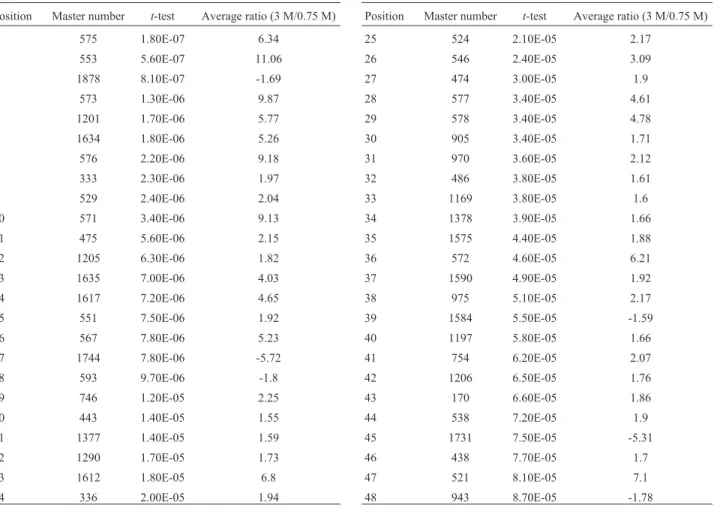
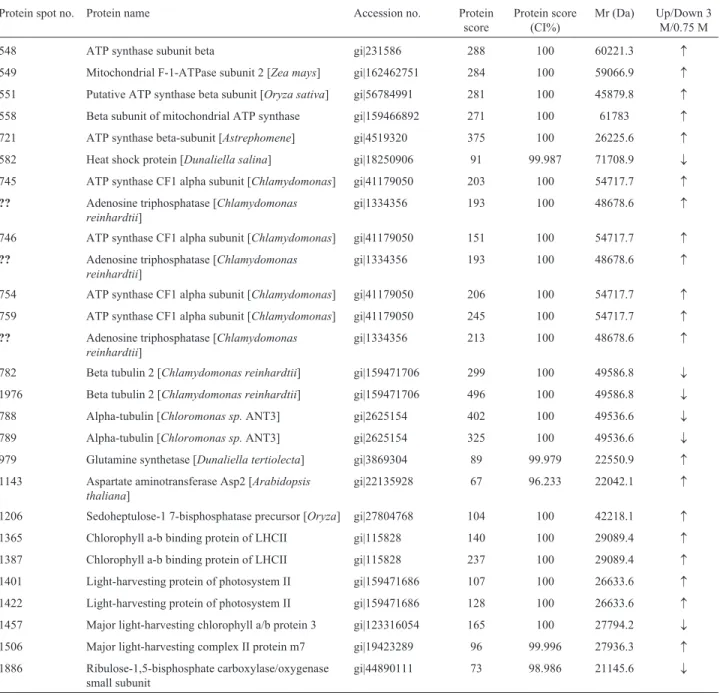
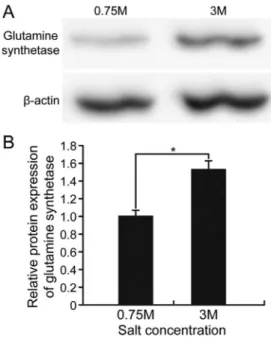
Proteomic analysis of halotolerant proteins under high and low salt stress in Dunaliella salina using two-dimensional differential in-gel electrophoresis
Texto
Imagem

Documentos relacionados
(Nilsson et al ., 2012) The proteomic analysis of plasma from obese and thin rats fed with a high fat diet by Choi and Wang revealed that the change in the levels of twelve
Este relatório relata as vivências experimentadas durante o estágio curricular, realizado na Farmácia S.Miguel, bem como todas as atividades/formações realizadas
a 2.2.1946; foi a mais liberal e democrática; muito emendada e modificada regularmente e por via de atos institucionais; sofreu impactos de crises e investidas golpistas, até que
redemocratização no Brasil, expressada pela Constituição de 1988 (BRASIL, 1988), estabeleceu um novo paradigma para as liberdades individuais, diferenciando-se do período de
1) Differential in gel electrophoresis identifies differences in the proteomic profile produced by salivary and larval proteins of Rhipicephalus microplus ticks fed on
In the present study, a proteomic approach using a label- free quantitative MS analysis identified eight differentially expressed proteins in the serum of DM1 patients compared
A proteomic analysis using Salmonella SraL mutant and SraL overexpressing strains detected the variation of the expression of several proteins involved in
In this study, we undertook a proteomic analysis of liver from the colubrid snake Elaphe taeniura using a combination of two-dimensional electrophoresis (2-DE) and matrix-assisted
