (2014) obtained molar
massesrangingfrom0.88to2.87×105dependingonextraction
andpurificationmethodologies.Ourresultsareinthesamerange ofthosepreviouslyobtainedforthisalgaespecies.
FT-IRandNMRspectroscopictechniqueswereusedtoanalyze thechemical structure ofPLS. TheIRspectrum ofPLS(Fig.1B) showedbroadbandsat1230and1261cm−1,characteristicof
T.V.Britoetal./CarbohydratePolymers151(2016)957–964
Fig.1.HPSECchromatogramofHypneamusciformispolysaccharideon ultrahydro-gelcolumn(A)andFT-IRspectruminKBrpellets(B).Wavenumbersrangefrom 1400to700cm−1.
1068cm−1(Barrosetal.,2013;Sekkal&Legrand,1993).The
posi-tionofsulfategroupscanbeinferredbybandsat800–850cm−1.
Intense signals at 930cm−1 and 848cm−1 were attributed to
3,6-anhydro-galactopyranose(DA-units) and galactose-4-sulfate (G4S-units),respectively,indicatingthepresenceof-carragenan. Alowintensebandat804cm−1,describedforasulfategroupat
C-2of3,6-anhydro-galactopyranose(DA2S-units),wassuggestive ofasmallamountof-carrageenan, whilenospecificbandsfor -carrageenanwerefound.(Kravchenkoetal.,2014;Villanueva& Montano,2003).
1H and 13C NMR spectra are presented in Fig. 2. 1H NMR
spectrum (Fig. 2A) shows protonfrom anomeric carbon of 3,6 anhydrogalactose(DA)atı5.1andofgalctose-4-sulfate(G4S)at
ı4.63,characteristicof-carrageenan (Kravchenkoetal.,2014; Villanueva&Montano,2003).Alowintensesignalatı5.3isdueto 3,6anhydrogalactose-2sulfate(Cosenzaetal.,2014)indicatingthe presenceofverysmallamountof-carrageenan(7.3%oftotal anhy-drogalactose),incorroborationwithFT-IRresults.Table1shows theattributionofH.musciforms1HNMRsignalsbasedonliterature data(Cosenzaetal.,2014;Kravchenkoetal.,2014;VandeVelde etal.,2002;Villanueva&Montano,2003).TheG4S/DAratiowas calculatedfromH-1protonfromDA(ı5.1)andH-4signalofG4S (ı4.83)duetooverlappingofH-1ofG4SandH-4ofDAsignals. TheratioofG4S/DAwas1.15,veryclosetotheideal-carrageenan
G4S/DAratio.
13C NMR spectrum of purified H.musciforms polysaccharide
(Fig.2B)showsawellresolvedspectrum,indicatingagood
chemi-Fig.2.1H(A)and13CNMR(B)spectraofHypneamusciformispolysaccharideinD2O 2.5%(w/v)recordedat353K.
Table1
1Hand13CNMRchemicalshiftsforHypneamusciformispolysaccharide.
Unita 1Hchemicalshift(ppm)
H-1 H-2 H-3 H-4 H-5 H-6
G4S 4.64 3.60 3.98 4.83 3.80 3.80 DA 5.10 4.13 4.51 4.62 4.62b 4.04b
Unita 13Cchemicalshift(ppm)
C-1 C-2 C-3 C-4 C-5 C-6
G4S 102.60 69.72 78.88 74.23 74.91 61.42 DA 95.28 70.02 79.34 78.43 76.91 70.02
aDiadnomenclatureproposedbyKnutsenetal.(1994). bNotdetected.
calhomogeneity.TheH.musciformsspectrumshowstwoanomeric carbonsintherangeofı95toı105,characteristicofC-1ofG-4S atı102.6and atı95.28due toC-1ofDA.Thisdatacorroborate withourpreviouslydescribedresultsindicatingthatthesulfated polysaccharideextracted from H.musciforms collectedat Ceará state(Brazil)isessentiallya-carrageenan.Theassignmentsof13C
NMRspectrumofH.musciformsaredepictedinTable1,basedon literaturereports (Cosenzaetal.,2014;Kravchenkoetal.,2014; VandeVeldeetal.,2002;Villanueva&Montano,2003).
Thesulfatedpolysaccharidedescribedhereindiffersfromthe onecharacterizedbyAlvesetal.(2012)forthesamealgalspecies collectedindifferentgeographicareasinBrazil,whichpresented a 13C NMR spectrum with four anomeric carbons, depicting a
largefractionof-carrageenanintheirsample.Dependingonthe
fractionationmethodologyused,theauthorsalsofounda larger complexity of polysaccharides, with d- and l-galactans or
T.V.Britoetal./CarbohydratePolymers151(2016)957–964
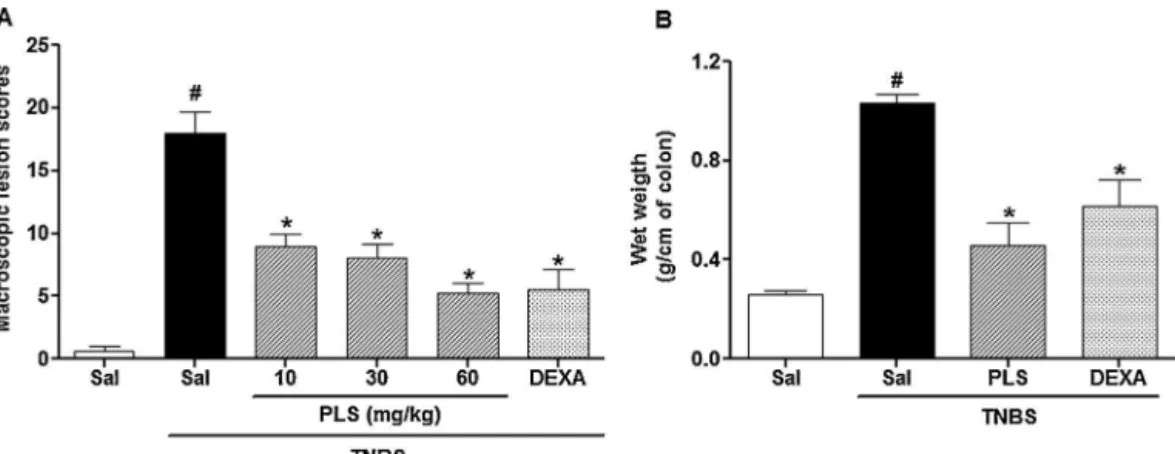
Fig.3. Sulfatedpolysaccharide(PLS)extractedfromHypneamusciformisreducesmacroscopicscores(A)andwetweight(B)ofthecolonsoftheratswithcolitisinducedby TNBS.
Fig.4. Sulfatedpolysaccharide(PLS)extractedfromHypneamusciformisreduceshistologicaldamageoftissueofthecolonsoftheratswithcolitisinducedbyTNBS.Saline (A),TNBS(B),TNBS+PLS(C)andTNBS+DEXA(D).
thechemicalstructureofcarrageenans,suchasalgalsourceand extractionprocedures.Indeed,unlikeproteinsandnucleicacids, carbohydrates are not synthesized through a template-driven mechanism,butratheradirectproductfromthelevelsof expres-sion,activityandsubstrate-specificityofrelatedanabolicenzymes togetherwiththeproperavailabilityofsubstratesforbiosynthesis (Pomin,2016;Varki&Sharon,2008).Thus,asthestructural fea-turesofsulftatedpolysaccharidesvaryasamultifactorialprocess, theirchemicalanalysisbecomesindispensablepriortoa pharma-cologicalormedicinalapproach.
3.2. PolysaccharidefromH.musciformispreventsTNBS-induced intestinaldamage
The search for compounds extracted from natural products withpharmacologicalpropertieshassignificantlycontributedto thediscovery of molecules withimportant biomedical applica-tions(Corrêa, Melo, &Costa, 2008; Sousa et al., 2008).In this context, marinealgae are valuable sources of diversebioactive molecules such as polyphenols, enzymes, and diverse polysac-charides(Karnjanapratum&You,2010).Inthiscontext,thePLS fromHypnea musciformisis an importantpharmacological tool, duetoitsspecificstructurewithhighcontentofradicalsulfates, topresentantioxidantactivity(Alvesetal.,2012), gastroprotec-tiveeffect(Damascenoetal.,2013)andanti-inflammatoryactions (Brito etal.,2013;Sousaet al.,2016).Therefore, inthepresent study,weinvestigatedtheprotectiveeffectofthepolysaccharide (PLS)extractedfromthemarineredalgaeH.musciformisagainst TNBS-inducedintestinaldamageinrats.
TNBS-inducedcolitisisa modelofchronicinflammationand ulcerationintheratcolonthat manifestsmanyof the histolog-icaland clinicalfeaturesofcolonic inflammatorybowel disease inhuman(Morrisetal.,1989),suchasexcessiveoxidativestress, enhancedvascularpermeability,prolongedneutrophilinfiltration andincreasedproductionofinflammatorymediators,accompanied ofasignificantincreaseofthecolonweight.
Inthepresentstudyweshowedclearlythattheintracolonic administrationofTNBS presentedanintensemacroscopic dam-age(18.00±1.7scoresoflesion)accompaniedbymucosalnecrosis
extending along the colon, bowel wall thickening and hyper-emia, showing scores of lesion very high (18.00±1.7 scores),
andincreasedwetweightofthecolon(1.03±0.03g). However,
pretreatment with PLS at the doses of (10, 30 and 60mg/kg p.o.)significantly(p<0.05)reducedinadose-dependentmanner themacroscopicboweldamage(10mg/kg:9.25±2.13;30mg/kg:
5.75±1.84;60mg/kg:5.00±1.87)(Fig.3A)anddecreasedthewet
weightofcolon(60mg/kg:0.45±0.09g)(Fig.3B)afterintracolonic
administrationofTNBS.Dexamethasone(1mg/kg,s.c.),areference drugfor treatmentofthe IBD,alsoreduced macroscopiclesion (5.50±1.65scores)andweightofthecolon(0.61±0.10g).
T.V.Britoetal./CarbohydratePolymers151(2016)957–964
Fig.5. Sulfatedpolysaccharide(PLS)extractedfromHypneamusciformisreduces myeloperoxidaseactivity(A),IL-1(B)andTNF-␣l(C)levelsofthecolonsofthe ratswithcolitisinducedbyTNBS.
3.3. PolysaccharidefromH.musciformisreducesneutrophil infiltrationandcytokinelevelsintheTNBS-inducedcolitis
Toreinforcethis data wemeasuredmyeloperoxidase (MPO) activity,anenzymeproducedmainlybypolymorphnuclear(PMN) leukocytes.MPOactivityisassociatedwithneutrophilinfiltration intissuesandhasbeenwidelyusedtodetectandmonitor intesti-nalinflammation(Halliwell,1996;Yamada,Marshall,Specian,& Grisham,1992).
OurresultsshowthatTNBSadministrationpromotedamarked increaseincolonicMPOlevels(10.03±2.55UMPO/mgoftissue),
ascomparedtosalinegroup(1.22±0.04UMPO/mgoftissue).On
theotherhand,pretreatmentwithPLS(60mg/kg,p.o.)significantly (p<0.05)preventedtheincreaseofcolonicMPOlevels(0.83±0.05
UMPO/mgoftissue)associatedwithTNBSadministration(Fig.5A). Thus,accordingtothesefindings,wecansuggestthatPLSproduced protectiveeffectintheTNBS-inducedcolitismodelbymodulation oftheneutrophilmigrationintotheinflamedcolon.
Theneutrophilmigration intothecolon duringthecolitis is accompanied by production of several inflammatory products, suchasfreeradicalsgenerationandreleasingofpro-inflammatory cytokines(Katz, Itoh, & Fiocchi, 1999). Cytokines as IL-1 and
Table2
EffectofPLSonGSH,MDA,andNO2/NO3levelsinTNBS-inducedcolitis.
Experimentalgroup GSH(g/gtissue) MDA(nmol/gtissue) NO2/NO3(M)
Saline 240.2±18.5 65.0±17.9 0.08±0.01
TNBS 91.5±14.9# 162.5±25.6# 0.22±0.03# PLS+TNBS 225.4±32.4* 76.2±16.3* 0.10±0.01*
Resultsareexpressedasthemeans±S.E.M.of5–6ratspergroup. #P<0.05,whencomparedwithsalinegroup.
*P<0.05,whencomparedwithTNBSgroup.
TNF-␣areincreasedininflamedtissueandresponsiblefor recruit-mentofneutrophilsandmononuclearcellsmediatedinpartbythe up-regulationofadhesionmoleculestowardsvascularendothelial cells.
Fig. 5 shown that TNBS instillation significantly (p<0.05) increased the IL-1 (Fig. 5B; 478.7±139.8pg/mL) and TNF-␣ (Fig. 5C; 99.29±18.42pg/mL) levels, when compared
to saline group (IL-1: 125.8±12.44pg/mL and TNF-␣: 41.23±8.595pg/mL, respectively). In addition, the treatment
with PLS (60mg/kg, p.o.) significantly reduced the concentra-tionsof thesecytokines (IL-1: 108.8±31.03pg/mL and TNF-␣
27.20±2.779pg/mL, respectively) in the inflamed intestinal
mucosa.
3.4. PolysaccharidefromH.musciformisreducesoxidativestress intheTNBS-inducedcolitis
Duringthedevelopmentofcolitis,freeradicalgenerationhas beenproposedasplayinganimportantearlyroleininstallationof thispathogenesis(Pavlicketal.,2002).Thisphenomenonhasan importantrelationshiptotheinitialneutrophilinfiltrationinthe inflamedcolonicmucosa.Therecruitmentandactivationofthese cellsbyreleasingofcytokinesresultinincreaseofthefree radi-calproductionthatoverwhelmsthetissue’santioxidantprotective mechanisms, resulting in a situation of oxidative stress, which definitivelyperpetuatescolonicinflammation(Grisham,1994).
To clarify this point in our experimental model, we inves-tigated two oxidative stress markers, malondialdehyde (MDA) concentrationandglutathione(GSH)levels.MDAisaproductof lipoperoxidativeprocessesthattakeplaceasaconsequenceofthe colonicoxidativeinsult(Loguercioetal.,1996).Studieshaveshown thatcolitiscanincreaseMDAlevelsinratsand,conversely,that variousagentsusedinthetreatmentofthediseasecandecrease thisparameter(Cetinkayaetal.,2006;Girisetal.,2007).Onthe otherhand,GSH;an endogenousantioxidant, protectsthecells againstoxidativedamage,keepingthesulfhydrylgroups(-SH)of proteinsreduced and preventing them fromreacting withfree radicals(Amirshahrokhi,Bohlooli,&Chinifroush,2011). Concen-trationsofendogenous antioxidantssuchas GSHare decreased significantlyinpatientswithinflammatoryboweldiseaseandin experimentalmodelsofcolitis(Spitz,Azzan,&Gius,2004;Tahan etal.,2011).
In this point, our results showed that TNBS promoted a significant (p<0.05) consumption of GSH levels (Table 2: 91.56±14.96mg/g of tissue) and produced an increase in
the MDA concentration (Table 2: 162.5±25.63nmol/g of
tis-sue) in the colonic mucosa, when compared to saline group (240.2±18.55mg/g oftissue and 65.07±17.93nmol/gof tissue,
respectively).However,pretreatmentwithPLSrestoredtheGSH levels(225.4±32.47mg/goftissue)anddecreasedMDA
concentra-tions(76.27±16.30nmol/goftissue)ininflamedtissues(Table2).
T.V.Britoetal./CarbohydratePolymers151(2016)957–964
wecaninferthatPLSdecreasedthemucosadamageactinginthe productionandactionofendogenousantioxidants.
Inaddition,inthepresent studywe showedthat damagein intestinalmucosawasaccompaniedwithhighlevelsofradicals derivedfromnitricoxide(NO).NO3−/NO2−isafreeradicalfrom
nitricoxidewithmoderatereactivitybutwithitsoverproductionby upregulationofinduciblenitricoxidesynthase(iNOS)caninhibit importantenzymesinthemitochondrialresponsibleforcell respi-ration,electrontransportandcausedamageintissuedirectlybythe peroxynitriteformationafterreactionwithsuperoxide(Kaulersch, Fiocchi,&Waldmann,1988;Kolios,Valatas,&Ward,2004).
The Table 2 shows that TNBS group increased level of NO3−/NO2− (0.22±0.03M) in colon tissue, when compared to saline group (0.08±0.01M). However, treatment with PLS significantly (p<0.05) reduces NO3−/NO2− concentration
(0.10±0.00M) in damage intestinal tissue caused by TNBS administration.
4. Conclusions
Inconclusion,ourresultsrevealedthatPLSextractedfromthe marine algae H. musciformis is essentially composedof a high molecularmass-carrageenan.Inaddition,PLSreducedthecolonic
inflammatoryresponseinducedbyadministrationofTNBSthrough inhibitionofcellmigrationintotheinflamedtissueandreduction ofoxidativeprocess.Apossiblemechanismforthisprotectiveeffect mayinvolvethedownregulationoftheinflammatoryresponseby inhibitingthesynthesisandreleasingofproductsofinflammation, suchaspro-inflammatorycytokines.
Conflictsofinterest
Theauthorsdeclarenoconflictofinterest.
Acknowledgments
Theauthorsgratefullyacknowledgethefinancialsupportfrom NationalCounselofTechnologicalandScientific Development– CNPq(Brazil)andResearchfoundationfromthestateofPiauí– FAPEPI.TheauthorsalsowishtoacknowledgeCENAUREMNfor recordingtheNMRspectra.
References
Albalasmeh,A.A.,Berhe,A.A.,&Ghezzehei,T.A.(2013).Anewmethodforrapid determinationofcarbohydrateandtotalcarbonconcentrationsusingUV spectrophotometry.CarbohydratePolymers,97,253–261.
Alves,M.G.C.F.,Dore,C.M.P.G.,Castro,A.J.G.,Nascimento,M.S.,Cruz,A.K.M., Soriano,M.E.,etal.(2012).Antioxidant:cytotoxicandhemolyticeffectsof sulfatedgalactansfromedibleredalgaHypneamusciformis.JournalofApplied Phycology,24,1217–1227.
Amirshahrokhi,K.,Bohlooli,S.,&Chinifroush,M.M.T.(2011).Theeffectof methylsulfonylmethaneontheexperimentalcolitisintherat.Applied Pharmacology,253,197–202.
Barros,F.C.,daSilva,D.C.,Sombra,V.G.,Maciel,J.S.,Feitosa,J.P.,Freitas,A.L., etal.(2013).Structuralcharacterizationofpolysaccharideobtainedfromred seaweedGracilariacaudata(JAgardh).CarbohydratePolymers,92(1),598–603. Bradley,P.P.,Priebat,D.A.,Christensen,R.D.,&Rothstein,G.(1982).Measurement
ofcutaneousinflammation:estimationofneutrophilcontentwithanenzyme marker.JournalofInvestigativeDermatology,78,206–209.
Brito,T.V.,Prudêncio,R.S.,Sales,A.B.,Júnior,F.C.V.,Candeira,S.J.N.,Franco,Á. X.,etal.(2013).Anti-inflammatoryeffectofasulphatedpolysaccharide fractionextractedfromtheredalgaeHypneamusciformisviathesuppression ofneutrophilmigrationbythenitricoxidesignallingpathway.Journalof PharmacyandPharmacology,65,724–733.
Campo,V.L.,Kawano,D.F.,daSilva,D.B.,Jr.,&Carvalho,I.(2009).Carrageenans: biologicalproperties,chemicalmodificationsandstructuralanalysis—a review.CarbohydratePolymers,77(2),167–180.
Cetinkaya,A.,Bulbuloglu,E.,Kantarceken,B.,Ciralik,H.,Kurutas,E.B.,Buyukbese, M.A.,etal.(2006).Effectsofl-carnitineonoxidant/antioxidantstatusinacetic acid-inducedcolitis.DigestiveDiseasesandSciences,51,488–494.
Corrêa,M.F.P.,Melo,G.O.,&Costa,S.S.(2008).Naturalproductsfromplantorigin potentiallyusefulintheasthmatherapy.RevistaBrasileiradeFarmacognosia, 18,785–797.
Cosenza,V.A.,Navarro,D.A.,Fissore,E.N.,Rojas,A.M.,&Stortz,C.A.(2014). ChemicalandrheologicalcharacterizationofthecarrageenansfromHypnea musciformis(Wulfen)Lamoroux.CarbohydratePolymers,102,780–789. Damasceno,S.R.B.,Rodrigues,J.C.,Silva,R.O.,Nicolau,L.A.D.,Chaves,L.S.,
Freitas,A.L.P.,etal.(2013).RoleoftheNO/KATPpathwayintheprotective effectofasulfated-polysaccharidefractionfromthealgaeHypneamusciformis againstethanol-inducedgastricdamageinmice.RevistaBrasileirade Farmacognosia,23,320–328.
Farias,W.R.L.,Valente,A.P.,Pereira,M.S.,&Mourão,P.A.S.(2000).Structureand anticoagulantactivityofsulfatedgalactans:isolationofauniquesulfated galactanfromtheredalgaeBotryocladiaoccidentalisandcomparisonofits anticoagulantactionwiththatofsulfatedgalactansfrominvertebrates.Journal ofBiologicalChemistry,275,29299–29307.
Giris,M.,Erbil,Y.,Dogru-Abbasoglu,S.,Yanik,B.T.,Alis,H.,Olgac,V.,etal.(2007). Theeffectofhemeoxygenase-1inductionbyglutamineonTNBS-induced colitis.InternationalJournalofColorectalDisease,22,591–599.
Green,L.C.,Wagner,D.A.,Glogowski,J.,Skipper,P.L.,Wishnok,J.S.,& Tannenbaum,S.R.(1982).Analysisofnitrate,nitrite:and[15N]nitratein biologicalfluids.AnalyticalBiochemistry,126,131–138.
Grisham,M.B.(1994).Oxidantsandfreeradicalsininflammatoryboweldisease. Lancet,344,859–861.
Halliwell,B.(1996).Antioxidantsinhumanhealthanddisease.AnnualReviewof Nutrition,16,33–50.
Hollander,D.(1988).Crohn’sdiseasesapermeabilitydisorderofthetight junction?Gut,29,1621–1624.
Ioannou,E.,&Roussis,V.(2009).Plant-derivednaturalproducts.Synthesis,function andapplication:naturalproductsfromseaweeds.pp.51–81.Berlin,Germany: Springer.
Karnjanapratum,S.,&You,S.(2010).Molecularcharacteristicsofsulfated polysaccharidesfromMonostromanitidumandtheirinvitroanticancerand immunomodulatoryactivities.InternationalJournalofBiological
Macromolecules,48,311–318.
Kaser,A.,Zeissig,S.,&Blumberg,R.S.(2010).Inflammatoryboweldisease.Annual ReviewofImmunology,28,573–621.
Katz,J.A.,Itoh,J.,&Fiocchi,C.(1999).Pathogenesisofinflammatoryboweldisease. CurrentOpinioninGastroenterology,15,291–297.
Kaulersch,W.,Fiocchi,C.,&Waldmann,T.A.(1988).Polyclonalnatureofthe intestinallymphocytepopulationsininflammatoryboweldisease:amolecular geneticevaluationoftheimmunoglobulinandTcellantigenreceptors. Gastroenterology,95,364–370.
Kim,S.H.,Choi,D.S.,Athukorala,Y.,Jeon,Y.J.,Senevirathne,M.,&Rha,C.K.(2007). AntioxidantactivityofsulfatedpolysaccharidesisolatedfromSargassum fulvellum.InternationalJournalofFoodSciencesandNutrition,12,65–73. Knutsen,S.H.,Myslabodski,D.E.,Larsen,B.,&Usov,A.(1994).Amodifiedsystem
ofnomenclatureforredalgalgalactans.BotanicaMarina,37(2),163–169. Kolios,G.,Valatas,V.,&Ward,S.G.(2004).Nitricoxideininflammatorybowel
disease:auniversalmessengerinanunsolvedpuzzle.Immunology,113, 427–437.
Kravchenko,A.O.,Anastyuk,S.D.,Isakov,V.V.,Sokolova,E.V.,Glazunov,V.P.,& Yermak,I.M.(2014).Structuralpeculiaritiesofpolysaccharidefromsterile formofFarEasternredalgaAhnfeltiopsisflabelliformis.Carbohydrate Polymers,111,1–9.
Liang,W.,Mao,X.,Peng,X.,&Tang,S.(2014).Effectsofsulfategroupinred seaweedpolysaccharidesonanticoagulantactivityandcytotoxicity. CarbohydratePolymers,101,776–785.
Loguercio,C.,D’Argenio,G.,DelleCave,M.,Cosenza,V.,DellaValle,N.,Mazzacca,G., etal.(1996).Directevidenceofoxidativedamageinacuteandchronicphases ofexperimentalcolitisinrats.DigestiveDiseasesandSciences,41,1204–1211. Manivasagan,P.,&Oh,J.(2016).Marinepolysaccharide-basednanomaterialsasa
novelsourceofnanobiotechnologicalapplications.InternationalJournalof BiologicalMacromolecules,82,315–327.
Marks,D.L.,Buchsbaum,R.,&Swain,T.(1985).Measurementoftotalproteinin plantsamplesinthepresenceoftannins.AnaliticalBiochemistry,147(1), 136–143.
Mihara,M.,&Uchiyama,M.(1978).Determinationofmalonaldehydeprecursorin tissuesbythiobarbituricacidtest.AnalyticalBiochemistry,86,271–278. Morris,G.P.,Beck,P.L.,Herridge,M.S.,Depew,W.T.,Szewczuk,M.R.,&Wallace,J.
L.(1989).Hapten-inducedmodelofchronicinflammationandulcerationin theratcolon.Gastroenterology,96,795–803.
Pavlick,K.P.,Laroux,F.S.,Fuseler,J.,Wolf,R.E.,Gray,L.,Hoffman,J.,etal.(2002). Roleofreactivemetabolitesofoxygenandnitrogenininflammatorybowel disease.FreeRadicalBiology&Medicine,33,311–322.
Pomin,V.H.(2010).Structuralandfunctionalinsightsintosulfatedgalactans:a systematicreview.GlycoconjugateJournal,27,1–12.
Pomin,V.H.(2016).Phylogeny,structure,function:biosynthesisandevolutionof sulfatedgalactose-containingglycans.InternationalJournalofBiological Macromolecules,84,372–379.
Recalde,M.P.,Canelón,D.J.,Compagnone,R.S.,Matulewicz,M.C.,Cerezo,A.S.,& Ciancia,M.(2016).Carrageenanandagaranstructuresfromtheredseaweed Gymnogongrustenuis.CarbohydratePolymers,136,1370–1378.
T.V.Britoetal./CarbohydratePolymers151(2016)957–964
Sedlak,J.,&Lindsay,R.H.(1968).Estimationoftotal,protein-bound:and nonproteinsulfhydrylgroupsintissuewithEllman’sreagent.Analytical Biochemistry,24,1992–2005.
Sekkal,M.,&Legrand,P.(1993).Aspectroscopicinvestigationofthecarrageenans andagarinthe1500–100cm−1spectralrange.SpectrochimicaActaPartA:
MolecularandBiomolecularSpectroscopy,49(2),209–221.
Sousa,F.C.F.,Melo,C.T.V.,Citó,C.O.M.,Félix,F.H.C.,Vasconcelos,S.M.M., Fonteles,M.M.F.,etal.(2008).Medicinalplantsandtheirbioactive constituents:ascientificreviewofbioactivityandpotentialbenefitsinthe anxietydisordersinanimalmodels.RevistaBrasileiradeFarmacognosia,18, 642–654.
Sousa,N.A.,Barros,F.C.N.,Araújo,T.S.,Costa,D.S.,Souza,L.K.M.,Sousa,F.B.M., etal.(2016).TheefficacyofasulphatedpolysaccharidefractionfromHypnea musciformisagainstdiarrheainrodents.InternationalJournalofBiological Macromolecules,86,865–875.
Souza,M.C.R.,Marques,C.T.,Dore,C.M.G.,Silva,F.R.F.,Rocha,H.A.O.,&Leite,E. L.(2007).Antioxidantactivitiesofsulfatedpolysaccharidesfrombrownand redseaweeds.JournalofAppliedPhycology,19,153–160.
Spitz,D.R.,Azzan,E.I.,&Gius,D.(2004).Metabolicoxidation/reductionreactions andcellularresponsestoionizingradiation:aunifyingconceptinstress responsebiology.CancerandMetastasisReviews,23,311–322.
Tahan,G.,Gramignoli,R.,Marongiu,F.,Aktolga,S.,Cetinkaya,A.,Tahan,V.,etal. (2011).Melatoninexpressespowerfulanti-inflammatoryandantioxidant
activitiesresultingincompleteimprovementofacetic-acid-inducedcolitisin rats.DigestiveDiseasesandSciences,56,715–720.
Usov,A.I.(1998).Structuralanalysisofredseaweedgalactansofagarand carrageenangroups.FoodHydrocoloids,12,301–308.
VandeVelde,F.,Knutsen,S.H.,Usov,A.I.,Rollema,H.S.,&Cerezo,A.S.(2002).1H and13ChighresolutionNMRspectroscopyofcarrageenans:applicationin researchandindustry.TrendsinFoodScience&Technology,13,73–92. Varki,A.,&Sharon,N.(2008).Historicalbackgroundandoverview.InA.Varki,R.D.
Cummings,J.D.Esko,H.H.Freeze,P.Stanley,C.R.Bertozzi,G.W.Hart,&M.E. Etzler(Eds.),Essentialsofglycobiology.ColdSpringHarbor,NewYork:Cold SpringHarborLaboratoryPress.
Villanueva,R.D.,&Montano,M.N.E.(2003).Finechemicalstructureof carrageenanfromthecommerciallycultivatedkappaphycusstriatum(sacol variety)(Solieriaceae,Gigartinales,Rhodophyta).JournalofPhycology,39, 513–518.
Wang,H.,Ooi,E.V.,&Ang,P.O.(2008).AntiviralactivitiesofextractsfromHong Kongseaweeds.JournalofZheijang,9,969–976.
Weber,C.R.,&Turner,J.R.(2007).Inflammatoryboweldisease:isitreallyjust anotherbreakinthewall?Gut,56,6–8.