J. Braz. Chem. Soc. vol.21 número9
Texto
Imagem


Documentos relacionados
Subsequently, after a review of the literature and attempting to obtain aromatic pyrazoles for further biological assays, we chose thionyl chloride/ pyridine as
This step comprised the evaluation of the effect of the solvent nature, investigation of the influence of the reagent concentration and evaluation of the time required
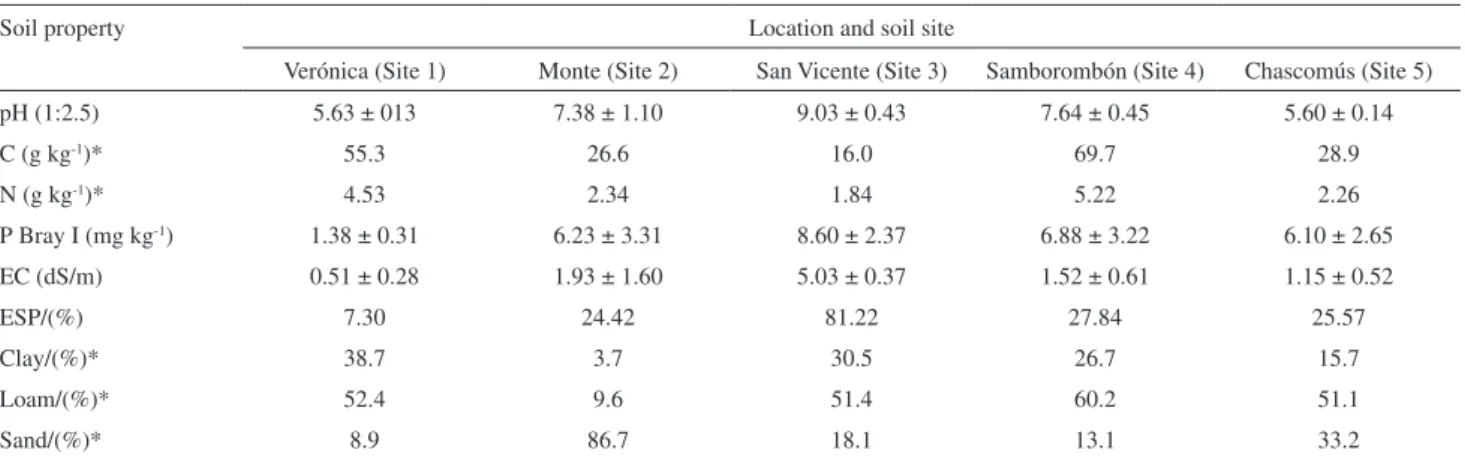
The oil constituents including biosynthetic classes and total phenol/tannin contents (treated as species data set) and soil parameters and foliar nutrients (environmental
Though the nature of catalytic species generated in situ is not known when the reaction was performed in the absence of PPh 3 affording the desired product in 15%
Stripping Voltammetric Determination of Cadmium in Sea Water using a Carbon Paste Electrode Modiied with Alginic Acid from Brown Algae.. Carolina Muñoz,* Manuel Zúñiga and
Initially, to different flasks containing aqueous solutions of chitosan (0.8 g in 0.1%, m/v chloridric acid) sulfosuccinic acid was added to obtain a inal composition range of
Therefore, a corresponding decrease in free T 3 and T 4 levels in the blood serum of the treated animals with respect to control will indicate antithyroid effects of
The speciic capacity obtained is about 60% higher than that obtained for lat MnO 2 surfaces showing that, in spite of the wettability being lower, some penetration
