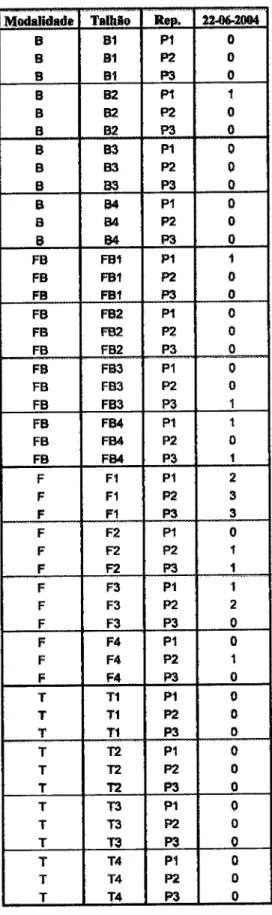
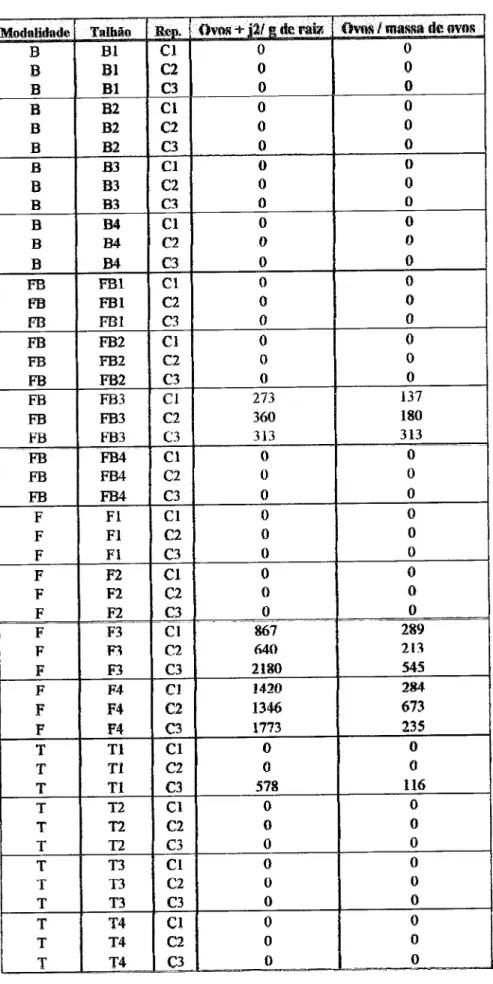
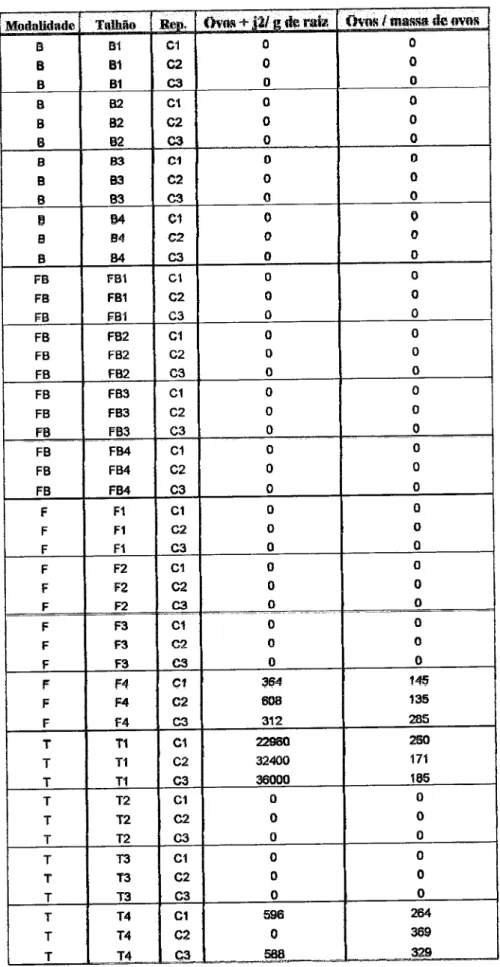
Untvrrddodr
dr
ÉvorEEvolução
do populoçõo
do Íungo
nemotófogo
Pochonlcr
clrlEíüfdospot{o
-holsdo
FcM
R,
qpó§
lnoculqçÊio, no
solo
e
Rs raE
do
lomstelro cullivqdo
em eslufs
Elirsôrtôçüô dê Mêrtrddo
em
Bblogriado
Prrogolr
Doançor da plontcrsorientsdo pclo
Prcf. DoutorMonuel
Melo cMots
l$lono
Rulo Rlbclro GIÕoVlnogro
fuorsmI
Unlversldode
de
ÉvoroEvoluçôo
do populoçôo
do
fungo Rêmolófogo
PoçhonÍo
chlomydosporlo
-lsslqds
PeMR,
opós
lnoculqção,
no
solo
e
nq
rqlz
ds tsmqlelro
çultlvqds
em
eslufo
Dissertoçôo
de
Mestrodo em Biologio de Progos e Doençosde
plontosorienlodo
pelo Prof. DoutorMonuel
Melo eMots
)(,fi
fuo
LlllEnE Bule Elbetro
Gl6o Vlnsgre
Prefacio
Exa
disserlaçaotbi
reaiizadano
ámbito
cioMexraqio
rie
}iriologiade
Fragas eDosnças
de
Plantasda
Liniversidadede
Evora
Êteye çomo
orienmdoro
ProfessorDoutor
lwanueiMçio
eMota.
us
esÍudos que conduziram à eiaboraçao da <iissçnaçiioioram
reaÍizadosno
Laboratoriç de iviicoiogia apiicada (iúÁIvi,l,
UniversiqÍaeiç deEvora e os
ensaiosde
campo
foram
exeçutadosno
Centrs
de
erperimentaçõss do Patacâo (r-aroJ cia ti'uecçào Regionai cieAgriruhura
do Aigarvo.
focio
o uabaiho
foi
finançiado peio
ProgramaaGRú,
e
insere-seno
Frojecto
n'244:
"Acçao do
firngonematófags
Yerttcriirwm chfurnycictsponum sobreo
neuarátocie cias gaiiras raciicuiares,Ágradecimentos
Gostaria de agradecer
a
todas as psssoas que contnbuíram paraa
realização dçstç trabalho, nomçadament§, aoProf. Doutor
ManuelMotq
à
Prof. DoutoraMaria Ivsne
Çlarq
ao Eng.
Cados Franco
e
aos
colegasdo
projççto da DRAALG, Eng.
Jose Entrudo,Eng" Nídia
Ramos e Eng" Eugenia Neto. Gosiaria de prestar, igualmenterútr
agradecimento à Profçssora Doutora Isabel Abrantçsç
Df
ClaraVieira
dos Santos doInstituto
de arnbiente evida
da Univçrsidade de Coimbra pela cedência dedados
Aos meus colegas de laboratorio João do Rosário, Susana Vtagriço e Sérgio DrartÇ, obrigada pelo apoio.Por
ultimo
agradeço aos meus pais e Ricardo Ramos daSilvq
por todo o apoio, inçentivo com os quais tudo se toÍna muito mais fácil.Rsumo
O fungo Poçhonia
chl*rydoryoria
e um potencial agsnte deçontrols biológico
dos nemátodçs-das-galhas-radiculares.Com çste
trabalhq
pretendeu-sç avaliar a efiçáçia de inoarlações ds çlamidósporos no solo, no estabeleçimento de uma população do fungo no solo e na raiz de tomateiroem
estufa
çom níveis
dç
dçnsidadeiggais
ou
zuperioresaos
çonsidçrados somo neçessários para um eficaz controlo dos nemátodes-das-galhas-radiculares.Ao
longo dedois
anosde
ensaio,foram
efecruadas inoculaçõesdo
isolado
PçMR
e
avaliada
adensidade ds fungo no solo ç na raiz.
As
inoculações efççtuadas permitiram
estabelecer
uma
população
de
P.no
soloç
atingir
os valsres de densidade pretendidos.No
e,ntanto, os valores pretendidos paracoloniação da raa
pelo fungo, tbram atingidos
apçnas no primeiro ano.Foi
igualmente demonstrada a capacidadedo fungo
em se mantçrno sols
durante longos períodos detempo
mesmo na ausôncia decultura
ç
em çondições adversas de humidade e temperatura.Abstract
Evolution of the
nematophagrus fungus Fochonia
chlarnydosporta-
PçMR
isolatepopulation, after inoculation, in
soiland root
of a greenhousetomato
culture
Pochania chlamydospona
is a
potential
root-knot
nsmatodebiological
çontrsl
agent.
The
aim
of
this work
was
to
evaluaÍe
ths
effectivenessof
chlamydospore inoçulations at thesoit
for
the establishment at both soil and greenhouse tomato rooLof
a fungus population in
dens§
levels squal or superior to thosç çonsidered as nççdçdfor
ancffeçtive
controlof
root-knot nematode.Along two
years, sevçral inoculations using the portuguçsç isolate PçMR wçre made andthç
dens§ of
fungus atthç
soil and rosts studiçd.Thçsç inoçulations allowed the stablishmçnt
of
a population of P. chlamydosparia atthç soi.t and achiçve the desirsd density values. Elowever, only in the
frst
yearof
assay,the desirçd values
of
root çolonization by funguswçrç
açhiçved.It
was also demonstrated thatP.
chlwnydnsporia çan survivçfor
itsetf atthç
soilfor
a
long period
of time
evenin
the
absençeof
plant
culture andin
advsrsemoist
and tempsrafu re conditions.ínaXe
ÍnOice
Prefácio Agradecimentos Rçsumo Abstract Ínaice Abreviaturas 1. Introdução 2. Revisão bibliográtioa 2. 1. rmportânçia do nçmátode-das-galhas-radiculares 2.2. Cwaçterizaçãodo
géneroMeloidogne
spp.2 "2. l . C lassificação ta<onomica
2,2.2. Caraçtçrização morfologica e
çiclo
deüda
2.3. Caraçtorização da doença2.4. Medidas de çontrolo
2.4.
l.Controlo Biológico
2.5. Çaraçterização do fungo Pochonia chlwnydosporia (Goddard) Zarç
&
Çpms,200
I
Var. chlwnydasporia2. 5. 1 .Çlassificação ta:ronomiça
2.5,2 : Caracterização morfotogica
2.6. Isolados de P. chlwnydosporia
-
oPcMR
2.7. P. chlamydosporia çomo 4gente dç cornrolo biologico 2.7.1. Colonização do solo e
daratz
I
ii
üi
iv
v
1 3 5 5 6 6 6I
10l1
14 L4l5
t7
18l8
[ndíce
2,7.2. Factores que afsstam a oolonizaçãs do solo ç da
rizosfsra
2A2.7.3,
Msdo
de acção de P. chlwnydosparia-
proççsso deparasitismo
22 2.7.4.A
relaçãotritrófica
fungs-nematods-planta
232.7.5" Utilização de
P.
chlwnydosporia çomo agsnle de conúolo dosNGR
263. Mat€rial e
metsdss
293.1. Delineamento
experimental
293.2. Cronograma do
ensaio
343.3.
Metodologia
353.3.1. Produção dç
inoçulo
353.3.2. Froçosso de inoculação no
campo
393.3.3. Cslheita das
amostras
403.3.4. Avaliação da colonização do solo
porP.
chlamydasporia e otrtrosfu4gos 4l
3.3.5. Avaliação da colonização da
ruzpor
P. chlwnydosporia e outrosfungos
42 3.3.6. Contagom das unidadçs formadoras de colonias defungos
423 .3.7 . TraÍasrento esaÍístico dos
dados
434. Apresentação dos resultados e
discussão
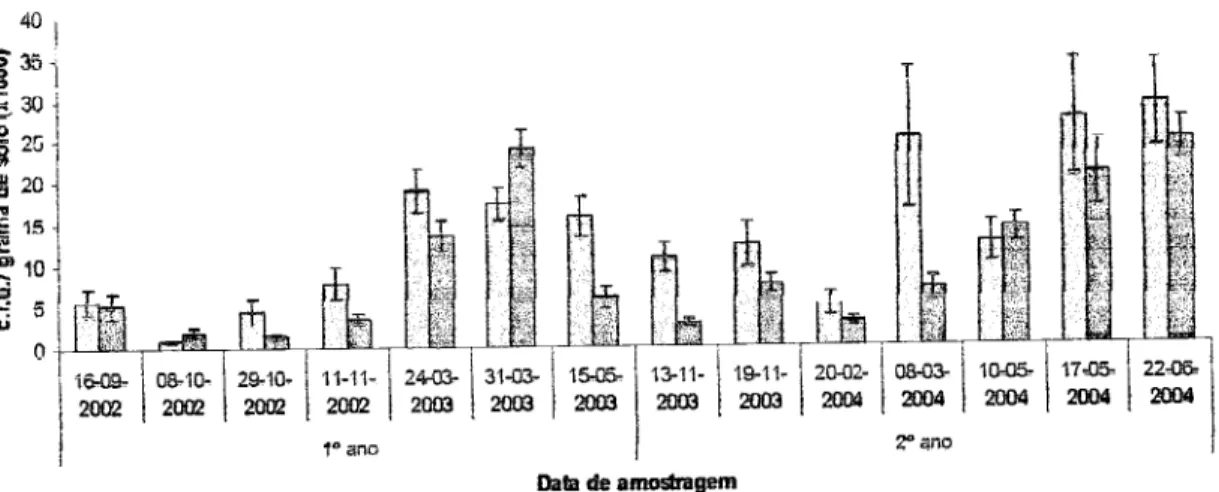
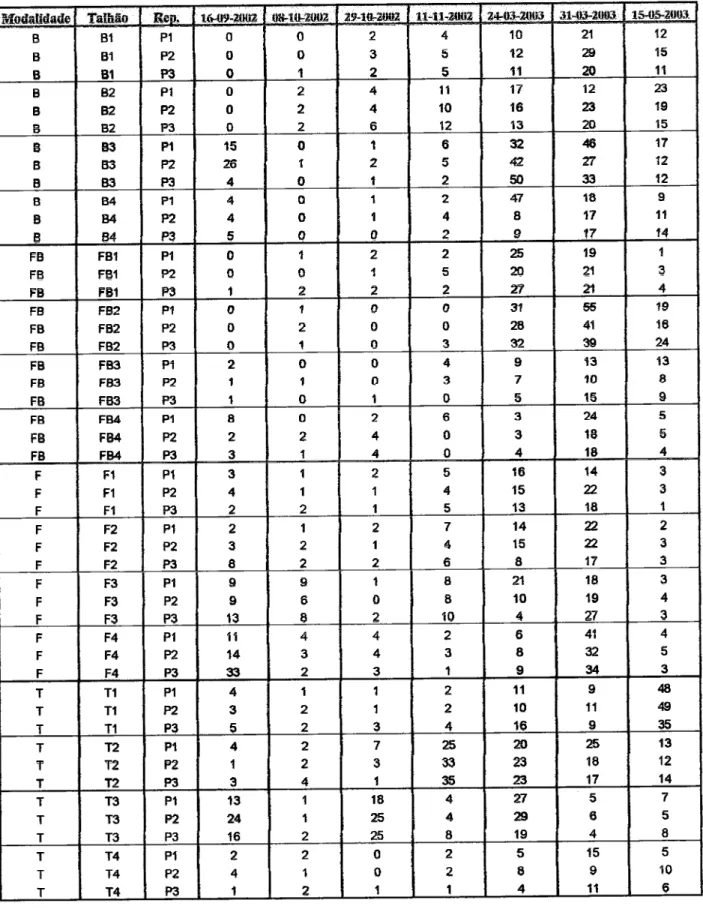
454.1. Avaliação da população
iniçial
do fungo P. chlwnydasporia no solo çm çada anqde
ensaio
454.2. Densidade
deP. chlwnydoryona
imçdiatamçnte após asinoculações
464.3. Evolução do fungo
P.
chlwnydosporia nosolo
48 4.4.Efeito
da apliçação de bioçida na população de P.chlamydo§poria
544.5. População de P.
chlamydosporiararan
554.5.1.
Efçits da
aplicação
de
bioçida
na
oolonização
da
ruz
pelo
fungoÍn{rce
4.6.
Importância
da
çolonização
do
solo
na
çolonizaçãoda raiz
pelo
fungo
P.chtamyda*poria
584.7 Outros fungos presentes no solo rçsisÍcnÍçs ao mçio
M§§
605.
esnçlusões
656. Considerações finais e perspsctivas
futuras
687. Rçfçrênçias
bibtiográficas
70Abreviaturas
Ah[CIVÀ
AnrílisgdevariânciraB Mdatidade dE tmtam€nto Bimida
e.Ês
Unidads formadora de colónia (Colony foming rmi$e.Y. cultivar
CMA
Corn Meal Agar- Agards farinha de milhoIlap
Densidade aprente ds mloIIRAÂLG
Drresçâo Fcgional de Agnqultura do AlganreF Mdalidad€ de tratam€nto Frmgo
rB
Mdalida& de trahmento Fungo + BiocidaICAM
InstitrÍo de ciências agrárias e meditemâricasJovem de primeiro estárlio de nematode
Jovem de wgundo estidis de nemlítode
Jonem de terceiro e$rfulio de nmiítode
Jovem de uuarto dátlio de neüi{tode
Labmic
l"aboratório de micologia apliadaMS§
Meio senri-slestivoNGR
Nemitodedas-galhas-radicríaresPcMR
Isolado do frmgo P- chlamydosporia utilirrdo ne'súemsio
Rêp"
Repeti$oSEM
Microseopia electróni@ de varrimento (§canningelctrmic
Sig.
Nível de signifieância§H§
Staüstic kckage for Soeial §cieilcss J1 J2 J3 J4 1 T Modalidade Tesemunhatlar. Variedade
VT'SA
VácuçflltÍadordesspensõesâquosasWA
Water-agar,meiodeágua+agarl.ldroduSo
1" Introdução
O
nemátode-das-galhas-radiçularesconstitui
actualmentç,
um
dos
maiores problemas fitossanitáriosçm
hortiçultura
protegida(furioq
2005).
Â
utilização
de pesticidas químicos paraa
minimizaçãodo
problematem
sido generalizada çausando elevados impacros ambientais ç risços para asúde
humana e dç outros organismos. Poresta
razão, as
restrições
à
aplicação
destesprodutos
são
çrescentes, tornando-se fundamçntal a disponibitização de meios dç çontrolo ahçrnativos, eficazes ç Çom meno§efeitos
sççundários indesejáveis.Uma
das
possibilidadesque maror
interçsse tem suscitado neste âmbito éo
çontrolo biologico (Àbrantes et al.,2OgZ). O nçmátod+das-galhas-radicularespossui vários
antagonistasnaturais
(Stirling
&
Mankau,
1978; Morgan-jonçseíal,
19S1; Sharon eÍ aL,1,998;Kçrry
&
Bourne, L999;PyrowolakisÀ
eÍal,
2üA2;Kiewniçk
&
§ikora, 2006).
O
antagonista Pochoniachlamydospwia
ç
um destes organismos, considçrando-se que poszuium
çlevado potencial para §€rutilizado
Çomo 4gente dç controlo do nemátodedas-galhas-radiçulares.Na
sequência dç varios trabalhos realizados em lnglaterra(Kerry
&
Crump,
l9E2;Abrantes
et
al.,
ãOAâ| efeçtuou-sea
prospecçãodo fungo
P.
chlamyda;paria
emPortugpl
e
çonstituiu-sçuÍna
colecçãode
isoladss (Francoet
a1.,2001).
Através
dçtestes
in
vitro
e
testes
em
vaso§,
foram
seleçcionadosalguns isolados
cujas çaractsrísticasfisiologiças
mais se adequam àutilizaçãs
çomo agentesdç çontrslo
do nemâtodç-das-galhas-radiçulares, nomeadamentea
capacidadede
parasitar
ovo§i
acapacidade
de
produção
de
clamidósporosç
a
çolonizaçãoda
rizosferade
plantas (Pombo, 2000; Franco et a1.,2001).I)e
entrç estcs, seleçcionou-s§ para estudos futuros, o isolado PçMR pelos elçvados desempeúos nsstçs tçstes (Franco eí a1.,2001).l.lntraduçtu
para que a cficáçia de P. chtwnydosparia ssja elsvada no çontrsls do
nçmátodç{as-galhas-radiçulareü e nççessário garantir que o fungo çolonize o solo e a raiz das plantas, em níveis çlçvados (dç
Lçü
etal.,
1993; Bourne et a1.,1999). Paratal,
é fundamçntal a çompreeagão da dinâmiçado
fungo
no
solo,
apósa
inoculaçãg
assim comoo
modoçomo
a
colonizaçãodo
solo pelo
fungo
afeçtaa
colonização adequadada
raiz
dasplantas.
Estç e um
aspecto flrndamentalpara
a
eficáçia
do fungo como
agente de çorúrolobiologico
uma vez qus sstç depende da capacidadç ds fungo para parasitar os ovos do nçmátode quÇ sç çnçontram ao longo da superficie radicular(I.erj
etal.,1993).
Estç
trúalho
tem como principal objectivo avaliar
a
çfiçaçia
dç
inoçulaçõçs suçessivas do fungo P. chlamydaryoria, no estabçlecimento de uma população do fungo no solo e na raiz dç tomateiro çuttivado çm çstufa. Para tal, pretende-se:.
Avaliar
a densidadçinicial
do fungo P. chlamydasporia no solo..
Avaliar
a variação da quantidade de propagulos do fungoP.
chlamydosporta no solo çausada por cada nova inocutação do fungo..
Çomparara
colonizaçãodo
solo
e
da raiz pelo fungo, entrs
talhõçsdç
solo tratado çem um biocida e talhões sem çste tratamento'.
Avaliar
a
colonizaçãoda raiz de tsmatçiro por
P.
chlamydosporia, apo§ a§ inoculações..
Verifiçar
a
influênçia
ds
nívçl
de
colonização
do
solo pçlq
fungo
P' chlamydosporta na çolonização daraiz
nosniveis
nçççssáriosà
oçorrência de çontrolo biológiço.â
Reusâo Uhtiagrefiea2. Revisâo
Bihtiográfiea
2.1.
Emportância
do
nemátodeda*gathas-radiculare§
As
çspeçiçs pertencentssao
géneroMelaiúrym*
desigoadaspor
nemiítsdçs-das-galhas-radicularcs(NGR),
constituem
um
dos
maiores problemas
fitossanitários çausadssçnr
nemátodçs em todoo
mundo(Kçrry,
1997). Os seusçfçitos
retlsçtçm-sçao
nívçl
da
quantidade
e
quatidade
das prduções
agrícolas
(Eiseúack
&
Triantaphyllou,
1991), sendo responsáveis pela redução çm médiadç
5% da produção mundial. Os estragosçm
situações isoladaspoderãq
tro eotaoto, sermuito
supsriores(Agrios,2005).
Estão dçscritas mais de 80 especies do generoMelaielagtne, seodo
M. incagnitu,M
arefleria,
M.
javanica e
M.
hapla as
especies çonsidçradasdç
maior
importância econsmiça uma vez que são responsáveis porcerm
de 95% das infecções causadas por nçmátsdçs daquele géneroçm
sistemas agrícolas (Eisenback&
Triantaphyllou,
1991; Baiçhevaet
a1.,2002).
A
distribuição mundial
das especies referidase muito
vasta,verifiçands-sç
a
sua presençaem
quasçtodo
o
muodoç
unra grandç capacidade deadaptação a variadas combinaçõçs dç
tipo
de solo e de tempsratura.A
sua presença éespecialmente problemátiça em países
dç çlimas
temperadosa
quentese
dç
lnvernos curtose
pouco rigorosos(Whitheaü
1997;Agrios,
2005). OsNGR
po§§uçm mais de2000
çspçciçsde
ptantas hospedeiras,incluindo
a maioú
das
plantas
çultivadas pertencentesa
farrílias
distintas çomo
Solanaceae, Cucurbitaceae, Leguminoxte,Compositae,
{lmbelliferae
ov
Crucderae
(Kerry,
1997;
§rios,
2005)'
Algumas espscies destes nemátodçs são pdogénicaspara
crrlturas importantesçomo
ççreais,batate sojA
banana, ÍabaÇo,cafrn çanade-açúcar
§
algsdão
(Kerry,
1997).
Oufroproblema
assoçiadsà
infeção por
especiesdo
gçneroMelaifugne
e o
façto
dçâ. FavtsÍ,o BbkrylÉfraa
induzirem
o
agmçntoda
susceptibilidade das plantasa
outros agentes fitopatogénicoseoÍno fungos
e
baptérias originando doenças
coryleles
e
oomequçntemente aumentando os prejuízos nasçulfiras
(Mitkswski &
Abawi,
2A0B).2.2.
Caracterizaçâo
do género
Meloldogme
2,2,1,
Classificaçãotaxonómica
De apordo
çsm
Eisenback&
Triantaphyllou
(1991)a
classifieaçáo ta:ronómica do géaeroMeloidogne
é a seguinte:Reino
Animal
Filo
Nematoda Claese SecernenteaOrdem
Tylenchida FamíIla Heteroderidae Géneno Meloidogyne2.22,
Camcterização morfológica
e ciclo devida
Os nematodes perteneentes ao género
Meloidogme
são endoparasitas obrigatórios,sçççssúando
dç uma
Plantaho@eira
pm
concluh
o
seueicb
de vida
(Hussey,
19E5;
Eisenbaclç 1998). As espécies pertencentes a estegénero
possuem
um
dimorfismosenral
acentuado, em queos
machossão
filifrrmes, redem
entre
1"2a
1,5
Fte' 1.Fh@
&Meloifuw
ry. (§EM) (fcto cedidaPu rarmic)mm de comprimento e 30 a
36 pm
de2- Revrsão Btbtngráfica
diâmetro e se encontram
livres
no solo.As
Íêmeas são piritormesou
ovóides, ÇQm umestrçitamento n& zona anterior, medindo entre 0,40 a 1,30 mm de comprimento por 0,27
a
0,75 mm de diâmetro(!ig.1)
(Agrios,2{i05;
Eisenbaclq 1985). Estas têm uma tbrmade vida
sedentáriae
encontram-setotal ou
parcialmente inseridasna raiz.
Podem reproduzir-se por anfimixia ou por partenogénese dependenrio das condições em que seencontram (Poinar Jr., I 983
;'I'riantaphyllou,
I 985).i-r ,.i:_^,*.^_ *^ i_i^ú-*i^
+4r drtlrrEtrrdr!l+ç !rIELrgr!Hg
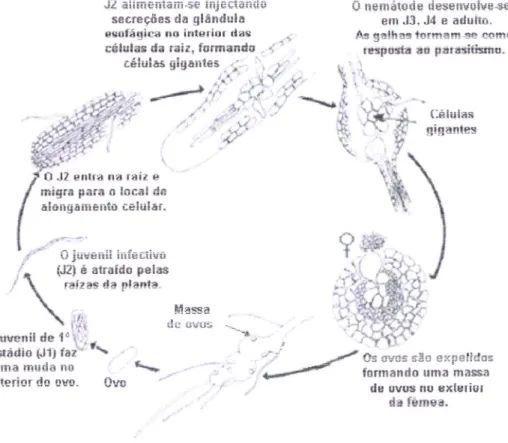
secreçôes da glÉndula erofágica no interior das crilulas da rair, fsrmands "
células gigautes O nemátooe desettuoiue+e em J3. J4 e aduhs. Âr arlhan lnrmtm.* mE* rÊüpottá to para*itlrnu-Céiuias giganles Os c:rcs sãa expelid== formando uma masaa de ovus no exlsrici
da fômsa. O J2 entra na taíz e
migra para o locai da
llongaurerrtt Eelulãí.
?
-*-unra niuila
riu
til
interior doovu.
OvoFig 2. Ciclo <ih vida <1o nematode-iias-gaihas-radiculares (aiiaptailo de Agrios, 2tltt5)
Em
çada postura,uma
têmealiberta
entre 500
a
maisde
1000ovos
quetiçam
envolvidos numa matriz
gelatinosa segregada peias glândulasrectais
(§{itkowski &
Abawi. 2003) que tem como função
a
protecção
dos ovos
da
<iessecação.e
de antagonistas e predadores. Este conjunto riesigna-sepor
Ínassa de ovos e e geraimente depositadono
exterior
da
raí2, podendo,no
entànto,ticar
retido
no
seuinterior
se atêmea
se
enÇontrar profundamenteinserida
na raiz (Agrios, 2005).
A
eclosão do nçmátode depende de tâctores como a maturâção do ovo, a temperatura e o tipo de solo."7
2- Rerasão E ihtia$âfica
O
primeiros
s€gundo çstádiosjuvenis
(Jr eJr)
dçsenvolvçm-ss nointçrior
do ovo.üç
nçmâteiiçs e,çiqrdem sob a ftrrma rieJz,
único
estádio infeccioso(Fig.3),
elibertam-se pâra
o
solo.
Osjovens
de segrrndo estádio.aptos
a
infecÍar
asplantas,
são moveis,
vermiformes
emedem
entre
0^3
a
0,9
mm
de comprimentoe
c€rsade
815
pm
deFig
3. Ovos de Meloidogne sp. e eclosiio dediâmetro (Eisenbach 1985;
umjonreme
segundo estádio (4tn:iXfoto cedidaEisenbaclq
J.D.
& Triaataphyllou
por Laburic)H.Il,
1991).
Ao
encontrarem
umaplanta susceptível, penetram nesta e deslocam-se pelos espaços interçelulares.
Na
zonatio
aiongarneutr;
ççiuiar
iixam=sç
einiciam
o
processode
aiimentação pelaintrodução
de
secreçõesda
glândula esofagica nas células claplanta,
atravésde uür
estilete.
Como
resultado,
ascelulas
de
alimentação
permanentesFig. 4. Galln na raiz corn jovern causada
por
sofrem
hipertrofia
e
ttlrnam=seMeloidogne
§p'
(40ox) (foto cedidapor
multinucleadas designando-se
por"ççlulas gigantes".
Originam-se
assim
as
galhas
radiçulares
típicas
da
doença (Eisenback&
'friantaphytlou,
1991; Bleve-Zacheo etal.,
1997). Seguidamente, dá-se urna neva muda que origina umjovem
de terçeiro çstádio (J:). Esta fasç e curtadando-sç a
terceira muda
logo
de
seguidae
originando um
jovem de
quarto
estádio (J+).Durantç o terceiro e quarto estádios o nemátode não se alimenta. Com a quarta e última
2
Revrsão Bbfngráfrcamuda, o nemátode atinge g estado adulto. Este poderá sçr um maçho vermiforme que §ç
tiberta
daraiz
c vive
livremçntç
no
solo,ou
uma Gmea que continuaa
aumentar devolumç e quÇ adquire forma
piriforme.
Çomou
semfe*ilização
do maÇho, esla produzovos
que Libertade
seguida para umamatriz
gelatinosa(Mitkowski
&
Abawi,
2003; Agrios. 2005).O
ciclo
rievida
dosNGR tem
a duração aproximada de 25 dias a 27"C,
podendo durar mais tempo, dependendo da temperatura.A
dispersão dos nemátodese tbita
principalmente atraves da águae utilização
dematerial agricola
contaminario.
A
maioria dos
nemálodes encontra-§eno
sistema radicular entre 5 a 25 cm abaixo eia superÍicie do solo (Agrios, 20Ü5)'2.3.
Caracterização da
doença
Os
sintomas
mais
saracteristiços
e
tlramâticos
resultantesda
infecção
pelos nemátodes-das-galhas-radiçularessituam-se
ao nivel
do
sistema
radicular.
Comoresultado
rJaalimentação destes,
as ralzes dilatam
no
ponto
de
infecção
e
sãoriçsenvoivtiias gaihas
iipicas
ua doençaque
podem
atingir
varias vezs§
odiâmetro
de
uma
ruiz
saudável§{itkowski
&
Abawi. 2003;
Agrios'
2005),
f
conlum
oÇoÍrÊrem várias intbcções ao longo daraiz
eont'enoelo-Fig.
5.
Raiz
corn
galhas causadaspor
lheum
asp€cto rugosoe
sinuoso (Fig'líeloidogtne
sp'(fotocedidaporlabmic)
5).
A
infecção
causadapor
algumas especies pode tambeminduzir
o çrescimento de pequena§ mas numerosas raizes na zonÍt acima da galha, resultando numa densirJade anormal de rarzes secundarias ne§§a zona'2. RevÍsão Wt@rffiea
Em geral, çomo resuhado da infecção por nsmátodss
do
géneroMeloidogmeo o sistçÍna radiçutartem
Çresçimedo re-duzidoç
psde,m dese,nvolver-§e nsçTose§(Agrios,
2oo5).Na
parte aérça das plantas infsçtadas é Êequente obsçrvar-sç o çresçimçnto reduzids eurrra m€nor guantidade dç folhas, gue sm gcral são de mçnores dimensões relativamente a uÍna planta saudável e de coloração vçrdc pálida
ou
amarçlada. Os rebentosç
frutos eçoÍrem igualmente çm menor quantidade e são de qualidade rçduzida. Çom o aumentoda
temperatura, as folhas tendem aindaa
murchar. Estçs sintoma§,as nívçl
da parte aerça da planta" oçorrçmdçvido
à absorção etmsporte
limitado
de água e nutrieutes para os nívçis superiores da planta quÇ! por sua vç2, rçsultam da existênçia de galhas na zona radiçular que impedemo
ssufluxo
normal. Por esta razão, a planta pode também apresentar sintomasdç
çarênçia de nutriçntçs.A
gravidade dçstas alterações depende,em geraf
de
três
façtorçs:da
densidade populacionalde
nemátodçs,da
especiç do nemátodç e da susceptibilidade do hosps.deiro (Whitehead, 1997;Mitkowski
&
Abawi, 2Oo3).2.4.
Mdidas
decontrolo
Existem
diversos mçios parao
controlo
de nemátodes fitoparasitas sçndoo
custo, eficiáçiae
o
tipo
de
cultura" algunsdos
factsrçsqus inÍluçnçiam
a
çsçolhads
mais adequedo(Agrios,
2005). Emgeral
e aplicada a çombinação de vá'rios métodos çomo asotarização,
o
use de
plantas
rçsistentçü
rotação
com
plantas
não
hospedeiras, incorporaçãode
materia orgânica &esçae
aplicaçãode
peslicidas (Whitehead, 1997;Agrios,2005).
Entre os vários meios eüstentes, a aplicação de nçmatiçidas quíruicos é o meio mais
fiável
e por isso um dos mais utilizados no controlo do nemátçdçdas-galhas-radiçularçsem
váriostipos
de
çuhuras, aBssaÍda
sua aplicaçãotçr
um impacto
considsrável ao2
RâtrS6oMtirytua
nível
ambiçntal, eçonómiço
ç
da
saúde humana.
Os
principais
efeitos
negativos resuhante-sda
suautilização
rçflççtem-sea
nível
geaetieo,psderdo
eausar anomaliasmorfológicas
e
fisiológiÇasçm
animais
as nível dss
sistemasnçrvoso! respiratóriq
irnunitáriro ou sndóGrino
(EP{
2006).A
bioacumulação, a persistência das substânciasnos
ççossist€masoa
acumulaçãonos
lençois
frerátiços,o
arrastamçntodos
produtos pulverizados pelo vento e alixiviação
são outros dçs riscos assoçr,ados à aplieação de nematiçidas quimicos. Pode rçferir-sç como exemploo
bromçto de metilo,um
bioçidade largo espectro muito utilizado em
hortisrltura
intensiva.E
urnaubstâneia muito
volatit que
facilmentç
sç
libçrtada para
a
atmosfçra
sendsum fortç dçstruidsr
da camada de ozono. Por çsta ra?Ão, de acordo Çomo
protocolo deMontrçal (1995)
serárçtirada do merçado atç ao ano 2010
(Kçrry,
2001; Portal da União Europera, 2006).A
çrçsççntç preoçupação çom a saude humana e ambientat devido ao uso de alguns pesticidas guímicos, aumenta o interessç pela pesquisa ç desenvolvimçnto de estrategias que integrem metodos de conlrolo variados e que reduzçnn a dependência dç substânçias químicas(Kçrry,
2001).2.4.1.
Controlo Biológico
Q controlo biqlógiÇo é
dçfinido
çomo o uso de organismos vivos Çom o objectivo dereduzir
a
densidade populacionalou
impaotode
dçterminedo organismo patogénico, tornando-o mçnos abundanteou
mçnos destrutivo rçlativamçntç ao quç açorúeççria na ausênçia do agente de çsnuolobiologico @ilenberg
2006).Os nemátsdçs fitoparasitas, em particular o gsnero
Meloidagme,
dçsçnvolvem umacomplexa retação
çom
o
hospedeiro
e
o
s€u
ciclo de vida
ofereçe
variadas possibilidades de perturbação. De fonna a çnçontrar um agents dç çontrslo biologico e adesenvolver oovas estratégias para
a
suautilização,
torna-se necçssário determrnar e2
Revrsão BiÉ,tiqráfieaestudar os anta€ooistas naturais quç existem
no
solo ç que podçm s€r respoosaYeis pelaeventual
suprsssão daspopuhções
de nematdes.
É,
no
entanto, neçassarioter
cm consideraçãoque
o
contrslo bislogiço nãs
pode sçr
visto
como
um
substituto
dss pesücidasquímicos,
ou
somo
um
modo
de
eliminação imçdiata
dos
organismos fitopmogéniÇos uma v€z que tem que sçr mantida a fonte dç aliments numçcrts oívçl
permitindo que o agsote de çontrolo biotógico possa sobreviver e multiplicar-s€. Devçrá antescsr
ençaradsçomo
um
compleÍnsnÍoa crtros
métodgs, numa perspectiva de protecção integradana
gestãoda
populaçãode
nsmátodese
prsveoçãode
futuras infestações graves da doçnça(Kerry,
1997),Apesar de possuir atgumas limitações, Gomo
a
lenta
actuação dos organislno§r aforte dependênçia das condi@es ambientais, ou a difiçuldade em atingir a totalidade dos
organismos patogénicos,
o
çontrolo
de
doenças
por
meios biologicos
apresenta vantagens na suautilização çomo a
especificidadedo
hospedeirÇoo
controls a
longo praza Ç o mener impacto a nível aurbiçntal, ecológico e da saúde humana(Kçrry,
1997).Um
agentede
çsntrolo biologico
de
nçmátodçsdevçrá
cumprir
os
seguintes requisitos: capacidade parasíticaçlçvadq
ter o
nemátode çomopnncipal
hospedeiro enão ser
patogenico
para as
plantas cultivadas
ç
outrss
animais.
Dçvç
ainda
ter
crescimçnto optimo em intervalos adequados depH
e tempsratura para a sua produçãoem
mass& desenvolvçr-§ççm mçio artificial, ter um
msdo de
acçãocoúecido,
serpossívet
formutá-ls
mma
forma
utiliável
e ter
caBacidadede
competir çom
outros microrganismos autóçtonçsds
sistçmaondç
e
introduzids (Sidiqqui
&
Mahamosd,lee6).
Açtualmente
já
são produzidos comercialmente alguns agçntçs dç çontrolo biologicocottaMelaidagtne
spp., no entanto, estes ainda não demonstraram a çfiçáçia sufiçiçnte2
Rer,nsâo Bibliogrúeapara se tomarem uma
alt$nativa
aos mçios deçontrsls
químico açtualmente exi§tçotç§ para a rnaior parte dos sistemas de produção de ptantas (Abrantes et a1.,2002).Os antagonictas dç nsmátodes ütoparasitas podçrão sçr vários çomo parasitas (vírus,
ba«erias,
protozoánios,fungos)
e
predadores (protozoários, áearos,
tardigrados, çolêmbslos, fungose
outros nsmatodes)(§ayrç
&
I#altçr,
1991;Pyrowolakis et
al-, ZAAZ1Dç
entrç os antagonista§, os fungos ooupamum
lugar de destaque(§idiqqui
&
Mahamood" 1996). Estes podem sçrdivididos
çm três categorias, segundoo
§çu msdode
actuação;i)
Fungos formadores
de
armadilhasque
podem
sçr
redesde
hifas, ramificações, botõos adesivos, ançis çonstritoresou
não constritores (ex.ArthroboÍrys
irregularis, Dactylella
spp. §Morncrasporiam
spp.) (Jatala, 1985;Kerry,
2001;Kerry
&
Jaffee, 1997);
ii)
Fungos
endoparasitasdç
nemátsdesvçrmiformes
que
são çaraçterizados peta produção de esporos adçsivos de pequenas dimensõçs que aderem à çutíçula do nçmátodç e penetram:na, colonizandoo
hospedeiro ç zoosporos mÓvçis ou §sporosque
nça€§sitamde
ser ingeridos parâ que
possaosoÍrçr
a
colonização dohospedeiro
(ex.
Drechmeria coniospora, Catenaria
anguillulae
e
ÍIirstella
rhossiliensis)
(Viaene
&
Abawi,
2000;
Jansson&
Lopezdlorçq
200a);
iü)
Fungos parasitas de ovos ou formas fixasdo
nemâtode quç possuÇm a capacidadç de parasitarovos
GÊmeas
sedentáriasde
nemátodes titoparasitas.As
especiesinçluidas
nçsteúltimo
grupo
sãs üequçntçmenlç çncontradasçm
solos agríçolase
são çonsidçradas potenciais agentesde çontrolo
na medidaem
que possxama
eapaoidadçde
inibir s
proçessoreprodutivo
dos nçmátodçsou a
ççlosão dosovos. Qs principais
meios dç acçãoçnvolvidos
So
a danifiçação de elementos estruturais dos ovos e da çutícuta donemáXode por actividadç enzirnátiça ç/ou fisiça ç distúrbio
fisiologico,
através da síntesee difusão dç
metúotitos
toxicos (Morgan-Jones&
Rodriguez-Kabane 1987). Foramjá
isoladas várias especies de fungos parasitas dç Íêmças sçdentárias, desvss
e de quistosE R€MSâO Bib@ráfrCa
de
nematodss,no
entanto apsnas algumas GoÍno Paecilomyceslilaçirrus e
Poehoflia ehtawrydospariatêm sido e.sfudâdas mais iffeosivamenta(fçrrl,&
Jaffee, 1997;Kerry,
199?;
Kerry,
2000). Uma das grandcs ventagços quç sstas çspççies aprç§sntam é o façtode
seÍem
parasrtas
façuttativos, possuindo
a
capacidade
de
se
alimentar saprofiticamentç. Destaformg
a
sua sçbrçüvênçia não
seçnçontra
completamente dependente da presença de aemátode§ tro solo(Kerry,1997;
Kerry&
Jaffçç, 1997).2.5.
Caracterização
do
fungo Pochonia chlamydasporia
(Goddardl
Zare
&
Gams,2fi)1 var.
chlanrydosporia
O
fungo
Pachonio
chlamylosporia
tbi
iniçialmçnte idçntiÍicado
em
l9l3
por Goddard apos este ots
recolhidodo
seujardim
(Ocq
2004). E um fungo endoBarasita dç formas sçdentárias de nemátodes fitoparasitas ejá foi
isolado dç ovos dç nçmáIodçsdos quistos (Heteroderú, spp.
e
GletbuÍera spp.), de oernátodes-das-galhas-radiçulares em variados loçaisdo
mundoe
embora raramçntç,dç
ovos de çaraçol(Kçrr)',
2001;Zwç
et
at.,
2AA\.
Trata-sedç um
parasitafacultativo com
uÍna
vasta distribuição ençontrands-se um pouço por todo o mundo(Kçrry,
2001, Àbrantçs ela1.,20A4.
O
fungo
P.
chlamydasporia
tem
sido
eaçontrado
eÍn
solos
srpressivos
de nemátsdçs-das-gathas-radicularçs e nçmátodes dos quistos çonsidçrando-sç gue possuium
potenciat csnsiderável quandointroduzido em
solos infestadsspor
estas especies (Kerry,&
Crump,
L977; Morgan-Joo§§ eÍa/.,
l98l;
Kerry
eíal.,
19824;Kçrry
et
al-,19S2b;
Kerr),&
Crump, 1998).2.5.1. Classifieaçâo
taxonómica
Taxonomicament§,
e
anamorfoPachonia
chlarnydosporiaestá
incluído
no
filo
Deuteromyçoüa
ç o
sçu telçsmortb,
Cordiceps
chlamyfusparia,
psrtençç
ao
filo
2
Remsão BibliogrfficaAscomycota
Estg foi
sbsçrvado sm ovos de mslusços na Amçriçado §ul, ns
çntantoas observaçõçs descritas da fase senrada do fungo são raras (Zwe et a1.,2CI01)'
Dentro do
género Pochoniaçxistçm pelo
menos mais duas espéciçs assoçiadas aovos dç nemátodes, a çspçciçP. suchla"sparia e P. rubescens íZare et a1.,2001).
A
especie
P.
chtmtydasporia ençontra-ss
dividida nas
variçdades
P-chlarnydasporiavw.
chtarnydasporia e P.chl*nydoyoriavar.
catenulala cuja principatdifersnça residç
no
modo de
formação
dos
çsnídio§,
çm
aglomeradosglobosos
e rnucilaginososno
casoda primeira
variçdadçs
sm
çadciana
ssgunda(Zwe et
al-, 2001).Desdç que
foi
idçntifrçadapela primeira vez
sm
1913foram
atribuidas diversas ctassificaçÕesç
nomss a çsta especie (Barron&
Onions,
L965; Zwe et al.n 2001). Em 2001foi
realizadq por
Zare, Gams e Evan§" a revisão da especie naqual
sçatribui
o nome dç Pochanio chlamydosporiaçm
substituiçãode YerÍicillium
chlamydosporium, denominaçãs ainda usada frequentomente pêre esê especie (Zare elal-,2001}
f
ercomorfpJf,ate=*lggOat Reino: Fungi§ilo:
Ascomycota Classe: Âssomycetes Ordem: H5,pocrealmFamlli* Claücepitaee
Género: CordycepsEspécie: C or@ceps chlamYdosPoria
Aapno$q$F.sÊ tssgç+ua$al Reino: Fungr
Filo:
Danteromycota Classe: Hyphomyeetes Ordem: Mooiales Familia: Mmilia§@Ê Género: PaclnnisEs@cie Pochonia chlamYdosPoria
Variedade:
Pochonta chlamydosporla var. chlamydasporia(Zare
et
a1.,2001;Kirk
e,al.,2A$l)
2. Revisão Eibttográfrca
2,5.2.
Caracterizaçâo morfológica
Na ibrma
assexuada desta especie conhecem-se doistipos
de esporos: os çonidiosdo
tipo
tialosporos, geralmente designados apsnâspor
conídiose
os conidiosdo
tipo aleuriosporos, frequentemçnte designadospor
ciamidosporos (Gams, 1988; Zare et al., 2001).Os tiaiosporos são produzicios em hit-âs
ve*iciiadas
(conidioforos) e possuem cercahig. ú. (a,y i'. ctriunrytiosporrrl. a -+izuni.iosptiro. h - + liuiirics nutt verltciir,. r: - liaiosporos cru agrupanlcnios mucilaginosos. (400 x) (bi1 P. chlamydospoia ern rneio de agar dÊ exúacto de malte. (c) clamidósporos de P.
chian&dosporia (§jii\4) { iotos ceriiilas por Labmi": I
cie
2.5-4.5
pm
:;
1.2-Z.Zlrm.,
apresentamtbrma ovai
ou
subglobosae superticle
ilsa {Zare etai.,2üüil.
Us
conidiotbros sào
geraimente
prostraclose
produzem
tialides
srmpÍes
ou veniciiadas com 2 oui
tiaiides ao longo do conidiotbro, ou conjuntos terminais de 4 ou5
tiaiicies. São estruturas t-rnas e tubularescom
Çercade
l2-2ó
pm
de comprimento ei.ü-i.5
pm de diâmetro lLarc
et
ul.,2OA1,i
e
possuemnas
suas extremiciacie§ ostlaioconidios em agregados giobosos mucilaginosos {Gams, 1988; Zare el
aL..20üii.
Os ciamidosporos são espor<;s de pare<ie espessa e possuem cerca
de
15-25pm
cieciiâmetro" Contêm
no
seuinterior
substàncias cie reservaque
ihes possibriitam umaviabiiiciadç mais
proiongada relativamente
aos
tialosporos
e
permitem
ao
tungo sobreviver em condições adversas (Zare elu\.,2ú0i).
São produzidos em hastes curtas2. Revrsão &btngráftca
(pequenos pedúnculos)
com çsrça
de
15-25
pm
de
comprimentoe
consistem numae$rutura
hialina formadapor
um grupo cie6
a9
celulas com paredes quÇ vão írçancioÇspessas
ao
longo do tempo.
Estes podem ser produzidos em grande quantidade nomiçétio
aereoou,
em menor quantidadeo submersos emilgar
(Gams, 1988; Zareei al.,
2r)rii).
As
hifas vegerarivas são hiaiinase
septadase
geraimenteo
micelio e
aereo etino
(Gams, 1988; Zare et a1.,2001).As
coionias do tirngo em meio de cuituraem iÜ
diaspossuem
entre
2ü
a
38
mm rie
diàmetro. lniciaimente são
brancas
adquirincio posteriormenteuma
cor
begee
um
aspecto puiverulento que resultaria
prociução declamidosporos
(Zare
e'tui.,
ZUOI).A
parte interior da
coionia
varia
entre
o
bege,amareio paiicio e iaranja.
2.ó.
lsolados
deP.
chlumyiittsprtria
-
o
PcMR
Existe uma grande variação de algumas caracterrsticas enire tsolados
do
fungo P.chlamydosporia
var.
chlamydosporiu (Abrantes
d
ui.,
20ü2).
As
caracterlsticas reiacionadascom
a
potencialidadedo
i'.
chlamydosporua Çomo agentede
controio bioiogico variam com os dit"erentes isoiacios(ln'ing
&
Kerry,
1986;Kerry
etai.,
i98ó,
Kerry, 2001i
como a velocidade rÍe çresçimento(Kerry et
ai,
1986. Crump &,trving.
lv92;
Kerry,
i995i,
capacidacie parasítica(lrving
&
Kerry,
198Ó;Crump
&
lrving,
lgg}),
temperaruraoptima
a
que
se reiacionamcom os
outros organismos (.Stiriing,lyyi),
Íemperatura optima de crescimento(Kerry
etai.,
198Ó; Crump&
ln'ing,
Lv92.;Kerry.
1995) capacidade de proiil'bração na rizost-êra(Kerry,
1995; Bourne etal.,
i99Ó), prociução de esporos (Kerry etai.,l98ó;
Stirling,
1991; Crump& ln'ing,
L992).Ate
a
actualidade
tbram
ja
recolhidos
<iiversos isolaciosdo
fungo
Pochtmia chiamydosporic. Aiguns destes têm sicioalvo
de estucio paÍa uma potencial utilização2. Rewsão &bttegrâítca
çome agentçs ds çontrolo
biologico
{kving
&
Kerry,
198ó;Hidalgo-i}iaz
eíaL,
200ii; Francoet
al..Z00U.
§enas
um
isolado que colonize faciimente a rizosfera e parasire rapidamente a§ ma§sas de ovos será capaz de çontrolar as populações dos NG-R (Kerry, 2001)Em portugal reçolheram-se varios isolarios do fungo
P.
chlamydosporla que t'bram submeticios a tesres cie selecção baseados na capacidade de parasitar ovostio
nemàto<ie Meioidog,tne spp., produção de ciamidosporos e çolonização da rizosfera de piantas em vasoe
in
viira.
Entreos
isoiados testados,o
isolado PgN{Rfoi o
que âpresentou ascaraçterístiças mais ariequadas
a futura
utilizaçãocomo
agente ciecontroio
bioiôgico contra os nemátories do géneroMelaitiog;ne,
tendo onginado diminuições significativas na popuiação de nemátodes relativaments a testsmunha (Esteves et al-" 2{-lui: Franco eÍal.;7üüi.:
Louro etal-,2001).
2.t.
P.
chiamydospttria
como agente
detontrolo
biológico
2.i.1.
Colonização do soio e daraiz
O fungo P. chlawydosporia é um parasita tàcultativo e não depende exciusivamente
tios
nematoties
Para
sua
nutnção(Stirling,
i99i;
Bourne&
Kerry,
?000)' Para aiémde
conseguir parasitar outros organismos, colonizao
solo deviclo à stracapacidade
saprofitica
quê
o
permitedegradar
uma
grande variedade
demoléculas
incluindo
a
quitina
€
ecelulose.
Tem ainda
a
capacidade deÍig
?. P. chiamldospona sotrre a rau {§llM,)(foto cedida por Lahmic)
2- Rewsão Bibliográfrca
proliferar
na rizosfera de diversas espesiesdc
plantas (Bsurne&
Kerry,
2000;Kerry,
2001).
Em
ged
ns
sslo
oçelrem todos
os
sstádios
asseruadosdo
hrngo
Pochoniachlarnydaspria.
Os çlamidosporos são estruturas de maior resistênçia relativamçnte aosfialósporos e permitem gue
o
fungo sobrçviva durante algum tempo quandos
matçrialorgânico
disponívelno
solo
e
os
nçmátodes hospedeirosSo
çsçassqs.A
relação de parasitismo entre os nçmiátodçs e a planta induz o fungo a produzir maior quantidade declamidosporos, demonstrando
a
importância
destetipo
de
esporosno
processo de desçnvolvimento do parasitismo dç fungo sobre o nemátode (.âbrantes et a1.,2A02;).Quanto à disporsão
do
fungo, sabe-sç que, emboralimitadq
édevidq
ao transporte de conidios pela áEua, ssndo a humidadç um façtor importante nacolonizaSo
do solo çda
raiz O
movimento
de
organismos
inverteAradosdo
solo permite tambem
adisseminação
dss
propágulosdo fungo a
distânçias consideráveis(LçU et
al.,
1993; Abrantes etal.,
2OO2;).Uma
boa colonizaçãodo
soloe
fundamçntal para assegura.rs
çstabçlççimçnto do fungoP.
chlamydosparia na raiz numnívçl
que providençie a reduçãosignifieativa
da populaçãode
NGR
(Lopez-LloÍç,aeÍ
al.,
20[{);
Esteveseí
a1.,2001).
eom
basç çm trabathcs realizados anteriormente çonsidera-seum
solo bem colonizado pelo fungo P.elolmydaryaria
quando este s€ encontra acimade
104 c.f.u. deP.
chlumydoqoria
pargrama
de
solo (medido em meio
semi-seleçtivo).No
çasoda
rabna coloniza$o
naordem
de
103c.fu.
por
gramade raiz
foi
zuficiente
paraprovoçar
a
reduÉo
da poputação deNGR
açima dos
80oÁ, relativamentea
plantas çultivadasçm
solo
não inoçulado çomo
firngo.No
çntantg
çm trabalhos realizados antçrisrmçntsfoi
possívelobter eolonizaÉo da
raiz
acimade
lOac.f,u.
por
graÍna deraiz
çom
inoeulações de10000 clamidósporos
por
gramade solo (de
Leij
et
al.,
L992b1'de
Leü
et
al.,
1993;Z Rêúsilo Bibliogréfrca
!,:t: :: :l:ll::: :l::::: :11:
:-Bourns
&
Korry,
1999).
Para que
seja possível
a
limitação das
populações deMetaifugme
spp., apos a inoculação do fungo no solo, este devçrá GrÇsççr e çolonizar aúosfçra
daplantq
em particular as galhas radiçulares induzidas pelo nernátode (deLeij
et
al.;1993).
Está demonstrado quso
contols
dos nemátodçs-das-galhas*adiçulares émais
efiçaz
sm
raízes
abundantemçnte
çolonizadas
por
P.
chlamydosporia,çomparativamçntç a raízçs em guÊ este e Çscasso (Bourne et a1.,1996).
Ao
çsurário
doque se psnsou iniçialmçnte, o
fiingo
além de sç dçsenvolver apenas na superficie da raiztem
çomportamsntosndofitico, ou
seja, penetra nas celulas epidérmiçase
cortiçais darai4
mas, não çausa nwrosôs ou descamação nem parççe afeçtar negativamente a planta (Lopez-Ltor ca et al., 2000; Bordallo etal.,
2OAZ).2.X.2.
Factore
que afectam a cotouizaçâo do solo e daraiz
A
colonização
do
sslo
e
raiz pelo fungo
P.
chlamydasyxia está
forternente depsndentedç
façtores biotiços
ç
abiotiççs uma
vez
que
estos
afççtam
a
§uasobrevivênçia e proliferação
(Kerry
&
Jaffs§,
1997)"Q
cresçimçntodo fungo no
solo está relacionado com otlpo
dç te&ura deste ç solss ricos em matçria orgânica permitemum
maior
desenvolvimento deste relativamentea
solos areoosos e pobresçm
matériaorgânicq
uma vez que têm disponível uma maior quantidade de alimçnto parao
ürngo (deLeij
et a1.,1993). Qs solos rnais ricos têm, oo entantq, maior abundânçia de outros microorganismos gue podem influçnçiar o çstabçlççimçnts do firngo na mçdida em que poderão competir çom este (Bourne&
Kerry,
2000).Para alem
da influênçia do
outros organisrnos sabç-sçgus
o
desçnvolvimçnto dofungo no
solo
e
tambcm
afeçtadopor
factoresúiotiços.
Estes ençontram-§e pauco estudados, conhççendo-sea
sua influência
apsnas
para alguns façtores çomo
a humidade e tsmpcratura.2. Reusiro Bbliográfrea
A
invasão das raíz€§por
nerniátodesjovens do
sçÊundoçstídio
é reduzida no ça§o de baixos níveis de humidadeno
solo, ma§o
fungo tema
caBacidade deproliferar
no sqlo em níveis dç humidade quç rsduzsm a invasão por parte dos nçmátodes (Bourns&
Kerry,
2000).
Assiq
a
efieaciado fungo
no
çontrolo de NGR
poderá sçr rnaior emsolos
menos húmidos,§
@n§€qusntemente,çom
msff)rçs
densidadçsdç
nemátodçs (Bourne&
Kerry,
2000).A
temperatura é outro facror detçrminantç paras çtabçlçcimçnto
ç çresçirnçnto dofungo
soloç
rizssfera. Estapde
influençiar
o
srescimçnto do fungoe
a produção de çonídios. Em tçstçs invitro
vçrifiçou-se que P. chlwnydanpana tçm çresçimento optimoçntrç 25 o 32"e,
açimados 35"C não
se observa crescimento. Gams(1988)
rçfere aproximadamente 10oC como a temperatura mínima de çrescimento do fungo. Quanto à produção de conídios,o
pico máximo
situa-se nos 22"C
e
entre as22
e
30oC e§ta ç reduzida (deLçü
etal.,1992).
Uma
boa proliferaçãodo
fungo naraiz
está dependentç nãoso
da capacidade de colonizaçãodo
isslado mas também em grande partç da espscie e variçdade da planta hospedeira que podorá possuir maior ou m€noÍ a,ptidão para suportaro
çrescimento dotirngo
(Bourne et a1.,1994; Bourne eí a1.,1996; Bçurne&
Kerry, 2000). Por exemplo, dsterminadas variedades de toÍnateiro têm uma aptidão maior para que a$ta
rizosfera seja colonizada pelo fungo que plantas de alfaçç (Esteves et a1.,2001).Na
rizosfera,
o
crescimerúode
P.
chlamydospria
pareceestar
grandemente relacionadoçom a
composiçãoe
quantidadçdç
exsudados libertadospsla raiz.
Estçsvariam
com a especie daplanta
como
seu estadogerd,
quepor
suavez
depende de façtores Çomo humidadç, temperaturae
luminosidade(BouÍr&
etal.,
1996; Bournç&
Kerry,
2000)"A
libertação de exsudados pela raiz é ainda modifiçada pelo rompimento2- Rewsáo Btblagráfna
da
sua superfiÇiç causadopela
po§luradas
ma§§a§dç
ovos
pelasftmea§
de
NGR
(tsourne&
KerrY, 2000).2.7.s.Modo
de acção àe P. chlamydasporia no proce§§o deparasitismo
Um
aspecto crucial para a esçolha eutilização
deum
isolado dofungo no
controlo dos NGR e a ÇapaÇida<ie dç para§itar o§ oYo§ de nemátode. O fungoP'
chlamydosporia não produz toxinas nem armadilhas como aconteÇe em algumas §species de fungos e aparentcmenÍç nào possui a capauidatiede
parasitar estádios
móveis
denemátodes
(Kerry
&
Jaffee,
1997;Kerq,, 2001)
Em
geral'
o
fi-rngoprolifera
tr&
rizosfera, crescendc
ôdrsoersando-se na
matnz
geiatinosa dasfilassas de ovos
à
medida que estassão
Fig. 8. P. chiam.v-tlosporia a para§tar ovo deproduzidas na superfic ie
da
raiz.Quan<io
NGR (sElví) (foto cedida por Labmic)as hitas
vegetalivas entramem
contaatocom
os
ovos
do
hospecieiro, aderema
suasupertlcie
(Kerry"
1995; Lopez-Llorca elsi.,20{.xl;
Morton et ai.,2OO4).f
orma-se entãouma
estrutura, denominada apressorio,uma
hità
especíafizada formadaterminal
ouiaiçrairnçntç
a
partir
dç
hiias
nãtrdiferenciades
e
quetem
como
funçãoaderir
e
penetrar
nos
hosPedeirosatravés
de
nma
hifa
on
c;r_rnha de penetração (Lopez-Llorca etal.,
2C0A;Kerry.
20{)}}.
APos
a
Penetração"Iig.
9. P. ehtamydosporia a parasitar massa de ovos rle NGR (SEI\{) (foto ccdida por l-atrnúc)2. Rewsáo Bibliagrfua
origina-se
um
bolbo
apartir do
qualo
miçelis
cresse radialmçnte, çolonizandoo
seuinterior
(Bourne& Kerry,
1996; Kerry,200U.
A
entradado fungo no
hospedçiro resultade uma
pressãoÍisiça
ç
tambem de degradaçãogúmica
somo resultadoda
seçreção de enzimaspelo
apressorio(Lopez-Llorça eí
al.,
2000;
Morton et
al,
2W4),
Uma
protease, designadapor
VePl
foi
parcialmente çaraÇterizada e pareçs ser uÍna çnzima çhave oo proçesso de infecção doovo
ds
nemátodç (Sçgerset
al.,
199ó;Kerry,
2m1).
Observaçõesao
miçrsscopio eleçtroniço mostraram que duranteo
processo de infecção, as enzimasprodwidas
pslofungo
Çausama
desintegraçãoda
canradavitçlina do ovo e a
dissoluçãoparcial
da çamada quitinosae
lipídica
(Kerry,
2001).
A
açtividade enzimática aumenta ainda a permsabilidade da çasça do ovo tsrnando-a mais susceptível à çntrada dç toxinas que sçençentrem
ne
ambiente
(Stirling
199U.
Todos
os
estádiosde
desenvolvimento embrionário do nçmátode podem ser parasitados pelo fungo, embsra em svos imafuros, cujas camadas da çasça ainda não estão completamente formadaq a infecção pelo fungoosorra mais
rapidamentçdo quç çm svss
çontendojovens
do primeiro
s
segundo estádio (Boume et a1.,1996; Abrantes eta1.,20A4"
2.7.4.
A
relação triúr,ófrcafuagenemátodeplanta
A
interacção entreos
diferenlçs organismos e,nvolvidosno
pÍoçossode
controlo biologico é cornplexa edifiçil
de quantilflcar, onde os ditbrçntçs çomponçntes depsndem de vánios façtores e se influenciam mutuamente(Ksrry,
2001; Abrantes et a1.,2002).2. Rewsêo Bbliagrffiea -É7 !2 i -t\cmârullc (h{eloidogare spp.} Á $ fr v
\
lnteracçllo nemátode-funguplâníaFig.10: Esquema da inlerac@ frrngo-nemrâtode-planta (adatrâdo de Kerry,
2ml}
Relação
fungolnemátode
Tal
çsmo açonteçe som a oolonização daúosfera,
também a aptidão para paÍa§itar ovos dç NGR varia çom a estirpsou
isolado do fungo.No
entanto, ditbrentes especissdo
género
Melaidogtne
pareÇampossuir
uma
susceptibilidade semçthantç
aoPlanta hos@eira
NIP
-,.r' ,&, Suugo chlamydosparia)a
Â
orsoeptibilidedÊ da plaúah@ira
afesla u uúlrrlaru ds Gtrttsrs d$ u§llüitul€ e ovos produzidosa
O
tarnaúo dâs galhasé
afectado pelaes$cie da planta, variedade e ttmsidade de nemr{tode
a
As vanedades de cada plmta diferem ua tolerância ao elâque de nemátodes{. &
isole&
da P. Chlqrydospria diferê!nrrl
prbgursuidíds çrurlçru
usuúttxb lúaloidogne spp.a A
susceptibilidadedo
nemátodeMefui&tgne .\BFr- âo fimgo
ê
tgualpra
diferentes isolados{.
Os ovos imamros são mas susçsptivçis ao fi:ngo que ovos maduros{.
G
neniáttxlÉs irrtluzerri o furrgo a pr«luzir enzirms.1. Dilerentes eryécies de plartas variam oa
eu0 cap8oidâdc dc suportar a colonizaçãq do fungo nas suas raizcs
*
Os isolados do fungo P- CklamT'dosporia rlifereur rxl $§l e4xcirlarúe rle r»lotiizar a raiz* O
aumcntoda
quanüdadedc
fungoaplieada
ntt solo
nâcr
aumentâ neces-c'riamente a colonieaç{o da raiz.ri.
O fungo P. í:hlo»r_ydossx»ria é nraisaMrüe
esl rdzesiÍfua&s
relativamrte a raízm saudáveis e o estímulovem emparte da plantaa
O mssçimsnto do firngo na rizss&ra é sstimuhdo pçla Wúrcão ds rnaesgd§ môspoloremát*
2
Reusão BbttogÍúcapaÍasitismo
pelo
P. çhlsÍnyhrportu
embora' as infeççõçsdo
ovo pdo
flrngo
variem aom o estádio de desenvolvimentc embrionário (Kerry, 2001; Abraates etal.,?;00fJ.
O
parasitismodo
fungo
nos ovos
é
tambçm
afeçtadopclo
tarnanhodas
galhasformadas"
Em
çulturas
susceptíveisaos
NGR
ocorrem
Gom
freguência
galhas radiçularesde
grandes dimçnsõçs que po§$]em uma grande proporçãode
massas de ovos Íro seuinterior
que não estão em conta€to Gomo
exterior. Umavez
gueo
fungoprolifera
essçnçialmente na zuperficie darara
não erüra em contaçto com grande partedos ovos. AssinL em
gera!
com
elçvadas densidadesde
nemátodesem
plantas susceptíveis, o frrngo é mçnos efiçiçntç que em çulturas çom baixas dçnsidades dçNGR
(Abranteset
al.,
2CIO2;Kerry;
2001).A
redução da população tendea
sermaior
emplantas
menos
susceptíveisao
nemátodeç
çom
uma tbrte
aptidão para
serçm çolonizadaspelo
fungo (Kerry, 2001;
Abrantes
et
al.,
20A2;).O
impacto
do
P.chlarryfurparia
na população dç NGR é ainda maximizado çom tsmpçraturasinfçriorçs
a 30oC umavcz
que acima dçstevalor
o
desenvolvimento embrionáriodo
nemátodç é ac€lerado enquanto quço
fungo tçm um
çrçsçimento mais lonto, oçorrçndo a eçlssão antes do fungo colonizar a massas de ovos (deLeij
et aÍ.,19924;K.rty,
2001).Relação
fungo/planta
Alguns aspectos desta relação forarn
já
úordados
anÍeriormçntç nestetrúalho. No
entanto
e
pertinentereferir
qu€
a
planta
hospedeiratçm um
pap€l fundamçntal
no cstabeleçimentodo fungo
resultando numamaior ou
menor efrçácia na supressão dosNGR.
Esta relação tem çomo pontos principaiso
façtoda
aptidão gus as plantas têm para ser colonizadas pelo fungo variar çom a especie da planta e por outro lado isoladosdiferçntes
do
P.
chlwny&xporia difsrirem
na
sua
capacidadede
çolsnização
da rizosfera. O fungo e em geral mais abundaüe em plantas infeçtadas pelo nemátode que2. Reusâo Bibliogrâfrca
em ptantas saudáveis s
o
aum€nto datas<a de çresçimento dcsteosolrç
aofim
de çerto tempo apos a instatação da çultura. Este estímulo pode rezultar doinício
da cotonizaçãodirwta
das massasdç
ovosou
dss nufiientes libertados pelaplanta
infeçtada (Kerry,2ool)"
A
densidadçdo fungo
em
plantashos@eiras
"fraça§" pode não sçr
aumentada significativamçnte mesmo com o aumento datara
de aplicação de inóculo"AssiÍL
dadaa
importânciada
planta nçsta relação, asesp*ies
e
variçdades çultivadasdçvçm
ser esçolhidas tendo em conta estesaspwtos
dç
forma
a
maximizaro
impastodo
fungo sobre a poputação dç nemátodç§(Kçrry,
2001; Abrantçs et a1.,20$21.Relação aemátode
/planta
A
susceptibilidade da planta afçctao
númçro de nçmátçdss guç invadçm araiz assimÇomo númçro dç fêmsas que se formam e a quantidade dç evos em cada massa dç ovos.
ConsequentementÇo a dimçnsão das galhas também varia ç por sua
vçz
influçnçia a tar<ade parasitismo pelo fungo e o sucçsso na supressão dos NGR.
2.7.5.
Utilização
de P. chlamydosporiaçomo
dcçoutrols
doNGR
O
fungo
P.
chlamydasporia possui potencialidadestre
sombate aosNGR
maso
modo
de rnaximíar
a
suaefiçaçia não
está ainda completamentç esçlarççido. Nosúhimos
anos têm-sç procurado astrategias decontrolo
deNGR am
que se çombina aaplicação
dç
P.
chlarnyfusparia
çom
outros
metodos aomeadamentsa
rotação
de culturas comhospdeiros
çom maior resistência aos nemátodes(Atkins
et a1.,2003). A§ plantasutilizadas
dçverão apressntaruma
boa capacidade para suportaro
fungo
narizosfera mas
seremfracos
hospedeirosdç NGR
formando galhas
p€(1uanas ÇQmoresposta à infecção,