Vol. 45, N. 1 : pp. 59 - 66, March, 2002
ISSN 1516-8913 Printed in Brazil
BRAZILIAN ARCHIVES OF
BIOLOGY AND TECHNOLOGY
A N I N T E R N A T I O N A L J O U R N A L
Leaf Structure and Taxonomy of
Petunia and Calibrachoa
(Solanaceae).
Claudia dos Reis1*; Maria das Graças Sajo 1
and João Renato Stehmann2
1
Departamento de Botânica, IBUNESP, Rio Claro, C. Postal 199, CEP 13506-900, Rio Claro - SP, Brazil;
2
Departamento de Botânica, ICBUFMG. Av. Antônio Carlos, 6627, CEP 31270-110, Belo Horizonte - MG, Brasil
ABSTRACT
We studied the leaf anatomy of sixteen species of Calibrachoa and eight species of Petunia. In Calibrachoa leaves, the vascular bundles sheath (endodermis) was formed by parenchymatous developed cells, different from those of the mesophyll. In Petunia, this sheath did not show a marked morphological differentiation. The Calibrachoa leaves could be separated according to the type of leaf margins, the distribution of the stomata on leaf surfaces, the organization of the mesophyll and the morphology of the trichomes. Based on these results, an indented dichotomous identification key was elaborated for the species of the genus Calibrachoa.
Key words: Solanaceae, Calibrachoa, Petunia, leaf anatomy, taxonomy
*Author for correspondence
INTRODUCTION
Solanaceae is an important family for possessing countless ornamental, medicinal and nutritious
species (Heywood, 1993; Sauer, 1994). Petunia
Jussieu, which belong to the subfamily
Cestroideae Schltdl - tribe Nicotianeae G. Don.
(Hunziker, 1979; Cosa de Gastiazoro, 1991) is a genus with many ornamental species. It is native
of South America and is popularly known as the
garden petunia, a highly cultivated hybrid for ornamental purposes derivated from a cross
between Petunia axillaris (Lamarck) Britton,
Sterns & Poggemburg and P. integrifolia (Hooker)
Schinz & Thellung (Wijsman, 1982).
Studies accomplished during the last decade, concentrating mainly on the genetic improvement
of the garden petunia, showed that in Petunia
sensu Fries (1911) there wese two genetically distinct groups designated by Wijsman & Jong
(1985) as two genera: Petunia (2n=18) and
Stimoryne Rafin. (2n=14). According to this work, the garden petunia (2n=14) was included in the
genus Stimoryne. However, as the name Petunia
was previously established among horticulturists,
Wijsnands et al. (1986) suggested the conservation
of P. nyctaginiflora Juss. (2n=14) as the
nomenclatural type of the genus Petunia
(Brummitt, 1989) and revalidated the genus
Calibrachoa La Llave & Lex. (2n=18). Twelve
species remained in the genus Petunia (Ando &
Hashimoto 1996) and 24 were recombined to
Calibrachoa (Wijsman, 1990; Stehmann & Semir, 1997).
In his study on the limits between the genera
Petunia and Calibrachoa, Stehmann (1999)
confirmed that they wese morphologically and
cytologically distinct. Petunia is characterized by
(1991) studied the anatomical structure of the
vegetative organs of some species of Petunia
sensu lato, embracing representatives of the two genera; however, the author did not relate the results obtained to the taxonomic alterations of the
group. Recently, Watanabe et al. (1999) described
for Petunia sensu lato three different seed surface
patterns: 1) thick, wavy middle lamellae and
anticlinal cell walls (all the species of Petunia
sensu Wijsman), 2) thin, wavy middle lamellae
embedded in straight anticlinal cell walls (C.
parviflora and C. pygmaea), and 3) straight middle lamellae and anticlinal cell walls (all the other
species of Calibrachoa).
Anatomical studies with representatives of Solanaceae, are those of Ahmad (1964), Roe (1971), Bernardello (1982), Ogundipe (1992) and Cosa de Gastiazoro (1993; 1994).
The present study describes the leaf structure of
eight Petunia and sixteen Calibrachoa species
with the purpose of pointing out anatomical
characters useful for the separation of these two
generaand for the characterization ofCalibrachoa
species.
MATERIAL AND METHODS
Plant material was collected from its natural habitats in São Paulo, Paraná, Santa Catarina and Rio Grande do Sul States and the vouchers wase deposited in the Herbarium of the Universidade Estadual de Campinas (UEC) under the following
specifications: Calibrachoa caesia (Sendtn.)
Wijsman, J. R. Stehmann (1946) et al.; C.
C. sendtneriana (R. E Fr.) Stehmann & Semir, J.
R. Stehmann (1799) et al; C. spathulata (L. B. Sm.
& Downs) Stehmann & Semir, J. R. Stehmann
(1671) & A. Ippolito; Petunia altiplana T. Ando &
Hashim., J. R. Stehmann (1876) et al.; P. axillaris
(Lam.) Britton, Sterns &t Poggenb., J. R.
Stehmann (1579) et al; P. bonjardinensis T. Ando
& Hashim., J. R. Stehmann (1808) et al; P. exserta
Stehmann, J. R. Stehmann (1603); P.
guarapuavensis T. Ando & Hashim., J. R.
Stehmann (1688) & A. Ippolito; P. integrifolia
(Hook.) Schinz & Thell., J. R. Stehmann (1836) et
al; P. reitzii L. B. Sm. & Downs, J. R. Stehmann
(1741) et al; P. scheideana L. B. Sm. & Downs, J.
R. Stehmann (1721) et al.
Adult leaves, unrelated with the reproductive
organs, located between the 3rd and 8th nodeswere
fixed in FAA 70 (Johansen, 1940). The middle region of the leaves were cut by free hand or by a
microtome in the case of material included in
paraffin (Johansen, 1940); the sections were stained with astra blue and basic fucsin (Sass, 1951) and mounted in “Entellan” (included material) or glycerinated gelatin (free hand sections).
The diagrams were executed by hand under the optical microscope, using a camera lucida and projection of a micrometric scale. The a JVC video camera coupled to a photomicroscope and a computer.
RESULTS AND DISCUSSION
Figures 1-2 - Micrographs of the middle region of leaves of Calibrachoa and Petunia in cross section. 1)
Calibrachoa paranensis, 2) Petunia integrifolia. (En=endodermis).
These results brought evidence to support the
acceptance of Petunia sensu lato as an artificial
genus.
Of the 16 species of Calibrachoa studied (about
70% of the genus), only in Calibrachoa
parviflora and C. pygmaea the endodermis was less developed.
Cosa de Gastiazoro (1991) studied the anatomy
of the vegetative organs of Petunia sensu lato
but did not report any morphological differences in the endodermis leaves, in spite of having studied species of the two genera. According to
the same author, in Petunia the leaf structure
was relatively uniform and showed a dorsiventral mesophyll (in most of the species),
epidermis formed by isodiametric cells, stomata
on both surfaces and glandular, uniseriated
trichomes with a stalk of two to six cells. These
results could be due to the reduced number of
species studied: two from the genus Petunia
sensu stricto (P. axillaris and P. integrifolia),
one from Calibrachoa (P. parviflora) and the
other probably from Fabiana (P. patagonica
(Speg.) Millan) (Stehmann, 1999).
Different from the reported by Cosa de
Gastiazoro (1991) for Petunia,the leaf structure
in Calibrachoa is quite diversified, both
externally and internally. When observed in a transversal section, they can be revolute (Fig. 3, Table 1), semi-revolute (Fig. 4, Table 1), conduplicate (Fig. 5, Table 1), semi-conduplicate (Fig. 6, Table 1) or plane (Fig. 7, Table 1).
Morphological variations could be observed even among individuals of the same species (Table 1). In
C. sellowiana the leaves wese plane, conduplicate or
semi-conduplicate and in C. linoides they wese plane
or semi-revolute. But on both species the mesophyll
was always dorsiventral with apalisade parenchyma
in the adaxial surface and a spongyparenchyma with
lobed cells in the abaxial surface.
Considering the size of the epidermal cells and the distribution of the stomata in the surfaces, it was
possible to recognize in Calibrachoa two different
groups of leaves (Table 1): one with the epidermis
formed by cells of similar sizes and stomata
distributed on both surfaces (Fig. 8) and other with
larger epidermal cells in the
adaxial surface and stomata only in abaxial surface (Fig. 9). According to Metcalfe & Chalk (1957),
amphistomatic leaves are common in Solanaceae,
Figures 3-9 - Diagrams and micrographs of the middle region of Calibrachoa leaves in cross section. 3) C. ericifolia, 4) C. micrantha, 5, 8) C. sellowiana, 6) C. spathulata, 7) C. pygmaea, 9) C. sendtneriana (br = 100µm,
Vb= vascular bundle, St= stomata).
Glandular trichomes wese common in
Calibrachoa leaves. They presented a basal cell and a stalk that sustained an apical secretory cell
(Fig. 10, 11). The cell number of the trichome
stalk varies depending on the species. In a same
leaf, short trichomes (with a stalk varying between 1-3 cells) and long trichomes (stalk with 4-12 cells) wese found (Table 1). This was the case of
C. linoides where the number of cells in the stalk varied from 2-9 and 12. The variation in the trichomes size was used by Fries (1911) for
differentiating some species like Petunia
linoides / P. regnellii and P.
Among the studied species, Calibrachoa
eglandulata (Table 1) was the only one lacking glandular trichomes. In its leaves there wese uniseriate nonglandular trichomes (Fig. 12) formed by 6-11 cells. According to Stehmann & Semir (1997), this species probably lost the capacity to produce secretions, because glandula r trichomes wese present in all the remaining species of the genus, as well as in the species of
Figures 10-12 - Micrographsof the middle region of leaves of Calibrachoa in crosssection. 10) C. ericifolia, 11)
C. elegans, 12) C. eglandulata.
Analyzing the organization of the mesophyll, it
was possible to separate the leaves of Calibrachoa
into four different groups (Table 1), namely: a) leaves with a homogeneous mesophyll formed only by lobed cells of the spongy parenchyma (Fig. 13), b) leaves with a homogeneous mesophyll formed only by palisade parenchyma (Fig. 14), c) leaves with a dorsiventral mesophyll
Figures 13-16 - Micrographs of the middle region of leaves of Calibrachoa in crosssection. 13) C. ovalifolia, 14)
C. pygmaea, 15) C. paranensis, 16) C. eglandulata. (Ex=bundle sheath extension).
Druse crystals, as described for some species of Solanaceae (Ogundipe, 1992; Cosa de Gastiazoro, 1991; 1993; 1994), were observed in the
mesophyll of Calibrachoa sellowiana and C.
caesia (Fig. 17). As the occurrence of the druse was restricted to two species, this characteristic has a taxonomic value, aiding in the determination of these taxa.
The vascular bundle sheath (endodermis) was formed by parenchymatous cells which wese bigger than those of the mesophyll (Fig. 1). In most of the species, the endodermis presented extension to both leaf surfaces in the median vein
region (Fig. 15, Table 1). In C. micrantha leaves
(Fig. 16), the extension was only in the abaxial
surface and, in C. paranensis (Fig. 15), only in the
adaxial surface. In the leaves of C. heterophylla,
C. humilis, C. parviflora, C. ericifolia and C. pygmaea (Fig. 6), the endodermis did not present any extensions.
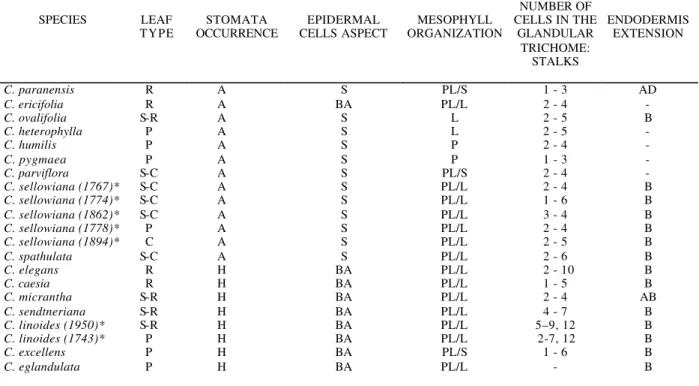
Considering the characteristics of the epidermis (size of the cells and distribution of the stomata), the organization of the mesophyll (morphology and distribution of the chlorenchyma, extension of the endodermis) and the number of cells of the trichomes stalks (Table 1), we separated the
species of Calibrachoa in the following way:
1. Non-glandular trichomes………...C. eglandulata
1. Glandular trichomes. 2. Amphistomatic leaves. 3. Revolute leaves.
4. Epidermal cells of similar sizes on both leaf surfaces………..…...C. paranensis
4. Larger epidermal cells on the adaxial surface...…C. ericifolia
6. Dorsiventral mesophyll.
9. Idioblast with druses in the mesophyll…...……….……C. sellowiana
9. Characteristic lacking.
10. Mesophyll with adaxial palisade parenchyma and abaxial lobed cells of the spongy
parenchyma……...……….C. spathulata
10. Mesophyll with adaxial palisade parenchyma and abaxial spongy
parenchyma……….…………...………….C. parviflora
2. Hypostomatic leaves.
11.Idioblast with druses in the mesophyll………..…………...………C. caesia
11. Characteristic lacking.
12. Revolute leaves………...………C. elegans
12. Semi-revolute or plane leaves.
13.Endodermis with an extension in the abaxial face in the median vein region
………...……..C. micrantha
13. Endodermis with extension in both leaf surfaces in the median vein region
14. Dorsiventral mesophyll with adaxialpalisade parenchyma and abaxial spongy
parenchyma…….………...………..C. excellens
14. Dorsiventral mesophyll with adaxial palisade parenchyma and abaxial lobed cells of the
spongy parenchyma.
15. Trichome stalkwith 4 to 7 cells………...………C. sendtneriana
15. Trichome stalkwith 2 to 7 and 12 cells………...……C. linoides
Table 1 – Anatomical characteristics of the Calibrachoa leaves.
SPECIES LEAF
TYPE
STOMATA OCCURRENCE
EPIDERMAL CELLS ASPECT
MESOPHYLL ORGANIZATION
NUMBER OF CELLS IN THE
GLANDULAR TRICHOME:
STALKS
ENDODERMIS EXTENSION
C. paranensis R A S PL/S 1 - 3 AD
C. ericifolia R A BA PL/L 2 - 4 -
C. ovalifolia S-R A S L 2 - 5 B
C. heterophylla P A S L 2 - 5 -
C. humilis P A S P 2 - 4 -
C. pygmaea P A S P 1 - 3 -
C. parviflora S-C A S PL/S 2 - 4 -
C. sellowiana (1767)* S-C A S PL/L 2 - 4 B
C. sellowiana (1774)* S-C A S PL/L 1 - 6 B
C. sellowiana (1862)* S-C A S PL/L 3 - 4 B
C. sellowiana (1778)* P A S PL/L 2 - 4 B
C. sellowiana (1894)* C A S PL/L 2 - 5 B
C. spathulata S-C A S PL/L 2 - 6 B
C. elegans R H BA PL/L 2 - 10 B
C. caesia R H BA PL/L 1 - 5 B
C. micrantha S-R H BA PL/L 2 - 4 AB
C. sendtneriana S-R H BA PL/L 4 - 7 B
C. linoides (1950)* S-R H BA PL/L 5–9, 12 B
C. linoides (1743)* P H BA PL/L 2-7, 12 B
C. excellens P H BA PL/S 1 - 6 B
C. eglandulata P H BA PL/L - B
Legends: R: revolute; S-R: semi-revolute; C: conduplicate; S-C: semi-conduplicate; A: amphistomatic; H: hypostomatic; S: similar; BA: big in the adaxial surface; PL: palisade parenchyma; S: spongy parenchyma; L: spongy parenchyma with lobed cells; AD: adaxial; B: both surfaces ; AB: abaxial; *: number of the collector.
ACKNOWLEDGEMENTS
This work was supported by CAPES.
RESUMO
Foram estudados, sob o ponto de vista anatômico, os limbos foliares de dezesseis espécies de
gênero Calibrachoa
.
REFERENCES
Ahmad, J. K. (1964), Cuticular studies in Solanaceae.
Canadian Journal of Botany, 42, 773-803.
Ando, T. and Hashimoto, G. (1996), A new Brazilian species of Petunia (Solanaceae) from interior Santa Catarina and Rio Grande do Sul, Brazil. Brittonia,
48(2), 217-23.
Bernardello, L. M. (1982), Estudios en Lycium (Solanaceae) I. Anatomia de la hoja y tallo, y sus diferencias con Grabowskia. Boletin de la Sociedad Argentina de Botanica, 21(1-4), 153-185.
Brummitt, R. K. (1989), Report of the Committee for Spermatophyta: 36. Taxon, 38, 301.
Cosa de Gatiazoro, M. T. (1991), Estudio morfoanatomico de organos vegetativos em Cestroideae (Solanaceae), I: Tribu Nicotianeae.
Kurtziana, 21, 111-152.
Cosa de Gastiazoro, M. T. (1993), Estudio morfoanatomico de organos vegetativos em Cestroideae (Solanaceae), II: Tribu Salpiglossideae.
Kurtziana, 22, 47-72.
Cosa de Gastiazoro, M. T. (1994), Estudio morfoanatomico de organos vegetativos em Cestroideae (Solanaceae), III: Tribu Schwenckieae.
Kurtziana, 23, 9-25.
Elliger, C. A.; Wong, R. Y.; Benson, M.; Gaffield, W. and Waiss, A. C. (1982), Diterpenes of Calibrachoa parviflora. Journal of Natural Products, 55, 1477-1487.
Fries, R. E. (1911), Die Arten der Gattung Petunia. Kongl. Svenska Vetenskapsakad. Handl., 461-472. Heywwod, V. H. (1993), Flowering plants of the world.
Oxford University Press, New York.
Stehmann, J. R. (1999), Estudos taxonômicos na tribo Nicotianeae G. Don (Solanaceae): revisão de Petunia
Jussieu, das espécies brasileiras de Calibrachoa La Llave & Lexarza e o estabelecimento de um novo gênero, Petuniopsis Stehmann & Semir. Tese, Universidade Estadual de Campinas, Campinas, Brazil.
Stehmann, J. R. and Semir, J. (1997), A new species and new combinations in Calibrachoa (Solanaceae),
Novon, 7, 417-419.
Watanabe, H.; Ando, T.; Nishino, E.; Kokubun, H.; Tsukamoto, T.; Hashimoto, G. and Marchesi, E. (1999), Three groups of species in Petunia sensu Jussieu (Solanaceae) inferred from the intact seed morphology. American Journal of Botany, 86(2), 302-305.
Wijnands, D. O; Bos, J. J.; Wijsman, W. J.; W, Schneider, F.; Brickell, C. D. and Zimmer, K. (1986), (856) Proposal to conserve 7436 Petunia with P. nyctaginiflora as typ. cons. (Solanaceae), Taxon, 35, 748-749.
Wijsman, H. J. W. (1982), On the Interrelationships of certain Species of Petunia. I. Taxonomic notes on the Parental Species of Petunia hybrida. Acta Botanica Neerlandica, 31, 477-490.
Wijsman, H. J. W.(1983), On the Interrelationships of certain Species of Petunia. II. Experimental Data: crosses between Different taxa. Acta Botanica Neerlandica, 32, 97-107.
Wijsman, H. J. W. (1990), On the inte-relationships of certain species of Petunia VI. New names for the species of Calibrachoa formely included into Petunia
(Solanaceae), Acta Botanica Neerlandica, 39(1), 101-102.
Wijsman, H. J. W., Jong, J. H. (1985), On the interrelationships of certain species of Petunia IV. Hybridization between P. linearis and P. calycina