INTRODUCTION
The action of low-level laser therapy (LLLT) seems to be related to biostimulatory events (1). It is believed to accelerate wound healing capacity by increasing the synthesis of collagen and other proteins as well as the proliferation of cells involved in these events. Decrease of inflammatory response and analgesia are other effects of low-power lasers (1,2). LLLT has also been shown to have a significant bactericidal potential without causing damage to the oral tissues (3). This effect has been documented both in vitro, on diverse microbial species, and in vivo, in the treatment of gingivitis, periodontitis and other oral diseases (3-6).
In Vitro
Effect of Low-Level Laser Therapy on
Typical Oral Microbial Biofilms
Fernanda G. BASSO1
Camila F. OLIVEIRA2
Amanda FONTANA2
Cristina KURACHI3
Vanderlei S. BAGNATO3
Denise M. P. SPOLIDÓRIO2
Josimeri HEBLING2
Carlos A. DE SOUZA COSTA2
1Piracicaba Dental School, UNICAMP - University of Campinas, Piracicaba, SP, Brazil 2Araraquara Dental School, UNESP - Univ. Estadual Paulista, Araraquara, SP, Brazil 3Physics Institute of São Carlos, USP - University of São Paulo, São Carlos, SP, Brazil
The aim of this study was to evaluate the effect of specific parameters of low-level laser therapy (LLLT) on biofilms formed by
Streptococcus mutans, Candida albicans or an association of both species. Single and dual-species biofilms - SSB and DSB - were exposed to laser doses of 5,10 or 20 J/cm2 from a near infrared InGaAsP diode laser prototype (LASERTable; 780 ± 3 nm, 0.04 W).
After irradiation, the analysis of biobilm viability (MTT assay), biofilm growth (cfu/mL) and cell morphology (SEM) showed that LLLT reduced cell viability as well as the growth of biofilms. The response of S. mutans (SSB) to irradiation was similar for all laser doses and the biofilm growth was dose dependent. However, when associated with C. albicans (DSB), S. mutans was resistant to LLLT. For
C. albicans, the association with S. mutans (DSB) caused a significant decrease in biofilm growth in a dose-dependent fashion. The morphology of the microorganisms in the SSB was not altered by LLLT, while the association of microbial species (DSB) promoted a reduction in the formation of C. albicans hyphae. LLLT had an inhibitory effect on the microorganisms, and this capacity can be altered according to the interactions between different microbial species.
Key Words: Biofilm, Candida albicans, low level laser therapy, Streptococcus mutans.
It has been shown that LLLT associated or not with a photosensitizer can cause destruction of oral bacterial and fungal species (7). In these cases, the antibacterial mechanism of action of LLL seems to be related to thermal effects and photo-disruption (7). However, LLLT may not cause immediate cell death, but it causes sublethal damages, such as destruction of the bacterial cell wall and accumulation of denatured protein in the bacterial cytoplasm (8). The integrity of the bacterial cell wall is closely related to the stability of gram-positive bacteria, such as Streptococcus mutans. Thus, damage to the bacterial cell wall may lead to growth inhibition and consequent cell lysis (9). Cytoplasmic accumulation of denatured proteins may induce cellular stress in an
attempt to maintain homeostasis, causing the death of the microorganism (8).
LLLT may also cause disruption of microorganisms by altering the interaction among themselves and with the substrate (10). However, the literature shows that the effects of irradiation on bacterial biofilms seem to be species-dependent (6).
Although oral microorganisms can coexist in an independent manner, they frequently interact, forming communities, which have been referred to as biofilms (11). The microcolonies that form the biofilm may be composed of populations of one or more microbial species, depending on the environmental conditions in which they are formed, such as properties of the surface to which they adhere, presence of nutrients, pH and oxygen availability (12). In some cases, the interaction of different microbial species, such as bacteria and fungi, in biofilm formation, may promote relationships of cooperation among species, such as commensalism and mutualism, increasing their chances of survival and growth (11). The association between S. mutans and Candida albicans is one of the most frequently studied interactions of microbial species (12).S. mutans is one of the main bacterial strains colonizing the oral cavity (13). C. albicans is a commensal yeast of the oral cavity, which, under specific conditions like immunosupression, may become pathogenic, causing candidiasis and affecting life quality by increasing patient morbity (13). It has been described that the interaction between microorganisms of different species may alter their behavior and growth (14). This study evaluated the effect of LLLT with specific parameters on single-species biofilms (SSB) of S. mutans and C. albicans and dual-species biofilms (DSB) associating both microorganisms.
MATERIAL AND METHODS
Microorganisms
Reference strains of S. mutans (UR 158) and C. albicans (ATCC 90028) were used for the formation of SSB and DSB.
The microorganisms were grown in brain heart infusion broth (BHI; Difco Laboratories, Detroit, MI, USA) at 37ºC during 18 h according to their physiological demands. These cultures were centrifuged at 1,500 xg for 10 min and washed with sterile phosphate buffer saline (PBS - pH 7.4). The optical density (O.D.) of these
suspensions were determined at 660 nm (Eppendorf AG 22331; Eppendorf, Hamburg, Germany) and adjusted to values of 0.15 and 1.0 for S. mutans and C. albicans, respectively, which correspond to a stock suspension of 107 colony-forming units per milliliter (cfu/mL).
Microbial Biofilms
Biofilm growth on cell culture plates was done as described by Thein et al. (15). For SSB formation, 1 mL of the suspension was transferred to each well of a culture plate. For DSB formation, equal volumes of suspension of each monoculture were mixed to reach a final volume of 1 mL and were transferred to the wells of a culture plate (1 mL/well). Suspensions of each microbial strain served as controls. The plates were incubated at 37ºC during 1 h and 30 min in an orbital agitator at 75 rpm to promote adhesion of the microorganisms to the surfaces of the wells. After the adhesion period, the cell suspension was aspirated and each well was washed twice with sterile PBS for removal of non-adhered microorganisms. Then, 1 mL of BHI was added to the each well to promote biofilm growth at 37ºC during 48 h. After this period, irradiation of the experimental groups was done. The irradiated and non-irradiated biofilms were analyzed 15 h after incubation. The experiment was performed in triplicate at different moments.
Irradiation
The LLL source used in this study was a near infrared indium gallium arsenide phosphide (InGaAsP) diode laser prototype (LASERTable; 780 ± 3 nm wavelength, 0.4 W power output). This device has been specifically designed to provide a uniform irradiation of the wells of culture plates in which cells or microorganisms are seeded, according to the parameters established for each study. The radiation originated from the LASERTable was delivered on the base of each 24-well plate at doses of 5,10 and 20 J/cm2 with irradiation times of 250, 500 and 1000 s, respectively. The laser doses were established according to Nussbaum et al. (4), who evaluated the bactericidal capacity of LLLT with different wavelengths.
(cfu/mL) and analysis cell morphology under scanning electron microscopy (SEM). The microorganisms assigned to the control groups received the same treatment as that of the experimental groups. The 24-well plates containing the control microorganisms were maintained in the LASERTable for the same irradiation times used in the respective experimental groups, though without activating the laser source (sham irradiation).
Analysis Biofilm Viability (MTT Assay)
Previous studies have already determined microorganisms viability by MTT assay (16,17). As biofilms are capable of aerobic respiration which is held by cytosolic enzymes (18) the MTT assay was carried out to evaluate the viability of the biofilms after irradiation
as described by Kwieciński et al. (16).
For this method, twelve specimens of each experimental and control group were used for analysis of cell metabolism 15 h after the active or sham irradiation. Cell metabolic activity was evaluated by SDH production, which is a measure of the respiratory metabolism of the cells. Each well containing the
biofilms received 300 μL of MTT solution (5 mg MTT
salt in 1 mL PBS). The plates were incubated at 37oC for 2 h. Thereafter, the MTT solution was carefully
aspirated from each well and replaced by 300 μL of
acidified isopropanol solution (0.04 N HCl) to dissolve the violet formazan crystals resulting from the cleavage of the MTT salt ring by the SDH enzyme produced by cellular aerobic respiration of viable microorganisms. After agitation and confirmation of the homogeneity of
the solutions, 3 aliquots de 100 μL of each well were
transferred to a 96-well plate(Costar Corp., Cambridge, MA, USA). The respiratory metabolism was evaluated by spectrophotometry as being proportional to the absorbance measured at 570 nm wavelength with an ELISA plate reader (Multiskan Ascent 354; Labsystems CE, Lês Ulis, France). The values obtained from the 3 aliquots were averaged to provide a single value. The absorbance was expressed in numeric values, which were subjected to statistical analysis to determine the respiratory activity on biofilms.
Biofilm Quantification
Quantification of biofilm was done to evaluate the effect of LLLT on the growth of the microorganisms. Fifteen hours after irradiation, the biofilms were
washed twice with sterile PBS to remove non-adhered microorganisms. Then, 1 mL of PBS was added to each well and the adhered biofilm was carefully scraped (15). The whole procedure was performed by a single operator following a standardized protocol. The suspension containing the microorganisms was aspirated and transferred to a tube, which was vortexed during 1 min. Serial dilutions of the final suspension (10-1 to 10-4) were prepared and inoculated in sucrose bacitracin agar (SB20) and Sabouraud dextrose agar (SDA) for growth of S. mutans and C. albicans, respectively. The plates were incubated at 37°C during 48 h, and the biofilm counts were expressed as cfu/mL.
Analysis of Cell Morphology by SEM
Two specimens of each experimental and control group were used for analysis of the morphology and number of microorganisms in the biofilms 15 h after the active or sham irradiation. Sterile 13-mm-diameter cover glasses (Fisher Scientific) were placed on the bottom of the wells of all groups immediately before seeding of the microorganisms. After exposure to the experimental conditions, the culture medium was removed and the viable cells that remained adhered to the glass substrate were fixed in 1 mL of buffered 2.5% glutaraldehyde solution during 2 h. Next, the cells were submitted to three 5-min rinses with 1 mL
PBS, post-fixed in 200 μL of 1% osmium tetroxide
during 60 min. The microorganisms adhered to the glass substrate were then dehydrated in a series of increasing ethanol concentrations [30%, 50%, 70%, 95% (twice) and 100% (twice) - 30 min in each solution], immersed in 1,1,1,3,3,3-hexamethyldisilazane (HMDS; Acros Organics, Springfield, NJ, USA) for 90 min, and stored in a desiccator during 12 h. The cover glasses were then mounted on metallic stubs, sputter-coated with gold and the morphology of the surface-adhered microbial cells was examined with a scanning electron microscope (JEOL-JMS-T33A Scanning Microscope; USA Inc., Peabody, MA, USA).
Statistical Analysis
RESULTS
Analysis of Biofilm Respiratory Activity (MTT Assay)
SDH production data (MTT assay) according to type of biofilm and laser dose are presented in Table 1. For all laser doses, the lowest SDH production was observed for S. mutans (SSB) (p<0.05) followed by C. albicans (SSB) and DSB. No significant difference (p>0.05) was found between C. albicans (SSB) and DSB.
LLLT did not have significant influence on SDH production by C. albicans (SSB) and DSB, regardless of
the laser dose (p>0.05). However, irradiation of S. mutans (SSB) caused significant reduction of SDH production (p<0.05) at all laser doses, without significant difference among them (p>0.05). In order words, regardless of the dose, LLLT influenced the respiratory activity of S. mutans (SSB).
Biofilm Quantification
Quantification of biofilm (cfu/mL), according to the type of biofilm and laser dose (J/cm2) is presented in Table 2. There was a significant difference (p<0.05) in the reduction in the growth of S. mutans (SSB) between the control and irradiated groups; in the groups subjected LLLT, the reduction was more accentuated and occurred in a dose-dependent fashion.
Regarding DSB, the 5 J/cm2 laser dose promoted a significant reduction in the growth of S. mutans, and this reduction was only slightly intensified when higher laser doses (10 and 20 J/cm2) were used. In association with C. albicans in a DSB configuration, S. mutans exhibited a more intense growth and greater resistance to LLLT compared to the yeast.
Table 1. SDH enzyme production (MTT assay) according to the type of biofilm and laser dose (J/cm2).
Irradiation dose (J/cm2)
Microorganism
S. mutans C. albicans S. mutans + C.
albicans
0 (control) 1.31 (1.27-1.32) A b 2.44 (2.37-2.59) A a 2.47 (2.29-2.55) A a
5 1.01 (0.93-1.08) B b 2.50 (2.20-2.68) A a 2.38 (2.02-2.69) A a
10 1.02 (0.75-1.27) B b 2.47 (2.07-2.65) A a 2.65 (1.91-2.69) A a
20 0.96 (0.81-1.12) B b 2.43 (2.17-2.73) A a 2.49 (1.81-2.66) A a
Values expressed as medians of SDH production (P25-P75) (n=12) (optical density). Same uppercase letters in columns and lowercase letters in rows indicate no statistically significant difference (Mann-Whitney, p>0.05).
Table 2. Quantification of biofilm (cfu/mL) according to the type of biofilm and laser dose (J/cm2).
Irradiation dose (J/cm2)
Biofilm (cfu x107)
S. mutans SSB C. albicans SSB DSB (S. mutans) DSB (C. albicans)
0 (control) 0.20(0.19-0.20) A c 0.58 (0.47-0.65) A a 0.41 (0.39-0.44) A b 0.40 (0.34-0.44) A b
5 0.09 (0.08-0.09) B c 0.28 (0.28-0.29) C b 0.35 (0.32-0.38) B a 0.34 (0.20-0.38) B ab
10 0.07 (0.05-0.07) C c 0.23 (0.20-0.31) C b 0.40 (0.36-0.43) AB a 0.21 (0.15-0.27) C b
20 0.02 (0-0.03) D c 0.39 (0.31-0.48) B a 0.39 (0.30-0.46) AB a 0.15 (0.14-0.15) D b
Values expressed as medians (P25-P75) (n=9) of the number of cfu x107. Same uppercase letters in columns and lowercase letters in
Irradiation of C. albicans (SSB) with doses of 5 and 10 J/cm2 resulted in a statistically significant reduction of growth compared to the non-irradiated control. Increase of laser dose to 20 J/cm2 produced only a slight intensification in biofilm growth inhibition compared to the lower doses, though significant when compared to the control group.
In association with S. mutans in a DSB configuration, C. albicans presented a statistically significant growth inhibition in the irradiated groups compared to control group, and this reduction was dose-dependent.
Comparing the behavior of the 3 types of biofilms,
it can be observed that when the microorganisms were associated in a DSB configuration, C. albicans was more susceptible to LLLT at the highest laser dose (20 J/cm2).
Biofilm Morphology
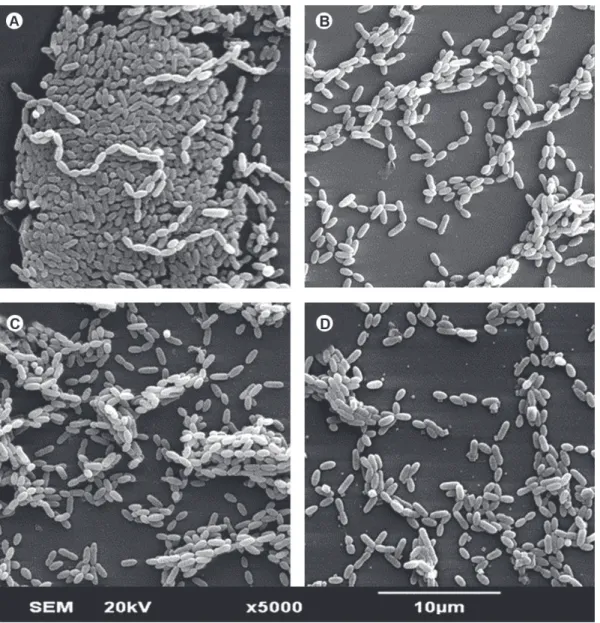
Figure 1 illustrates the effects of LLLT on S. mutans (SSB). The irradiation promoted a decrease in the number of microorganisms, though without significant difference among the laser doses or the types of biofilm. Regarding the morphological alterations, there were no difference between the control group (Fig. 1A) and irradiated groups (Fig. 1 B-D).
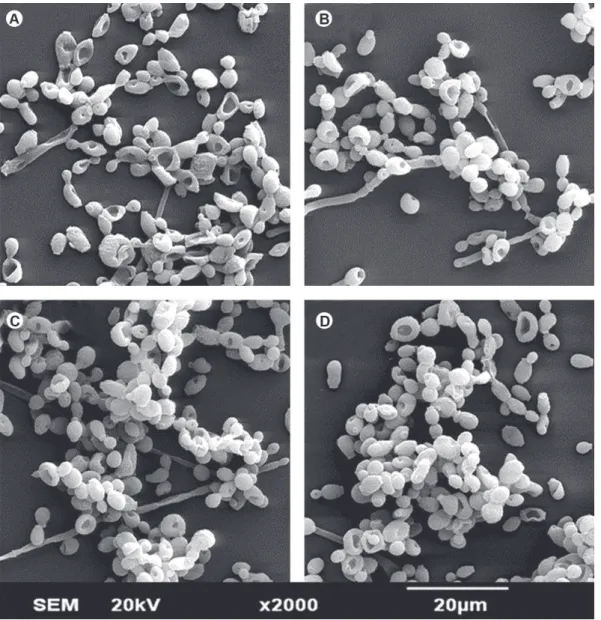
Figure 2. SEM micrograph showing the morphology and structure of SSB of C. albicans (Original magnification ×2000). A= Control, B-D= Biofilm after irradiation with laser dose of 5, 10 and 20 J/cm2.
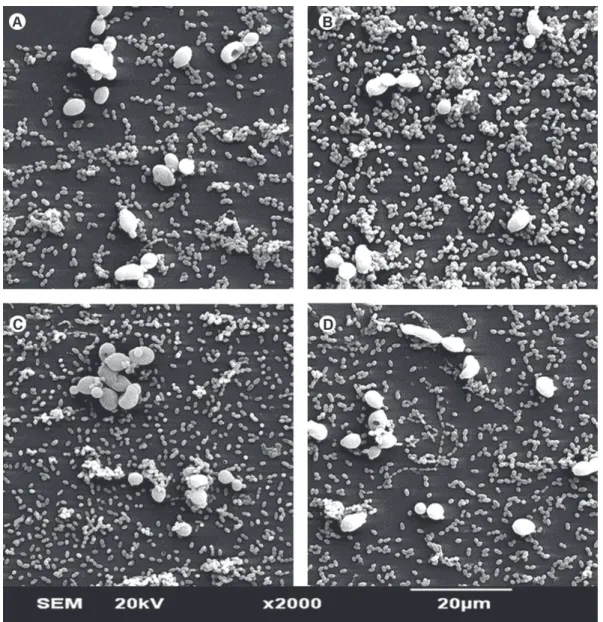
Figures 2 and 3 illustrate the effects of LLLT on the morphology and structure of singles-species biofilms of C. albicans and DSB, respectively. Regardless of laser dose, irradiation of C. albicans (SSB) did not cause morphological alterations in the yeasts nor affected the number of microorganisms that remained adhered to glass substrate (Fig. 2). On the other hand, Figure 3 clearly shows that the association of microbial species caused a reduction in the formation of C. albicans hyphae.
DISCUSSION
Little research has been done on the formation
Figure 3. SEM micrograph showing the morphology and structure of DSB (S. mutans plus and C. albicans) (Original magnification ×2000). A= Control, B-D= Biofilm after irradiation with laser dose of 5, 10 and 20 J/cm2.
of microorganisms in the biofilms, as demonstrated by biofilm growth (cfu/mL) and SEM analysis. The decrease in the number of biofilm microorganisms was similar for all laser doses (5, 10 and 20 J/cm2), indicating that the effect of laser on S. mutans was not dose-dependent.
Nussbaum et al. (4) also demonstrated that LLLT (λ=810
nm) at 5, 10, 20 and 50 J/cm2 doses was effective in inhibiting growth of Pseudomonas aeruginosa and Staphylococcus aureus. However, differently from our findings for S. mutans, those authors found dose- and species-dependent results (4,5). This variation in the responses to the treatment could be attributed to the fact that the different microbial species used in the studies
cfu counts were not observed in the same intensity for this microorganism.
Regarding the morphology of irradiated microorganisms, there were no morphological alterations in the microorganisms after LLLT. However, the analysis of SEM micrographs showed a decrease in the number of S. mutans adhered to the glass substrate in the irradiated groups compared to the group control. This result suggests that LLLT might have altered biofilm organization, leading to the loss of microbial adherence to the substrate, as demonstrated by Krespi et al. (10). Those authors observed a disaggregation of microorganisms after irradiation of S. aureus biofilms with a LLL emitting at 810 nm. Although there is no evidence of induction of cell death, irradiation in the parameters used in the present study seems to have promoted disorganization and disaggregation of the microorganisms in the biofilm, inhibiting their growth and metabolism.
The association of the microbial species in a DSB configuration seemed to have had a protective effect for S. mutans, since no reduction in SDH production or in the number and growth of this microorganism was observed. The interaction between C. albicans and S. mutans biofilms has been characterized as a mutualistic relationship, which is beneficial for both species (21). The yeast may offer a physical scaffold for the proliferation of S. mutans, thus increasing numbers of this bacterium in the biofilm (21). It has also been demonstrated an increase in the growth of C. albicans when associated with S. mutans. This finding confirms not only mutualism, but also coaggregation, which increases the adhesion of both microbial species to the biofilm (12).
In the present study, LLLT was not effective in reducing SDH production in C. albicansi (SSB). However, irradiation with a dose of 5 J/cm2 inhibited the growth of this microorganism. In the same way as for the S. mutans (SSB), this result indicates the laser may cause early or later effects on this yeast. The reduction of cell respiration may be one of the later effects caused by LLLT (8).
The irradiation of DSB of C. albicans and S. mutans inhibited the growth (cfu/mL) of the yeast without reducing its SDH production. Regarding the morphology, the interaction of the microbial species promoted a reduction in the formation of C. albicans hyphae. This effect has also been recently described by Jarosz et al. (20), who suggested an inhibitory effect of S.
mutans over C. albicans by the release of acid molecules in the environment. The presence of these molecules in the environment seems to inhibit the formation and maturation of C. albicans. Some authors investigating the interaction between these microorganisms in biofilm formation have suggested that C. albicans inhibition in the presence of S. mutans in multi-species biofilms is due to competition for adhesion niches and nutrition, thus decreasing the adhesion of these microorganisms when associated (11).
Under the conditions tested in the present study, LLLT had an inhibitory effect on typical oral microbial biofilms, and this capacity can be altered according to the interactions between different species. Further research is needed to evaluate the behavior of different microorganisms typically found in the oral cavity, and their interaction in biofilms when subjected or not to LLLT.
RESUMO
O objetivo deste estudo foi avaliar o efeito de parâmetros específicos de irradiação com laser de baixa intensidade sobre biofilmes formados por Streptococcus mutans (S. mutans),
Candida albicans (C. albicans) ou associação de ambas as espécies. Biofilmes isolados ou associados destes microrganismos foram irradiados com um dispositivo laser infra-vermelho próximo de diodos InGaAsP (LaserTABLE 780 ±3 nm, 0,04W), utilizando-se para isto o dispositivo LASERTable. Quinze horas após a irradiação, foi demonstrado, por meio da avaliação da viabilidade celular (Teste de MTT), da morfologia das células (MEV) e do crescimento do biofilme (UFC/mL), que esta terapia foi capaz de reduzir o metabolismo celular, número de microrganismos presentes no biofilme, bem como seu crescimento no local. Quanto à viabilidade celular, a resposta à irradiação do biofilme de S. mutans (SSB) foi semelhante para todas as doses de energia, sendo que o crescimento do biofilme foi dose dependente. Porém, quando associado à C. albicans, este microrganismo apresentou resistência à fototerapia. Já a C. albicans associada ao S. mutans apresentou redução de crescimento significativa, sendo este resultado também foi dose dependente. A morfologia dos microrganismos não foi alterada pelas irradiações realizadas quando em biofilmes isolados. A associação entre os microrganismos promoveu redução na formação de hifas pela C. albicans. A laserterapia de baixa intensidade apresentou efeito inibitório sobre microrganismos, sendo que esta capacidade pode ser alterada de acordo com a interação entre diferentes microrganismos.
ACKNOWLEDGEMENTS
REFERENCES
1. Marques MM, Pereira AN, Fujihara NA, Nogueira FN, Eduardo, CP. Effect of low-power laser irradiation on protein synthesis and ultrastructure of human gingival fibroblasts. Lasers Surg Med 2004;34:260-265.
2. Damante CA, De Micheli G, Miyagi SPH, Feist IS, Marques MM. Effect of laser phototherapy on the release of fibroblast growth factors by human gingival fibroblasts. Lasers Med Sci 2009;24:885-891.
3. Moritz A, Schoop U, Goharkhay K, Schauer P, Doertbudak O, Wernisch J, et al.. Treatment of periodontal pockets with a diode laser. Lasers Surg Med 1998;22:302-311.
4. Nussbaum EL, Lilge L, Mazzulli T. Effects of 630-, 660-, 810-, and 905-nm laser irradiation delivering radiant exposure of 1-50 J/cm2 on three species of bacteria in vitro. J Clin Laser Med Surg
2002;20:325-333.
5. Nussbaum EL, Lilge L, Mazzulli T. Effects of low-level laser therapy (LLLT) of 810 nm upon in vitro growth of bacteria: relevance of irradiance and radiant exposure. J Clin Laser Med Surg 2003;21:283-290.
6. Lino MDMC, Carvalho FB, Oliveira LR, Magalhães EB, Pinheiro ALB, Ramalho LMP. Laser phototherapy as a treatment for radiotherapy-induced oral mucositis. Braz Dent J 2011;22:162-165.
7. Maver-Biscanin M, Mravak-Stipetic M, Jerolimov V, Biscanin A. Fungicidal effect of diode laser irradiation in patients with denture stomatitis. Lasers Surg Med 2004;35:259-262.
8. Dworkin M. Endogenous photosensitization in a carotinoidless mutant of Rhodopseudomonas speroides. J Gen Physiol 1958;43:1099-1112.
9. Rosenberg B, Kemeny G, Switzer RC, Hamilton TC. Quantitative evidence for protein denaturation as the cause of thermal death. Nature 1971;232:471-473.
10. Krespi YP, Kizhner V, Nistico L, Hall-Stoodley L, Stoodley P. Laser disruption and killing of methicillin-resistant Staphylococcus aureus biofilms. Am J Otolaryngol 2011;32:198-202.
11. Shirtliff ME, Peters BM, Jabra-Rizk MA. Cross-kingdom interactions: Candida albicans and bacteria. FEMS Microbiol Lett 2009;299:1-8.
12. Pereira-Cenci T, Deng DM, Kraneveld EA, Manders EMM, Del Bel Cury AA, ten Cate JM, et al.. The effect of Streptococcus
mutans and Candida glabrata on Candida albicans biofilms formed
on different surfaces. Arch Oral Biol 2008;53:755-764.
13. Marsh PD. Microbial ecology of dental plaque and its significance
in health and disease. Adv Dent Res 1994;8:263-271.
14. Karkowska-Kuleta J, Rapala-Kozik M, Kozik A. Fungi pathogenic to humans: molecular bases of virulence of Candida albicans, Cryptococcus neoformans and Aspergillus fumigatus. Acta Biochim Pol 2009;56:211-224.
15. Thein ZM, Samaranayake YH, Samaranayake LP. Dietary sugars, serum and the biocide chlorhexidine digluconate modify the population and structural dynamics of mixed Candida albicans and
Escherichia coli biofilms. APMIS 2007;115:1241-1251. 16. Kwieciński J, Eick S, Wójcik K. Effects of tea tre (Melaleuca
alternifolia) oil on Staphylococcus aureus in biofilms and stationary phase. Int J Antimicrob Agents 2009;33:343-347. 17. Wang ZC, Fan LY, Jiang JQ, Cai W, Ding Y. Study on the
counting of Streptococcus mutans, Streptococcus sanguis, Haemophilus actinomycetemcomitans by methyl thiazolyl tetrazolium colorimetric method. Hua Xi Kou Qiang Yi Xue Za Zhi 2010;28:306-310.
18. Nguyen PTM, Abranches J, Phan T, Marquis RE. Repressed respiration of oral Streptococci grow in biofilms. Curr Microbiol 2002;44:262-266.
19. Singleton S, Treloar R, Warren P, Watson GK, Hodgson R, Allison C. Methods for microscopic characterization of oral biofilms: analysis of colonization, microstructure, and molecular transport phenomena. Adv Dent Res 1997;11:133-149.
20. Jarosz LM, Deng DM, van der Mei HC, Crielaard W, Krom BP. Streptococcus mutans competence-stimulating peptide inhibits Candida albicans hypha formation. Eukaryot Cell 2009;8:1658-1664.
21. Dortbudak O, Haas R, Bernhart T, Mailath-Pokorny G. Lethal photosensitization for decontamination of implant surface in the treatment of peri-implantitis. Clin Oral Implant Res 2001;12:104-108.