Mechanisms of Invasive Species Success
Robyn A. Zerebecki1, Cascade J. B. Sorte1,2*
1Marine Science Center, Northeastern University, Massachusetts, United States of America,2Bodega Marine Laboratory and Department of Evolution and Ecology, University of California at Davis, Bodega Bay, California, United States of America
Abstract
Invasive species are predicted to be more successful than natives as temperatures increase with climate change. However, few studies have examined the physiological mechanisms that theoretically underlie this differential success. Because correlative evidence suggests that invasiveness is related to the width of a species’ latitudinal range, it has been assumed – but largely untested – that range width predicts breadth of habitat temperatures and physiological thermotolerances. In this study, we use empirical data from a marine community as a case study to address the hypotheses that (1) geographic temperature range attributes are related to temperature tolerance, leading to greater eurythermality in invasive species, and (2) stress protein expression is a subcellular mechanism that could contribute to differences in thermotolerance. We examined three native and six invasive species common in the subtidal epibenthic communities of California, USA. We assessed thermotolerance by exposing individuals to temperatures between 14uC and 31uC and determining the temperature lethal to 50% of individuals (LT50) after a 24 hour exposure. We found a strong positive relationship between
the LT50and both maximum habitat temperatures and the breadth of temperatures experience across the species’ ranges.
In addition, of the species in our study, invasives tended to inhabit broader habitat temperature ranges and higher maximum temperatures. Stress protein expression may contribute to these differences: the more thermotolerant, invasive speciesDiplosoma listerianumexpressed higher levels of a 70-kDa heat-shock protein than the less thermotolerant, native
Distaplia occidentalisfor which levels declined sharply above the LT50. Our data highlight differences between native and
invasive species with respect to organismal and cellular temperature tolerances. Future studies should address, across a broader phylogenetic and ecosystem scope, whether this physiological mechanism has facilitated the current success of invasive species and could lead to greater success of invasives than native species as global warming continues.
Citation:Zerebecki RA, Sorte CJB (2011) Temperature Tolerance and Stress Proteins as Mechanisms of Invasive Species Success. PLoS ONE 6(4): e14806. doi:10.1371/journal.pone.0014806
Editor:Peter Roopnarine, California Academy of Sciences, United States of America
ReceivedJuly 17, 2010;AcceptedJanuary 18, 2011;PublishedApril 26, 2011
Copyright:ß2011 Zerebecki, Sorte. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding:Funding was provided by an American Association of University Women (www.aauw.org) American Fellowship to C.J.B.S., a California Environmental Quality Initiative Fellowship to C.J.B.S. from the California Ocean Protection Council (www.opc.ca.gov), the National Science Foundation (OCE-0549944 to S. Williams and M. Bracken; www.nsf.gov), and the Three Seas Program (www.eastwest.neu.edu). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing Interests:The authors have declared that no competing interests exist.
* E-mail: cjsorte@ucdavis.edu
Introduction
Two of the greatest threats to biodiversity and ecosystem functioning are species invasions and global climate change [1–3]. Both introduced species and climate change have, individually, had wide-ranging impacts on species’ abundances, distributions, and interactions, and they have resulted in local extinctions [1, 4–11]. Furthermore, climate change and invasions may interact, with climate-change conditions favoring – and, thus, facilitating the spread of – non-native species [12–15]. The mechanism underlying this interaction between climate change and biological invasions, however, remains unclear. Here, we address tempera-ture tolerance as a potential contributor to the success of invasive species, and we examine heat-shock protein (Hsp) expression as a physiological mechanism of this differential temperature tolerance. Eurythermality, or the ability to maintain physiological function over a wide range of temperatures, is a trait that is likely to be favored as the earth warms [5,8] and may be enhanced in invasive, relative to native, species [10,12,16–18]. If invasive species have broader and/or greater physiological tolerances than natives
and we related habitat temperatures to empirically-determined temperature tolerances.
In addition to addressing eurythermality as a trait conferring success to invasive species, we examined the expression of heat-shock proteins (Hsps) as a potential subcellular mechanism underlying organismal temperature tolerances [27–29]. Heat-shock proteins (including the 70-kda Hsp that we studied, Hsp70) are molecular chaperones that assist in the refolding of denatured cellular proteins and, by so doing, minimize the cellular cost of protein degradation and replacement [27,30–31]. Hsp70 expres-sion can be both constitutive and stress-induced. The level of expression may, therefore, be related both to the innate ability of the organism to withstand environmental stresses and to the amount of stress-induced protein damage the organism has already experienced [31–34]. Recent studies have shown that cellular concentration of Hsp70 can be directly related to organismal temperature tolerance (e.g. [33–36]). Invasive species could have higher temperature tolerances if they express more Hsp70 constitutively or induce Hsp70 expression more quickly, and our study is one of the first to examine differences in Hsp expression between native and invasive species (see also [37–40]). We addressed the hypothesis that invasives are more euryther-mal than native species – and examined heat-shock protein expression as a potential underlying mechanism of thermotoler-ance – in the marine epibenthic community of Bodega Harbor, California, USA. This community is primarily composed of sessile organisms, including tunicates (sea squirts) and bryozoans, that colonize subtidal substrata such as docks, boat hulls, and natural rocky reefs. There has been a recent increase in biological invasions in epibenthic communities over the past several decades, both globally [41] and at our study site in Bodega Harbor, where local ocean temperature has also been increasing [15,42]. We tested two specific hypotheses for nine common species that account for almost 80% of the occupied space in the Bodega Harbor epibenthic community. First, we examined whether geographic temperature limits are related to temperature toler-ance, indicating greater eurythermality in invasive species, based on results from a lethal temperature tolerance experiment.
Second, we used immunoblotting methods to quantify heat-shock protein expression in the nativeDistaplia occidentalis and invasive
Diplosoma listerianum to assess whether heat-shock protein expres-sion could be a subcellular mechanism underlying differences in temperature tolerance. Within this set of common species in a subtidal community, higher temperature tolerance in invasive species was related to geographic temperature range. Further-more, our results for Distaplia and Diplosoma join a small, but growing, body of literature on the potential for Hsp70 expression to underlie temperature tolerance differences between native and invasive species.
Results
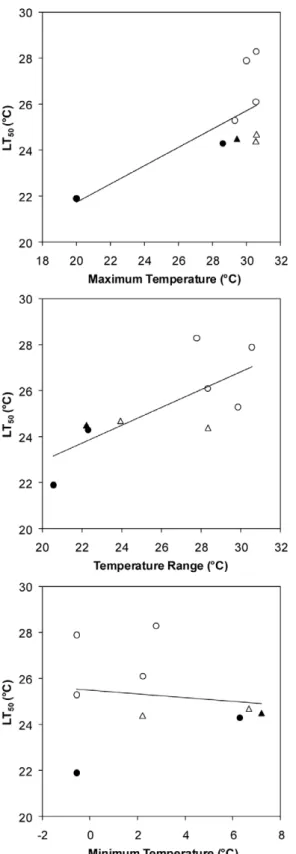
In individuals from this California subtidal epibenthic commu-nity, higher temperature tolerances were related to broader geographic temperature ranges (linear regression,F1,7= 8.54,p=
0.022) and higher maximum habitat temperatures (F1,7= 6.44,
p= 0.039) across these species’ ranges (Figure 2). Minimum habitat temperature was unrelated to this measure of a species’ upper lethal temperature (p= 0.738). Habitat temperature breadth was slightly more strongly related to maximum (p= 0.087) than minimum (p= 0.139) habitat temperature.
The greater average LT50of invasives compared with native
species in our study was driven by differences at 24uC, where mortality was only 14.0% (66.11 SE) for invasives but was 57.8% (621.20) for natives (t-test, t= 2.649, p= 0.033; Figure 3). Mortality curves for individual species are available in Figure S1 in the supporting information. Overall, the invasive species inhabited locations with, on average, broader temperature ranges (28.1uC 60.9) than the natives (21.7uC 60.6; p= 0.003). The invasives also tended to live at higher maximum temperatures (30.3uC60.2 for invasives; 26.0uC63.0 for natives;p= 0.069).
Our analysis of expression of a 70-kda heat-shock protein (Hsp70), a potential subcellular mechanism of organismal temperature tolerance, indicated that Hsp70 levels were higher in the more thermotolerant, invasive tunicateDiplosomathan in the less thermotolerant, native Distaplia (ANOVA, F1,20= 8.065,
p= 0.010; Figure 4). For Distaplia, Hsp70 levels were about twice as high in individuals exposed to temperatures below the species’ LT50 of 21.9uC than above this threshold (t-test, t= 1.855,
p= 0.083; Figure 4). For Diplosoma, we were limited by a low sample size above its LT50 of 27.9uC, which approached our
maximum treatment temperature; however, a decline in Hsp70 expression was not observed at increased temperature for this species.
Discussion
We provide empirical data which indicate that, for species in a subtidal epibenthic community, attributes of geographic temper-ature ranges are related to higher tempertemper-ature tolerances. Among these species, invasives tend to inhabit locations with broader temperature ranges and higher maximum temperatures than native species. Range width has been suggested as a general characteristic of invasion success [16,19,21], attributable either to greater physiological tolerance (e.g., eurythermality), increased propagule pressure (see [43]), or both. Because introduced species have to pass through several abiotic (and biotic) filters in the multi-stage invasion process, species with broader physiological tolerances may be more likely to survive and become established as invaders [44–46]. At the same time, species that are able to occupy larger areas may have more source populations, increasing their likelihood of being transported elsewhere [43,46]. Our results lend some support to this physiological tolerance hypothesis, Figure 1. Conceptual model of greater invasive eurythermality.
Figure 2. Relationship between lethal temperature tolerance and geographic temperature attributes. Each datapoint repre-sents one of the 9 study species, including 3 natives (closed symbols) and 6 invasives (open symbols) within 2 phyla: tunicates (circles) and bryozoans (triangles). Higher LT50values were associated with higher
maximum temperatures (linear regression, p= 0.039) and broader geographic temperature ranges (p= 0.022), calculated by subtracting the minimum monthly mean temperature from the maximum monthly mean temperature across the species’ ranges (see Table 1 and Materials and Methods), but were not related to minimum temperatures. doi:10.1371/journal.pone.0014806.g002
Figure 3. Native and invasive species temperature tolerance. Temperature tolerance of native (closed symbols, solid line;n= 3) and invasive species (open symbols, dashed line;n= 6) shown as mean (61 SE) mortality after a 24 h exposure to six temperature treatments between 14 and 31uC. Logistic curves indicate the higher average LT50
for invasives (dashed line) than natives (solid line) due to the difference in mortality at 24uC (*,p,0.05).
doi:10.1371/journal.pone.0014806.g003
Figure 4. Level of heat-shock protein (Hsp70) in the invasive
Diplosoma(A) and nativeDistaplia(B).Total Hsp70 (shown as mean [61 SE] immunoblot pixel intensity relative to an internal standard) in
without discounting a role for increased propagule pressure, and have implications for the relative impacts of climate change on invasive and native species. To that end, given that (1) broad ranges are characteristic of invaders [20], and (2) we found that temperature tolerance increases with geographic temperature range (across both native and invaded ranges), the possibility that invasive species could be poised for greater success with climate change merits close consideration.
Indeed, a direct comparison of these species in a temperate epibenthic community revealed that invasives are more tolerant of higher temperatures than natives on average (also see [15]). The temperatureversustolerance relationships and differences between natives and invasives were driven by the tunicate species data. For example, as far as tolerance rank, the least tolerant species were natives (Table 1), with the exception of the native bryozoanBugula californica. We suspect that an age effect may partly explain whyB. californica – which had to be hand collected at a more advanced stage due to low recruitment – appeared to be the most thermotolerant native species, when the other two bryozoans (Bugula neritina and Watersipora subtorquata) were the least thermo-tolerant invasives. A positive relationship between age and temperature tolerance (e.g., [47,48]), and between age and survival [49], has been shown for many taxonomic groups, and mostB. californicaindividuals in the LT50experiment were,3–4 weeks older than individuals of the other 8 species. A possible influence of age on B. californica temperature tolerance was indicated by an ad hoc comparison of the tolerance of younger (natural recruitment) and older (dock-collected) individuals. At 20, 24, and 28uC, mortality in replicates composed of younger individuals (followed by sample size) was 50% (1), 50% (2), and 100% (2), respectively, whereas mortality of older individuals –0% (4), 33% (3), and 67% (3) – was lower at all three treatment temperatures. Although this trend was not significant at any temperature (t-test,p.0.2), if age effects led to an increase in LT50
forB. californica, then we would have underestimated the difference in LT50values between native and invasive species. Alternatively,
such departures from the general pattern of higher tolerance in invasives highlight the importance of broadening the phylogenetic scope of physiological tolerance studies of invasion success.
To assess the role of tolerance in invasion success and whether invasive species may be favored in a changing climate, it is also important to consider our temperature tolerance results concur-rent with studies of additional physiological and demographic processes. For example, these results corroborate conclusions from the same system based on differential temperature effects on recruitment [14,50], as well as on growth and survival [15]. Recruitment is positively related to temperature for 5 of the 6 invasives but not for the 2 natives (all butAscidia) considered here [50]. In addition, when mean temperature was increased by 4.5uC, survival was not affected for invasives but was decreased for the one native considered (Distaplia), and temperature impacts on growth rates were, in general, more positive for invasives than natives [15].
In Bodega Harbor, species with the lowest temperature tolerances may be the most susceptible to global warming, whereas abundances of species with the highest temperature tolerances may be most augmented. The common nativeDistaplia
is likely to experience the greatest reduction in abundance as not only does it have the lowest LT50 (Table 1) but, in relation to
invasives, it also experienced higher mortality and lower enhancement of growth when mean temperature was increased by 4.5uC [15]. Conversely, the invasive tunicates Diplosoma and
Botryllus schlosseri had the highest LT50 values, and they
experienced decreases in survival and increases in growth rate at
+4.5uC ([15]; M. Cockrell & C. Sorteunpubl. data). Among this set of species, there was no indication of an inverse relationship between thermotolerance and habitat temperature, as has been documented in several other systems [51–54], either across their geographic ranges or in Bodega Harbor (where they all inhabit the same maximum temperatures). However, a study of geographic variation in temperature tolerance of four of these invasive species (Botrylloides, Botryllus, Diplosoma,andB. neritina) did find evidence of more tolerant species living closer to their tolerance limits, suggesting the need for comparative studies to detect such a relationship at a geographic scale [55].
The expression of heat-shock proteins is a subcellular mechanism that could be partly responsible for differences in organismal responses to increased temperature. In our inter- and
Table 1.Geographic temperature limits and lethal temperatures (LT50) for the 9 study species.
Limits of Geographic Temperature Range
Temp. Limits (6C)
Species Origin Taxon Minimum Maximum Min. Max. LT50
Distaplia occidentalis Native Tunicate Anchorage, AK, USA La Jolla, CA, USA [66] -0.6 20.0 21.9
Ascidia ceratodes Native Tunicate Departure Bay, BC, Canada Panama City, Panama1 6.3 28.6 24.3
Bugula neritina Invasive Bryozoan Boston, MA, USA [69] Key West, FL, USA 2.2 30.6 24.4
Bugula californica Native Bryozoan Cape St. James, BC, Canada Grand Isle, LA, USA 7.2 29.4 24.5
Watersipora subtorquata Invasive Bryozoan Port Townsend, WA, USA [72] Red Sea [67],2 6.7 30.6 24.7
Botrylloides violaceus Invasive Tunicate Anchorage, AK, USA [67] Palau2 -0.6 29.3 25.3
Didemnum vexillum Invasive Tunicate Bar Harbor, ME, USA [67] Key West, FL, USA 2.2 30.6 26.1
Diplosoma listerianum Invasive Tunicate Anchorage, AK, USA Miami Beach, FL, USA -0.6 30.0 27.9
Botryllus schlosseri Invasive Tunicate Eastport, ME, USA Naples, FL, USA 2.8 30.6 28.3
1M. Carman,pers. comm.; temperature data were from the Smithsonian Tropical Research Institute [76]. 2Temperature data were from the World Ocean Atlas [73].
Species are in order of increasing LT50(temperature lethal to 50% of individuals). Geographic limits are from the Ocean Biogeographic Information System [71], and temperature measurements are from NOAA tide gauge [74] and Canadian lighthouse data [75], except where indicated by footnotes. Temperature limits are the minimum and maximum monthly means at these geographic temperature limits (see Materials and Methods).
intra-specific comparisons of expression by the tunicatesDistaplia
and Diplosoma, we found that Hsp70 levels were higher in the invasiveDiplosomathan in the nativeDistaplia, corresponding with the invasive’s relatively greater temperature tolerance. Because individuals of these two species had the same thermal history, this variation between species is not likely due to differential heat-hardening but, rather, may represent either higher constitutive, or more rapidly induced, Hsp expression inDiplosoma. InDistaplia, individuals exposed to temperatures below the species’ LT50had
relatively high Hsp70 levels and low mortality rates; however, in individuals exposed to temperatures above the LT50, Hsp70 levels
were decreased approximately by half and mortality was high. These results are consistent with previous studies that have illustrated a positive relationship between Hsp70 expression and temperature tolerance (see [27,56]). This is among the first sets of studies to address the role of the heat-shock response in explaining differences in physiological tolerances between native and invasive species. Hofmann & Somero [37] and Braby [38] both demonstrated greater induction of Hsp70, as well as a higher Hsp70 induction temperature, in the invasive mussel Mytilus galloprovincialisthan its native congenerMytilus trossulus.In addition, Henkel et al. [40] found a greater increase in hsp70 gene expression in the invasive kelp Undaria pinnatifida than in two native California species. Studies have also revealed alternative mechanisms of native versus invasive physiological tolerances, including enzyme function [57] and heart function [58]. Together, these comparisons suggest that invasives may currently have a greater capacity to deal with thermal stress than native species.
As ocean temperatures continue to increase in Bodega Harbor, as well as worldwide, it is possible that the resident epibenthic species could evolve increased temperature tolerances. However, no study to date has examined the possibility of adaptation to climate change in tunicate or bryozoan species, even though local adaptation, including of temperature tolerance [59], has been observed over distances of ,60 km owing to relatively short dispersal distances of many epibenthic species [59,60]. Local adaptation could help to explain the finding that for 8 out of 9 species, LT50 values for California populations were lower than
the maximum habitat temperatures across the species’ ranges. In fact, populations ofB. neritina, Botrylloides, Botryllus, andDiplosoma
living on the USA east coast had higher tolerance limits than these populations in California [55].
Adaptive potential should be increased in species with shorter generation times and with greater phenotypic – suggesting greater genetic – variation. Generation times are short in these species; for example,Botrylluscan reach sexual maturity in 1 month [61], and populations in Maine, USA sexually reproduce 1–2 times per year [60]. Intraspecific phenotypic variation in temperature tolerance is apparent as differential mortality among individuals in the species mortality plots (Figure S1). All individuals of the 9 species experienced 100% mortality at 31uC. However, some survival in response to a 28uC exposure was observed for all 6 invasives and for 1 of 3 natives (B. californica), and some individuals of all invasive species and 2 of 3 natives survived at 24uC (Figure S1). Conversely,
Distapliashowed no individual variation in mortality across any of the temperature treatments (average mortality at each temperature was either 0% or 100%), suggesting limited variation and, consequently, limited adaptive potential. Thus, even if adaptation occurs in these epibenthic species, the 6 invasives seem to have both an initial advantage over the natives, due to their current higher LT50values, and a potential adaptive advantage because of
their greater range of responses to the temperature exposures. In conclusion, we present data from nine of the most common species in a subtidal community that serve as a starting point for
examining the hypothesis that habitat temperatures across the species’ ranges are associated with temperature tolerance. Our comparison of native and invasive tunicate species indicates the possibility that differences in Hsp70 expression could partly underlie the higher temperature tolerance of invasives. Future studies that include a greater number of species will help to clarify whether greater temperature tolerance and higher Hsp70 expression are general characteristics of invasive speciesper se.
Temperature tolerance differences between natives and inva-sives may be especially important in determining the impacts of extreme high temperature events which are predicted to increase in frequency and severity over the next decade [9,62]. Although daily average temperatures in Bodega Harbor typically ranged from 9–17uC (between the years 2005–9; [50]), well below the LT50of all 9 epibenthic species, a heat wave in July 2006 led to
peak hourly temperatures of almost 20uC, with 19uC sustained over a 24 h period [63]. If, as predicted by climate models, mean ocean temperature increases by 3–4uC [9,64], and heat wave severity increases by ,2uC [62], over the next century, then Bodega Harbor temperatures during extreme events could exceed the LT50 of Distaplia (21.9uC) within the near future and of all
three natives (Table 1) by the end of the 21st century. Further studies that address the extent to which these findings for common species in a marine epibenthic community reflect a general pattern will continue to elucidate whether differences in eurythermality, temperature tolerance, and Hsp expression may predispose invasive species for greater success as global temperatures increase.
Materials and Methods
We focused our study on nine species from the epibenthic community of Bodega Harbor, Bodega Bay, California (38.3290u
N, 123.0581uW), including three natives and six invasive species from two phyla (Table 1). These 9 species accounted for approximately 79% of occupied space (excluding mussels which act, themselves, as substrate) based on a survey we conducted of the adult dock community.
Geographic range and temperature limits
We determined the geographic range of each focus species by consulting primary literature, field guides and an online database (e.g. [65–72], M. Carman pers. comm.). Within each species’ geographic range, we identified the geographic temperature range using data provided by the NOAA National Oceanographic Data Center [73,74], Canadian Department of Fisheries and Oceans [75], and Smithsonian Tropical Research Institute [76]. Minimum temperatures were average monthly temperatures from the coldest months, and maximum temperatures were from the warmest months, across the species’ ranges; neither were necessarily experienced at the latitudinal extremes. We used t-tests to compare the geographic temperature ranges of native and invasive species, and we used linear regression to relate temperature range and limits to temperature tolerance (LT50).
Lethal temperature experiment
recruitment, colonies were collected from the docks by hand and were connected to tiles with wire.
Individuals were acclimated in the running seawater table at approximately 12uC for 24 h prior to the experiment. During the experiment, tiles were suspended in separate containers (1 L). Temperature was raised gradually (1uC per 15 min) to 6 treatment temperatures (approx. 14, 16, 20, 24, 28, and 31uC) using aquarium heaters (7.5W, #T10401, Hydor USA, Sacramento, CA, USA). Each species 6 temperature combination was
replicated 5 times with the exceptions that forB. neritina,n= 7 at 20 and 24uC, andAscidiawere only exposed to 20, 24, and 28uC (n = 3, 3 and 2, respectively) due to low recruitment. Species6
temperature combinations were randomly assigned to tanks and experimental runs (9 runs over a 4 wk period). Water temperatures were monitored using a handheld digital thermometer (HH21, Omega Engineering, Inc., Stamford, CT, USA), and the average treatment temperatures across all replicates were calculated and used in the LT50analyses.
Survival was determined by zooid responsiveness, with individuals scored as ‘live’ or ‘dead’. Tiles were examined immediately after the 24 hr temperature exposure and after 2 days of recovery in ambient running seawater. Because survival values did not vary with recovery time, we used the first observation for all analyses. We calculated species’ LT50 values
using Probit analysis, and t-tests were used to compare survival of native and invasive species at each temperature treatment. All data were analyzed using SAS v.9.1 (SAS Institute Inc., Cary, NC, USA).
Heat-shock protein analysis
Hsp70 expression patterns across treatment temperatures were quantified for two species of colonial tunicates: the nativeDistaplia
and invasiveDiplosoma. Individuals used in the LT50 experiment
were flash frozen in liquid nitrogen after 2 days of recovery in running seawater, and tissue was stored at280uC. We successfully detected Hsp70 in 3 species (Distaplia,Diplosoma,andDidemnum) but not in 3 other species (Botrylloides, Botryllus, and the cryptogenic hydroid Obeliasp.) with the primary antibodies tested (SPA-822 and SPA-805, StressGen, Victoria, BC, Canada).
Samples were prepared by vortexing with glass beads in 30– 50mL of homogenization buffer. The homogenate was heated in a water bath for 5 min at 100uC and centrifuged at 14000 rpm for 15 min. Supernatant protein levels were determined using a BCA protein assay with amounts modified for a NANO-Drop 1000 spectrophotometer (both by Thermo Fisher Scientific Inc.,
Wilmington, DE, USA). Samples (20mg protein) were run on a
10% polyacrylamide gel and transferred to nitrocellulose mem-brane (75 min at 80V). Memmem-branes were stored overnight in blocking solution (0.1% Tween220+5% Non-fat dry milk in TBS [Tris buffer]).
Hsp70 was detected by western blotting. The primary antibody was a mouse monoclonal anti-HSP70 (1:1000, 90 min; SPA-822, StressGen, Victoria, BC, Canada) and secondary antibody was a goat anti-mouse IgG (1:1000, 60 min; SAB-100, StressGen, Victoria, BC, Canada). Hsp70 was visualized using Enhanced Chemiluminescent reagents (ECL-Plus, GE Healthcare, Piscat-away, NJ, USA) and quantified using Adobe Photoshop CS3 v.10.0.1 (Adobe Systems Inc., San Jose, CA, USA). Hsp70 levels reported are relative to a positive control (field-collected
Didemnum), and relative pixel intensity was compared between species and temperature treatments with a 2-way ANOVA (including species, temperature, and their interaction) run in SAS. All data are presented as means61 SE.
Supporting Information
Figure S1 Mean (61 SE) mortality at 6 treatment temperatures between 14 and 31 degrees Celsius for each species (A, B. neritina; B, Watersipora; C, Botrylloides; D, Didemnum; E, Diplosoma; F, Botryllus; G, Distaplia; H, Ascidia; I, B. californica). LT50 after a 24 h exposure is denoted by the dashed lines.
Found at: doi:10.1371/journal.pone.0014806.s001 (0.38 MB TIF)
Acknowledgements
We especially thank L. Miller for generously helping with the Hsp analyses, M. Cockrell for her assistance in the field and ‘Penthouse’ lab, and M. Bracken for providing invaluable suggestions. We thank J. Cerda, E. Jones, A. Newsom, and S. Williams for assistance and feedback. G. Trussell and S. Vollmer provided space and equipment at Northeastern University’s Marine Science Center. Comments from M. Bracken, A.R. Hughes, and L. Miller greatly improved this manuscript. R.A.Z. thanks East West XXV and S. Genovese for guidance and support. This publication is contribution number 277 of the Marine Science Center of Northeastern University and a contribution of the Bodega Marine Laboratory, University of California at Davis.
Author Contributions
Conceived and designed the experiments: RAZ CJBS. Performed the experiments: RAZ CJBS. Analyzed the data: RAZ CJBS. Wrote the paper: RAZ CJBS.
References
1. Vitousek PM, Mooney HA, Lubchenco J, Mellilo JM (1997) Human domination of Earth’s ecosystems. Science 277: 494–499.
2. Sala OE, Chapin III FS, Armesto JJ, Berlow E, Bloomfield J, et al. (2000) Global biodiversity scenarios for the year 2100. Science 287: 1770–1774.
3. Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, et al. (2008) A global map of human impact on marine ecosystems. Science 319: 948– 952.
4. Ruiz GM, Fofonoff P, Hines AH, Grosholz ED (1999) Non-indigenous species as stressors in estuarine and marine communities: Assessing invasion impacts and interactions. Limnol Oceanogr 44: 950–972.
5. Carlton JT (2000) Global change and biological invasions. In: Mooney JT, Hobbs RJ, eds. Invasive species in a changing world. Washington: Island Press. pp 31–54.
6. Grosholz E (2002) Ecological and evolutionary consequences of coastal invasions. Trends Ecol Evol 17: 22–27.
7. Walther GR, Post E, Convey P, Menzel A, Parmesan C, et al. (2002) Ecological response to recent climate change. Nature 416: 389–395.
8. Parmesan C (2006) Ecological and evolutionary responses to recent climate change. Annu Rev Ecol Syst 37: 637–669.
9. Intergovernmental Panel on Climate Change (2007) Climate Change 2007: The Physical Science Basis. Cambridge: Cambridge University Press.
10. Hellman JJ, Byers JE, Bierwagen BG, Dukes JS (2008) Five potential consequences of climate change for invasive species. Conserv Biol 22: 534–543. 11. Dijkstra JA, Harris LG (2009) Maintenance of diversity altered by a shift in dominant species: implications for species coexistence. Mar Ecol Progr Ser 387: 71–80.
12. Dukes JS, Mooney HA (1999) Does global change increase the success of biological invaders? Trends Ecol Evol 14: 135–139.
13. Byers JE (2002) Impact of non-indigenous species on natives enhanced by anthropogenic alteration of selection regimes. OIKOS 97: 449–458. 14. Stachowicz JJ, Terwin JR, Whitlach RB, Osman RW (2002) Linking climate
change and biological invasions: ocean warming facilitates nonindigenous species invasions. Proc Natl Acad Sci USA 99: 15497–15500.
15. Sorte CJB, Williams SL, Zerebecki RA (2010) Ocean warming increases threat of invasive species in a marine fouling community. Ecology 91: 2198–2204. 16. Rejma´nek M (2000) Invasive plants: approaches and predictions. Aust Ecol 25:
497–506.
17. Kolar CS, Lodge DM (2001) Progress in invasion biology: predicting invaders. Trends Ecol Evol 16: 199–204.
19. Forcella F, Wood JT (1984) Colonization potential of alien weeds are relative to their ‘‘native’’ distributions: implications for plant quarantine. J Aust Inst Agric Sci 50: 35–40.
20. Rejma´nek M (1995) What makes a species invasive? In: Pysˇek P, Prach K, Rejma´nek M, Wade M, eds. Plant invasions: general aspects and special problems. Amsterdam: SPB Academic Publishing. pp 3–13.
21. Rejma´nek M (1996) A theory of seed plant invasiveness: The first sketch. Biol Conserv 78: 171–181.
22. Rejma´nek M, Richardson DM (1996) What attributes make some plant species more invasive? Ecology 77: 1655–1661.
23. Carveth CJ, Widmer AM, Bonar SA (2006) Comparison of upper thermal tolerances of native and nonnative fish species in Arizona. Trans Amer Fish Soc 135: 1433–1440.
24. Human KG, Gordon DM (1996) Exploitation and interference competition between invasive Argentine ant, Linepithema humile, and native ant species. Oecologia 105: 405–412.
25. McMahon RF (2002) Evolutionary and physiological adaptations of aquatic invasive animals: r selection versus resistance. Can J Fish Aquat Sci 59: 1235–1244.
26. Holway DA, Suarez AV, Case TJ (2002) Role of abiotic factors in governing susceptibility to invasion: a test with Argentine ants. Ecology 83: 1610–1619. 27. Feder ME, Hofmann GE (1999) Heat-shock proteins, molecular chaperones,
and the stress response. Annu Rev Physiol 61: 243–282.
28. Hochachka PW, Somero GN (2002) Biochemical adaptation: mechanism and process in physiological evolution. Oxford: Oxford University Press. 478 p. 29. Tomanek L (2008) The importance of physiological limits in determining
biogeographical range shifts due to global climate change: the heat shock response. Physiol Biochem Zool 81: 709–717.
30. Lindquist S (1986) The heat-shock response. Annu Rev Biochem 55: 1151–1191.
31. Parsell DA, Lindquist S (1993) The function of heat-shock proteins in stress tolerance: degradation and reactivation of damaged proteins. Annu Rev Genet 27: 437–439.
32. Roberts DA, Hofmann GE, Somero GN (1997) Heat-shock protein expression inMytilus californianus: acclimatization (seasonal and tidal-height comparisons) and acclimation effects. Biol Bull 192: 309–320.
33. Sørensen JG, Kristensen TN, Loeschcke V (2003) The evolutionary and ecological role of heat shock proteins. Ecol Lett 6: 1025–1037.
34. Sorte CJB, Hofmann GE (2005) Thermotolerance and heat-shock protein expression in Northeastern PacificNucellaspecies with different biogeographical ranges. Mar Biol 146: 985–993.
35. Sanders BM, Hope C, Pascoe VM, Martin LS (1991) Characterization of the stress protein response in two species of Collisella limpets with different temperature tolerances. Physiol Zool 64: 1471–1489.
36. Sun W, Montagu MV, Verbruggen N (2002) Small heat shock proteins and stress tolerance in plants. Biochim Biophys Acta Gene Struct Expr 1577: 1–9. 37. Hofmann GE, Somero GN (1996) Interspecific variation in thermal
denatur-ation of proteins in the congeneric musselsMytilus trossulusandM. galloprovincialis: evidence from the heat-shock response and protein ubiquitination. Mar Biol 126: 65–75.
38. Braby CE (2004) Physiological ecology of native and invasive blue mussels (genus
Mytilus) in central California. Ph.D. Thesis. Stanford University, California. 39. Henkel SK, Hofmann GE (2008) Differing patterns ofhsp70gene expression in
invasive and native kelp species: evidence for acclimation-induced variation. J Appl Phycol 20: 915–924.
40. Henkel SK, Kawai H, Hofmann GE (2009) Interspecific and interhabitat variation inhsp70gene expression in native and invasive kelp populations. Mar Ecol Progr Ser 386: 1–13.
41. Lambert G (2007) Invasive sea squirts: A growing global problem. J Exp Mar Biol Ecol 342: 3–4.
42. Boyd MJ (1972) Fouling community structure and development in Bodega Harbor, California. PhD Thesis. University of California, Berkeley, California. 43. Pysˇek P, Jarosˇik V, Pergl J, Randall R, Chytry M, et al. (2009) The global invasion success of Central European plants is related to distribution characteristics in their native range and species traits. Diversity Distrib 15: 891–903.
44. Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, et al. (2000) Biotic invasions: causes, epidemiology, global consequences and control. Ecol Appl 10: 689–710.
45. Theoharides KA, Dukes JS (2007) Plant invasion across space and time: factors affecting nonindigenous species success during four stages of invasion. New Phyt 176: 256–273.
46. Olyarnik SV, Bracken MES, Byrnes JE, Hughes AR, Hultgren KM, et al. (2009) Ecological factors affecting community invasibility. In: Rilov G, Crooks JA, eds. Biological invasions in marine ecosystems: ecological, management, and geographic perspectives. Heidelberg: Springer. pp 215–240.
47. Winne CT, Keck MB (2005) Intraspecific differences in thermal tolerance of the diamondback watersnake (Nerodia rhombifer): effects on ontogeny, latitude and sex. Comp Biochem Physiol Mol Integr Physiol 140: 141–149.
48. Bowler K, Terblanche JS (2008) Insect thermal tolerance: what is the role of ontogeny, ageing and senescence? Biol Rev 83: 339–355.
49. Gosselin L, Qian P (1997) Juvenile mortality in benthic marine invertebrates. Marine Ecol Progr Ser 146: 265–282.
50. Sorte CJB, Stachowicz JJ. (in review) Patterns and processes of composi-tional change in a California epibenthic community. Mar Ecol Progr Ser). 51. Stillman JH, Somero GN (2000) A comparative analysis of the upper thermal
tolerance limits of eastern Pacific porcelain crabs, genusPetrolisthes: influences of latitude, vertical zonation, acclimation, and phylogeny. Physiol Biochem Zool 73: 200–208.
52. Somero GN (2005) Linking biogeography to physiology: Evolutionary and acclimatory adjustments of thermal limits. Front Zool 2: 1–9.
53. Deutsch CA, Tewksbury JJ, Huey RB, Sheldon KS, Ghalambor CK, et al. (2008) Impacts of climate warming on terrestrial ectotherms across latitude. Proc Natl Acad Sci USA 105: 6668–6672.
54. Somero GN (2010) The physiology of climate change: how potentials for acclimatization and genetic adaptation will determine the ‘winners’ and ‘losers’. J Exp Biol 213: 912–920.
55. Sorte CJB, Jones SJ, Miller LP. Geographic variation in temperature tolerance as an indicator of potential population responses to climate change. J Exp Mar Biol Ecol, (in press).
56. Sorte CJB, Hofmann GE (2004) Changes in latitude, changes in aptitudes:
Nucella canaliculata(Mollusca: Gastropoda) is more stressed at its range edge. Mar Ecol Progr Ser 274: 263–268.
57. Fields PA, Rudomin EL, Somero GN (2006) Temperature adaptation of cytosolic malate dehydrogenases from native and invasive species of marine mussels (genusMytilus): implications for biogeographic patterning and invasive success. J Exp Biol 209: 656–667.
58. Braby CE, Somero GN (2006) Following the heart: temperature and salinity effects on heart rate in native and invasive species of blue mussels (genusMytilus). J Exp Biol 209: 2554–2566.
59. Grosholz E (2001) Small spatial-scale differentiation among populations of an introduced colonial invertebrate. Oecologia 129: 58–64.
60. Yund PO, Stires A (2002) Spatial variation in population dynamics in a colonial ascidian (Botryllus schlosseri). Mar Biol 141: 955–963.
61. Grosberg RK (1988) Life-history variation within a population of the colonial ascidianBotryllus schlosseri. I. The genetic and environmental control of seasonal variation. Evolution 42: 900–920.
62. Meehl GA, Tebaldi C (2004) More intense, more frequent, and longer lasting heat waves in the 21st century. Science 305: 994–997.
63. Sorte CJB, Fuller A, Bracken MES (2010) Increase in non-native species dominance triggered by a simulated heat wave. Oikos 119: 1909–1918. 64. Sokolov AP, Stone PH, Forest CE, Prinn R, Sarofirm MC, et al. (2009)
Probabilistic forecast for twenty-first-century climate based on uncertainties in emissions (without policy) and climate parameters. J Clim 22: 5175–5204. 65. Morris RH, Abbott DP, Haderlie EC (1980) Intertidal invertebrates of
California. StanfordCalifornia: Stanford University Press. 690 p.
66. Lambert CC, Lambert G (1998) Non-indigenous ascidians in southern California harbors and marinas. Mar Biol 130: 675–688.
67. Cohen AN (2005) Guide to the exotic species of San Francisco Bay. OaklandCA: San Francisco Estuary Institue, Available: http://www.exoticsguide.org. Ac-cessed 2010.
68. Abbott DP, Lambert CC, Lambert G, Newberry AT (2007) Ascidiacea In: Carlton JT, ed. The Light and Smith manual: intertidal invertebrates from central California to Orgeon. Berkeley and Los Angeles: University of California Press. pp 949–963.
69. Harris LG, Dijkstra JA (2007) Seasonal appearance and monitoring of invasive species in the Great Bay estuarine system. Report to New Hampshire Estuaries Project, U.S. Environmental Protection Agency National Estuaries Program. 70. Locke A (2009) A screening procedure for potential tunicate invaders of Atlantic
Canada. Aquat Invas 4: 71–79.
71. Ocean Biogeographic Information System. Available: http://www.iobis.org. Accessed 2010.
72. Richardson M (2009) A new invader. Newsletter, Padilla Bay National Estuarine Research Reserve, Available: http://www.padillabay.gov/newsletter/Winter09-10.pdf. Accessed 2010.
73. Locarnini RA, Mishinov AV, Antonov JI, Boyer TP, Garcia HE (2010) World Ocean Atlas 2009, Volume 1: Temperature. In: Levitus S, ed. NOAA Atlas NESDIS 68. Washington DC: U.S. Government Printing Office, 184 p. Available: http://www.nodc.noaa.gov/OC5/SELECT/woaselect/woaselect. html.
74. NOAA (National Oceanic and Atmospheric Administration) (2010) National Oceanographic Data Center Coastal Water Temperature Guide. Available: http://www.nodc.noaa.gov/dsdt/cwtg/index.html. Accessed 2010.
75. Canadian Department of Fisheries and Oceans (2010) Lighthouse Data. Available: http://www.pac.dfo-mpo.gc.ca/science/oceans/data-donnees/lighthouses-phares/index-eng.htm.Accessed 2010.