THE MALE GENITAL ACCESSORY GLAND COMPLEX OF THE
CATTLE TICK
Boophilus microplus
(CANESTRINI, 1887)
(ACARI: IXODIDAE)
CASIMIRO GARCIA-FERNANDEZ,1,2 SONIA M. LAUER DE GARCIA2 and ROSANE NUNES GARCIA3
1Instituto de Biociências, Pontifícia Universidade Católica do Rio Grande do Sul, Avenida Ipiranga, 6681,
CEP 90619, Porto Alegre, RS, Brazil
2Instituto de Ciências Básicas da Saúde, Departamento de Ciências Morfológicas, Universidade Federal do Rio
Grande do Sul, Rua Sarmento Leite, 500, CEP 90046-900, Porto Alegre, RS, Brazil
3Instituto de Biociências, CPG em Genética e Biologia Molecular, Universidade Federal do Rio Grande do Sul,
Avenida Bento Gonçalves, 9500, CEP 91501-970, Porto Alegre, RS, Brazil
Correspondence to: Casimiro Garcia-Fernandez, Rua Aneron Correa de Oliveira, 53, Jardim do Salso, CEP 91410-070, Porto Alegre, RS, Brazil
Received May 19, 1997 – Accepted June 05, 1998 – Distributed August 28, 1998
(With 5 figures)
ABSTRACT
A topographical and histological study of the male genital accessory gland complex of Boophilus microplus was undertaken. Ten lobes were found, the most prominent of which is the single
dorso-median lobe, subdivided into antero-dorsal and postero-dorsal lobes. The other lobes are: a pair of postero-ventral lobes, a pair of lateral lobes (subdivided into dorso-lateral and postero-lateral lobes), a pair of antero-ventral lobes, a single medio-ventral lobe and a pair of latero-ventral lobes. These lobes were observed to present individual topographical and histological characteristics, with inde-pendent openings, hence the conclusion is that they form a gland complex. The secretory nature of the dorso-median lobe duct was also analysed.
Key words: male reproductive system, genital accessory glands, ticks, Ixodidae.
RESUMO
O complexo glandular genital acessório masculino do carrapato bovino Boophilus microplus (Canestrini, 1887) (Acari: Ixodidae)
Uma análise topográfica e histológica do complexo glandular sexual acessório do sistema reprodutor masculino de Boophilus microplus foi realizada no presente trabalho. Dez lobos foram encontrados;
o mais proeminente deles é o lobo ímpar dorso-mediano, subdividido em lobos ântero-dorsal e pós-tero-ventral. Os demais lobos são: um par de lobos póstero-ventrais, um par de lobos laterais (subdivi-didos em lobos dorso-laterais e póstero-laterais), um par de lobos ântero-ventrais, um lobo ímpar mé-dio-ventral e um par de lobos látero-ventrais. Foi observado que estes lobos apresentam características topográficas e histológicas individuais, com aberturas independentes, concluindo-se, portanto, tratar-se de um complexo glandular. A natureza secretora do ducto excretor do lobo dorso-mediano também foi analisada.
Palavras-c have: aparelho reprodutor masculino, glândulas genitais acessórias, carrapatos, Ixodidae.
INTRODUCTION
In Arthropoda, the reproductive system usu-ally possesses some kind of secretory structure,
either simple cells of the epithelium that line its ducts, or accessory glands.
forma-tion (Fänger & Naumann, 1993). The secretory production of these glands can be so highly spe-cialized that, in some instances, different sper-matophore zones may come to be secreted by different cells of the accessory glands and/or other portions of the reproductive system, such as the excretory duct (Fänger & Naumann, 1993).
In Ixodidae these glands are at first respon-sible for the formation of the spermatophore (Tatchell, 1962; Chinery, 1965; Oliver, 1991). Leahy & Galun (1972) demonstrated in Ar gas persicus that their secretions stimulate oogenesis
and vitelogenesis. Pappas & Oliver (1972) sug-gested an oocyte-stimulating factor in Dermacen-tor andersoni. In addition, Shepherd et al. (1982)
mentioned that a polypeptide (12.5 Kda molecu-lar weight) from the extracts of male genital acessory glands was able to evert spermatids in vitro experiments. Also, Oliver et al. (1984)
re-port the stimulation of the female reproductive system by male extracts.
The male accessory genital glands of ticks are located immediately posterior to the syngan-glion. They present a complex morphology since there are great variations according to the lobe considered. For ixodid ticks, morphological analyses of these glands were carried out by Russer (1933), Yalvac (1939), Douglas (1943), Till (1961), Chinery (1965) and Mulmule & Thakare (1985). For argasid ticks, the glands were studied by Robinson & Davidson (1914), Wagner-Jevseenko (1958), Roshdy (1961) and El Shoura (1987).
The ixodid Boophilus microplus is an
im-portant vector of diseases in cattle, one which is babesiosis, which affects cattle in Brazil and else-where. Moreover, in attaching to the bovine host, it causes direct losses in the quality of the leather (Morán, 1976).
The present work deals with a topographical and histological study of the male genital acces-sory gland complex of Boophilus microplus. In
the proximal future the ultrastructure of each gland of the complex will be studied in the elec-tron microscopy and also the nature of their se-cretions will be investigated.
MATERIAL AND METHODS
Male specimens of Boophilus microplus
we-re collected on a dairy farm in Viamão, Rio
Grande do Sul, Brazil. Their reproductive system was dissected, drawn and photographed, with the use of a dissecting microscope.
Isolated gland complexes were fixed either in Bouin solution to be included in paraffin or in 4% paraformaldehyde to be included in historesin. The material fixed in Bouin was dehydrated in ascending alcoholic grades and included in paraffin. Five µm sections were obtained and haematoxylin-eosin was used as stain.
The material fixed in 4% neutral buffered pa-raformaldehyde was dehydrated in ethanol ascend-ing grades, pre-infiltrated in a mixture of historesin and absolute ethanol at the same proportion for 2 hours at room temperature. Infiltrations was car-ried out from solution A (A* – historesin 50 ml to 0,5 g activator), in which the material remained for one night. The material was then transferred to plastic molds containing 1 ml of solution B (B* – infiltration solution A – 15 ml, accelerator, 1 ml) and left at room temperature for 2 hours for po-lymerization. Sections of about 3 µm were ob-tained with glass knives in an automatic micro-tome. Haematoxylin-eosin was also used as stain.
RESULTS
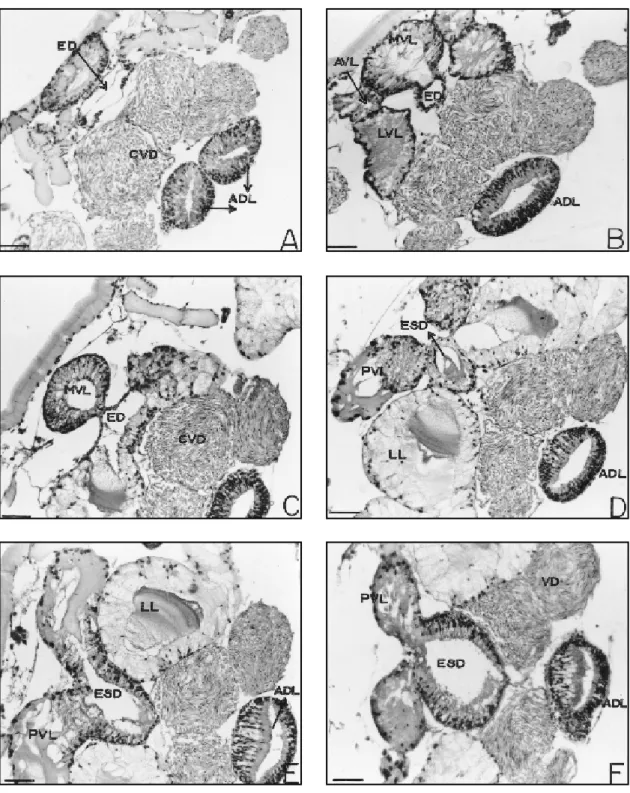
The genital accessory gland complex (Figs. 1, 2, and 3), except for the dorso-median lobe lies postero-ventrally to the synganglion and to the common vas deferens. It is composed of struc-turally distinct lobes. The most prominent lobe is the dorso-median which is subdivided in two regions: the antero-dorsal (ADL) and postero-dorsal (PDL) lobes. From the ventral region, on the border between the anterior and the posterior region, there appears a duct that preserves the same secretory nature of the remaining gland, the excretory-secretory duct (ESD) and discharges into another duct with thinner walls, the ejacu-latory duct (ED) which, in turn, runs to the genital opening. The remaining lobes comprising the accessory gland complex are listed below as they appear in a postero-anterior direction:
● a pair of postero-ventral lobes (PVL),
● a pair of lateral lobes, subdivided in a
pair of dorso-lateral lobes (DLL) and a pair of postero-lateral lobes (PLL);
● a pair of latero-ventral lobes (LVL); ● a single medio-ventral lobe (MVL); ● a pair of antero-ventral lobes (AVL).
The pairs of lateral, latero-ventral and antero-ventral lobes and the single medio-antero-ventral lobe open in the thin-walled duct, ejaculatory duct, which discharges into the genital opening and also receives the opening of the common vas deferens.
Dorso-median lobe
Antero-dorsal lobe
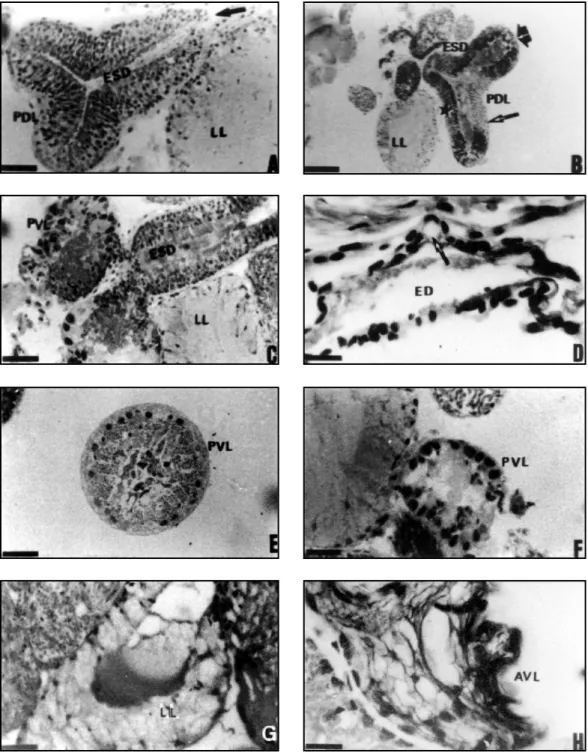
It begins in a cephalic position, almost over-laying the synganglion. In its anterior region it appears bifurcated, forming two lateral sacs that project anteriorly and laterally. These sacs appear to be joined at the median region forming a single chamber, which ultimately constricts to give rise to the postero-dorsal lobe, a markedly larger structure. (Figs 1, 2, 3 – ADL, PDL).
This lobe consists of narrow, columnar se-creting cells which are juxtaposed, have ill-defined limits and nuclei at varying levels. The gland lumen is large and well-defined (Figs. 4 A-F). The secreting cells nuclei are elongated or
rounded, presenting a very compact chromatin. Basal and rounded nuclei are also observed. The secretion present in the cells is made up of little acidophilic granules.
Regionalizations are observed in this lobe, which presents cells completely filled with gran-ules or cells with clear cytoplasm. As transver-sal sections toward the median region of the lobe are examined, cells of the ventral wall become taller, displaying clear cytoplasm and gland se-cretion accumulated in the apical region (Fig. 4E and F – ADL).
Externally, the whole dorso-median lobe is lined with squamous cells, with compact nucleus and eosinophilic cytoplasm.
Postero-dor sal lobe
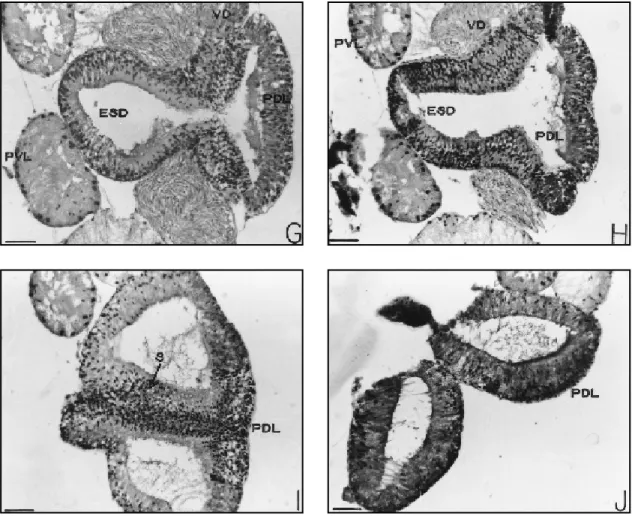
Initially single, it soon displays an external longitudinal constriction which subdivides it caudally into two identical sacs. Internally, the constriction is accompanied by a septum which gives rise to two chambers (Fig. 4G and J – PDL). Ventrally, at a point in the limit between the antero-dorsal and postero-dorsal lobes, there ap-pears the excretory-secretory duct that runs ante-riorly (Figs. 4G and H, 5A and B – ESD), receiv-ing in its most proximal part the discharge from postero-ventral lobes (Fig. 5C – ESD, PVL), and
Figs. 1-3 — Male accessory sex gland complex of Boophilus microplus. 1 — Ventral view; 2 — Dorsal view; 3 — Lateral
further extending as a duct with a smaller diam-eter, namely, the ejaculatory duct (Figs. 5A – arrow and 5D – ED), which, before opening to the outside, receives the discharge from the re-maining lobes, as well as from the common vas deferens (Fig. 5D – arrow).
Though the postero-dorsal lobe presents dif-ferent regionalizations, these are histologically similar. The epithelium that composes this lobe consist of narrow, tall cells resting on the basal lamina, as in the antero-dorsal region of this lobe (Figs. 4G and J – PDL). Basal squamous cells are also observed lining the gland lobe externally. The secreting cells show a variety of secretions, with some cells containing basophilic granules and others showing homogeneous and acidophilic secretion. In the broad and well-defined lumen, the secretion is granular or homogenous and colloidal in aspect.
At least three distinct groups of secretory cells are observed in the most dilated part of the gland wall, opposite and lateral to the appearance of the excretory duct: in the mid portion of the dorsal wall, opposite the excretory-secretory duct, cells are clearer and filled with a finely granular material and slightly larger granules, and the presence of a clear homogeneous secretion is also observed (Fig. 5B – thin arrow); on the lateral walls of this lobe, cells present a fine granular, highly eosinophilic material, as well as dense drops of homogeneous acidophilic secretion (Fig. 5B – broad arrow); though at the end of the lateral walls, reaching the walls that form the ex-cretory-secretory duct, the histological nature is similar to the rest of the gland, the secretion is predominantly granular and not so eosinophilic (Fig. 5B – asterisk).
The wall of the septum that divides inter-nally the postero-dorsal lobe into two chambers is histologically similar to the rest of the lobe and also displays a secretory character (Fig. 4I – S).
Excretory-secretory duct
Initially it has a secretory character, display-ing similar histology to the dorso-median lobe from which it arises (Figs. 3, 4E and H and 5A and C – ESD).
As it approaches the thin-walled duct, the ejaculatory duct, its cells become lower and lower, losing their secretory character (Figs. 4D – ESD and 5A – arrow). In its bordering region
with the ejaculatory duct, it receives the discharge from the pair of postero-ventral lobes (Fig. 5C).
Postero-ventral lobes
They correspond to a pair of elongated glands that run caudally, traversing the postero-dorsal lobe (Figs. 1, 2 and 3 – PVL). They dis-charge into the excretory-secretory duct of the postero-dorsal lobe at its connection with the thin-walled duct (Fig. 5C). These lobes are con-stituted by two histologically distinct zones. Most of them is made up of secretory cuboid or pyra-midal cells, with nuclei located at the lower third with compact chromatin.
They present regular and clearly eosino-philic granular material in the portion above the nucleus through the cell apex. The basal portion of the secreting cell shows a homogeneous and basophilic cytoplasm (Fig. 5E – PVL).
In the gland portion near the opening into the excretory-secretory duct, the secreting cells present a distinct histological aspect. At this level, some of the nuclei of the secretory cells are rela-tively large while others are very small. Both are rounded and have a compact chromatin. The cells of this region display a compact and highly aci-dophilic cytoplasm with indistinguishable cell borders. Sometimes cells in this region are highly vacuolated (Figs. 5C and F – PVL).
Lateral lobes
These consist of a pair of lobes that open before the postero-ventral lobes, reaching the thin-walled duct. Each component of the pair presents its gland body subdivided into two lobes: the dorso-lateral and postero-lateral lobes. Its configuration is Y-shaped.
Steresoscopic microscopy of this lobe re-veals a lighter tone than the remaining lobes. Both dorso-lateral lobes run in caudal direction, dorsally, while the two ventro-lateral lobes run in caudal direction, ventrally (Figs. 1 and 3 – DLL, PLL).
Fig. 4 A-J — Sequence of transversal histological sections through the male accessory gland complex of Boophilus microplus,
Fig. 4 A-J (cont.) — Sequence of transv ersal histological sections through the male accessory gland complex of Boophi-lus micropBoophi-lus, showing the discharge of the different lobes into the excretory-secretory duct (D-H) of postero-ventral lobe
or into the ejaculatory duct (A-C) (thin walls). Haematoxylin-eosin stain. Scale bars = 10 µm. ESD – excretory-secretory duct; PDL – postero-dorsal; PVL – postero-ventral lobes; S – Septum; VD – vas deferens.
Antero-ventral, medio-ventral and latero-ventral lobes
Ventrally, at a proximal position and right below the antero-dorsal lobe of the dorso-median lobe, there are five glands (two paired and one single) surrounding ventrally and laterally the eja-culatory duct: a pair of antero-ventral lobes, a single medio-ventral and a pair of latero-ventral lobes. Owing to their localization, more cephalic and circumscribing the thin-walled duct, we will call this group of lobes as the “necklace” (Figs. 1 and 3 – AVL, MVL, LVL).
Antero-ventral lobes
Histologically these are the most peculiar parts of the “necklace” group due to the
vacu-olated aspect and intense basophilic cytoplasm of their cells.
The nuclei are distributed irregularly and most of them are highly compact. The cell bor-ders are difficult to define. Externally to the secreting cells are squamous cells whose nuclei contain highly compact chromatin. The lumen is poorly defined (Fig. 5H – AVL).
Medio-v entral lobe
This is a single lobe and the most central of the “necklace” group. It is a globose structure, the largest of this group.
filled with secretory granules, nuclei at different levels, compact chromatin and secretion of two types: eosinophilic granules or a non-granular secretion, forming large drops as they fall into the lumen. Externally it appears lined by a squamous epithelium. Its lumen is evident, filled with both granular and colloidal secretion (Fig. 5I – MVL).
Latero-ventral lobes
These correspond to a pair of globose glands. Histologically their secreting cells pos-sess rounded borders, nuclei with compact chro-matin and eosinophilic granular secretion of variable sizes, though zones of dense and homog-enous secretion are also observed.
Large vacuoles filled with scarce granules are noted. the lumen is not clear-cut (Fig. 5J – LVL).
DISCUSSION
The morphology of male genital accessory glands of Boophilus microplus is similar to that
reported for other ixodid ticks (Russer, 1933; Douglas, 1943; Till, 1961; Chinery, 1965; Mulmune & Thakare, 1983). Some dissimilari-ties were observed as listed below:
1. The ventral prolongation of the
median-dorsal lobe is regarded by Chinery (1965) as the collecting duct because along its course it re-ceives the discharge of the other components of the accessory gland in Haemaphysalis spinigera.
this structure in Rhipicephalus appendiculatus is
considered by Till (1961) as the median part of the dorso-median lobe. In Boophilus microplus
it has a clear-cut secretory character and does not receive the discharge from the other lobes, except for the postero-ventral lobes which open in their bordering region with the thin-walled duct. Since in Boophilus microplus the remaining lobes
dis-charge into the thin-walled duct, and as the duct of the dorso-median lobe has secretory charac-teristics, we consider it as an excretory-secretory duct of the dorso-median lobe. On the other hand, the thin-walled duct that receives the other lobes and the discharge of the common vas deferens was the ejaculatory duct.
2. The lobes present in the genital accessory
complex of Boophilus microplus correspond to
those described by Chinery (1965) for Haema-physalis spinigera. Mulmule & Thakare (1985)
showed a correspondence between the gland lobes in Rhipicephalus sanguineus and those described
by Chinery (1965) in Haemaphysalis spinigera.
However, Mulmule and Thakare (1985) reported a lobe, referred to as the dorso-lateral lobe, that was observed neither in Haemaphysalis spinigera
nor in Boophilus microplus. The structure referred
to by Mulmule and Thakare does not correspond to the dorso-lateral lobe reported for Haemaphysa-lis spinigera and Boophilus microplus. It
corre-sponds to one of the lateral lobes in these two species. Even more, Mulmule and Thakare desig-nated as postero-lateral the corresponding lateral lobes in Haemaphysalis spinigera and Boophilus microplus. What is called by Chinery (1965) in Haemaphysalis spinigera and in the present work
in Boophilus microplus as postero-lateral lobe
corresponds to a region of the lateral lobes, once that in Haemaphysalis spinigera and in Booph ilus microplus, the lateral lobes are subdivided in two
regions, the dorso-lateral an postero-lateral lobes. The histological analysis of the dorso-median lobe in Boophilus microplus shows a variety of
se-cretions, whether among regions of the lobe or among cells belonging to the same region. This characteristic was observed in Haemaphysalis spiniger a by Chinery (1965) when the presence of
secretory granules of varying sizes was reported. Studying Rhipicephalus sanguineus, Mulmule &
Thakare (1985) regionalized the secretory activ-ity of the median lobe in the postero-dorsal por-tion in five areas which they called categories A, B. C, D and E. In Boophilus microplus, the
pres-ence of regionalization in the postero-dorsal portion of the median-dorsal lobe is evident, and at least three regions were established in the present study.
Comparison between gland lobes of Boo-philus microplus with specimens from the argasid
group is difficult because there appears to be no complete correspondence among them. However, the dorso-median lobe is constant in all cases reported in the literature and is always the most prominent one.
Robinson & Davidson (1914) have histo-logically classified the male genital accessory gland of Argas persicus in two great groups:
Granular lobes are described as constituted by cylindrical cells filled with irregular granules. Later, Russer (1933) and Douglas (1943) described spongy and granular tissue in genital accessory glands of Hyalomma aegyptium and Dermacentor andersoni, respectively. However, Till (1961) and
Chinery (1965), in studying Rhipicephalus appen -diculatus and Haemaphysalis spinig era,
respec-tively, considered that all lobes are granular, since they do not display this granular aspect until short after the tick has begun to feed.
On the other hand, Chinery (1965) con-cluded that the reticular aspect of the dorso-lateral and postero-lateral lobes, in the absence of gran-ules, would bear some resemblance with the “spongy” tissue in Argas persicus. Mulmule &
Thakare (1985), studying the genital accessory glands in Rhipicephalus sanguineus, largely agree
with Till (1961) and Chinery (1965).
In our observation in Boophilus microplus
it became clear that some lobes, namely, the pair of lateral lobes, the antero-ventral pair and the latero-ventral pair, do show a “spongy” aspect due to the presence of a great number of cell vacuoles.
All the remaining lobes are predominantly granular. We have considered these as histologi-cal characteristics of the gland lobes, since we have worked with adult specimens. It was also mentioned and justified by El Shoura (1987) that the appearance of granular or spongy lobes in the accessory genital glands of Ornithodorus (pavloskyella) erraticus does not represent any
specific phase of secretory activity because the secretory lobes had always the same appearance.
Acknowledgements — The authors are indebted to biologist
Eliane de Oliveira Borges, technician of the Laboratory of Histology and Embriology of the Morphological Sciences Department of the Universidade Federal do Rio Grande do Sul, for her collaboration in the preparation of histological sections in paraffin. This work was supported by Propesp/ UFRGS and CNPq/UFRGS.
REFERENCES
CHINERY, W. A., 1965, Studies on the various glands of the tick Haemaphysalis spinigera Neumann, 1897. Acta Trop., 22(3): 235-266.
DOUGLAS, J. R., 1943, The internal anatomy of Dermacen-tor andersoni Stiles. Univ. Calif. Publ. Entomol., 7:
207-272.
EL SHOURA, S. M., 1987, Fine structure of the vase def-erentia, seminal vesicle, ejaculatory duct, and accessory glands of male Ornithodoros (Pavlo vskyella) erraticus
(Acari: Ixodoidea: Argasidae). J.Med. Entomol., 24(2):
235-242.
FÄNGER, H. & NAUMANN, M.,1993, Correlation between the mesodermal male genital ducts and the spermato-phore structure in a ditrysian moth, Zygaena trifolii
(Esper, 1783) (Insecta, Lepidoptera, Zyg aenidae). Acta Zool., 74(3): 239-246.
LEAHY, M. G. & GALUN, R., 1972, Effect of mating on oogenesis and oviposition in the tick, Argas per sicus
(Oken). Par asitology, 65: 167-178.
MORÁN, C. G. R., 1976, Investigaciones realizadas en el campo de la parasitología veterinaria. Ministerio del Desarollo Agropecuario, Universidad de Panama, 56. MULMULE, S. & THAKARE, V. K., 1985, Cytology and
cytochemistry of male acessory gland in the dog tick,
Rhipicephalus sanguineus (L.) (Acarina: Ixodidae). Z.
Mikrosk. Anat. For sch., 99(1): 25-36.
OLIVER JR., J. H., 1991, Tick reproduction: sperm devel-opment and cytogenetics. In: F. Obenchain & R. Galun
(eds.), Physiolo gy of Ticks, 245-275. Perg amon Press,
New York.
OLIVER JR., J. H.; POUND, J. M. & ROSS, H. A., 1984, Induction of e gg maturation and oviposition in the tick
Ornithodoros parkeri (Acari: Argasidae) J. Parasit., 70(3): 337-342.
PAPPAS, P. J. & OLIVER JR., J. H., 1972, Reproduction in ticks (Acari: Ixodoidea). 2. Analysis of the stimulus for rapid and complete feeding of female Dermacentor variabilis (Say.). J. Med. Entomol., 9:47-50. ROBINSON, L. E. & DAVIDSON, J., 1914, The anatomy of
Argas per sicus Ok en. Parasitology, 6: 342-424. ROSHDY, M. A., 1961, Comparative internal morphology
of subgenera of Argas ticks (Ixodoidea, Arg asidae). 1.
Subgenus Carios: Argasvespertilionis (Latreille, 1802). J.Parasit., 47: 987-994.
RUSSER, M., 1933, Beiträge zur kenntnis des Chitins und der Muskulatur der Zeck en (Ixodidae). Z. Morph. Oek. Tiere, 27: 199.
SHEPHERD, J. G.; OLIVER, J. H & HALL, J. D., 1982, A polypeptide from male accessory glands which triggers maturation of tick spermatozoa. Int. J. Invertebrate Reprod. and Develop., 5: 129-137.
TATCHELL, R. I., 1962, Studies on the male acessory re-productive gland and the spermatophore of the tick Argas persicus (Oken.). Par asitology, 52: 133-142.
TILL, W. M., 1961, A contribution to the anatomy and his-tology of the brown tick, Rhipicephalus appendiculatus
Neumann. Mem. Ent. Soc. Southern Africa, 6: 1-24. WAGNER-JEVSEENKO, O., 1958, Fortpflanzung bei
Ornithodorus moubata und genitale Ubertragung von
Borrelia duttoni. Acta Trop., 15: 118-168.