w ww . e l s e v i e r . c o m / l o c a t e / b j p
Original
Article
Distribution
patterns
of
flavonoids
from
three
Momordica
species
by
ultra-high
performance
liquid
chromatography
quadrupole
time
of
flight
mass
spectrometry:
a
metabolomic
profiling
approach
Ntakadzeni
Edwin
Madala
a,∗,
Lizelle
Piater
a,
Ian
Dubery
a,
Paul
Steenkamp
a,baDepartmentofBiochemistry,UniversityofJohannesburg,AucklandPark,SouthAfrica bCSIRBiosciences,NaturalProductsandAgroprocessingGroup,Pretoria,SouthAfrica
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received15January2016 Accepted14March2016 Availableonline20May2016
Keywords: Momordica
UHPLC-qTOF-MS Flavonoids
Principalcomponentanalysis Chemotaxonomy
Hierarchicalclusteranalysis
a
b
s
t
r
a
c
t
PlantsfromtheMomordicagenus,Curcubitaceae,areusedforseveralpurposes,especiallyfortheir nutri-tionalandmedicinalproperties.Commonlyknownasbittergourds,melonandcucumber,theseplants arecharacterizedbyabittertasteowingtothelargecontentofcucurbitacincompounds.However, sev-eralreportshaveshownanundisputedcorrelationbetweenthetherapeuticactivitiesandpolyphenolic flavonoidcontent.Usingultra-highperformanceliquidchromatographyquadrupoletimeofflightmass spectrometryincombinationwithmultivariatedatamodelssuchasprincipalcomponentanalysisand hierarchicalclusteranalysis,threeMomordicaspecies(M.foetidaSchumach.,M.charantiaL.andM. bal-saminaL.)werechemo-taxonomicallygroupedbasedontheirflavonoidcontent.Usingaconventional massspectrometric-basedapproach,thirteenflavonoidsweretentativelyidentifiedandthethreespecies werefoundtocontaindifferentisomersofthequercetin-,kaempferol-andisorhamnetin-O-glycosides. OurresultsindicatethatMomordicaspeciesareoverallveryrichsourcesofflavonoidsbutdocontain differentformsthereof.Furthermore,tothebestofourknowledge,thisisafirstreportontheflavonoid contentofM.balsaminaL.
©2016SociedadeBrasileiradeFarmacognosia.PublishedbyElsevierEditoraLtda.Thisisanopen accessarticleundertheCCBY-NC-NDlicense(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Introduction
Momordicaspeciesareversatileplantsbelongingtothefamily Cucurbitaceaeandareoftenreferredtobyvariouscommonnames suchasbittergourd,bittercucumberandbittermelon(Nagarani etal.,2014a).Indifferentpartsoftheworld,Momordicaplantsare consumedasavegetableandareknownfortheirbittertastedueto thepresenceofphytochemicalssuchasalkaloidsandcucurbitacins (Chenetal.,2005;Riosetal.,2005;Nagaranietal.,2014a).Within thisgenusthereareseveralspecieswidelydistributedacrossthe globe,mainlyinthetropicalandsubtropicalregionsofAfrica,Asia andAustralia.InarecentreviewarticlebyNagaranietal.(2014a), undisputedscientificevidenceontheoriginoftheseplantswas presentedandbelievedtobeendemictoIndia,whileearlier bio-geographicaloriginsofthesespeciescanbefoundelsewhere(Dey etal.,2006;Singhetal.,2007;Gaikwadetal.,2008).
Apartfrom thenutritious value,Momordica species arealso usedfortheirmedicinalproperties.Forexample,M.charantiahas
∗ Correspondingauthor.
E-mail:[email protected](N.E.Madala).
beenusedin Chinesefolkmedicineforthetreatmentof differ-entchronicdiseases(Zhang,1992).InsomepartsofSouthAfrica, thesespeciesarecurrentlyusedasmedicationforsugardiabetes andchronichypertensiondiseases,withoutanyscientificbacking. DuetothewiderangeofphytochemicalsinMomordica(Nagarani etal.,2014a),itisverydifficulttopointoutoneactivecompound, eventhoughnumerousmetaboliteswithknownpharmacological activitieshave beenidentified (Singhet al.,2011; Kenny etal., 2013; Nagaranietal., 2014b).Momordicaspecies are knownto containlargequantitiesofpolyphenoliccompounds(Kubolaand Siriamornpun,2008)andamongsttheseareflavonoidswhichare knowntopossess severaltherapeuticactivities.Furtherreports havesuggestedapossiblelinkbetweenthemedicinalpropertiesof
Momordicaspeciesandtheirflavonoidcontent(Horaxetal.,2005; LinandTang,2007;WuandNg,2008;Zhuetal.,2012).The distri-butionofflavonoidsintheseplantsrepresentsanotherinteresting dimension.Forinstance,Kennyetal.(2013)reportedthepresence offlavonoidsinthebittermelon(M.charantia)fruit,however,the levelsvaryacrossdifferentsectionsofthefruit.Inaseparatestudy, Nagaranietal.(2014b)reportedaveryinterestingdistribution pat-ternacrossdifferentspeciesofMomordica(M.tuberosa,M.charantia
andM.cochinchinensis).Here,theflavonoidrutinwasdetectedin
http://dx.doi.org/10.1016/j.bjp.2016.03.009
theformertwospeciesbutabsentinthelater.Assuch,theaim ofthecurrentstudywastoinvestigatetheflavonoiddistribution patternswithinthethreeMomordicaspecies(namelyM. charan-tiaL.,M.foetidaSchumach.andM.balsaminaL.)currentlyusedfor diabeticcontrolandnutritionalvalueintheLimpopo(Northern) regionsofSouthAfrica.
Inadditiontotheirnutritionalandmedicinalproperties,plant metabolitescanalsobeusedtotaxonomicallyclassifyplantsto ensurethatthecorrectspecie(s)areusedformedicinalpurposes.In thepast,flavonoidshavebeenusedforchemo-taxonomical classifi-cationofplants(Iwashina,2000;ElShabrawyetal.,2014;Martucci etal.,2014).Toachieveourobjective,thecurrentstudywasdivided intotwoparts:thefirstaimingtoestablishachemo-taxonomical relationshipbetweenthethreespeciesusingametabolomic profil-ingapproachwiththeaidofUHPLC-qTOF-MSandmultivariatedata models,andinthesecond,theflavonoidcompositionofthethree specieswasinvestigatedusingtargetedMS-basedflavonoid iden-tification strategies presented elsewhere (Cuyckens and Claeys, 2004).
Materialsandmethods
Plantmaterialsandchemicals
Momordicaplants,Curcubitaceae,werecollectedinandaround theVendaregionofSouthAfricawiththehelpofthelocalfarmers. Briefly,M.charantiaL.wascollectedfromafarmintheNwanedi farmingarea,about80kmsouthoftheZimbabweanborder.The othertwospecies,M.balsaminaL.andM.foetidaSchumach.were collectedfromvariousvillagesaroundThohoyandou.Thespecies wereidentifiedwiththehelpofMrPhilipRamela(Madzivhandila CollegeofAgriculture,SouthAfrica)andforfurtherconfirmation, theplantmaterialswerealsocomparedtothenational herbar-iumspecimensattheSouthAfricanNationalBiodiversityInstitute (SANBI) (Pretoria, South Africa). Voucher herbarium specimens (withvouchernumberNEM003(M.balsamina),NEM004(M. cha-rantia)and NEM005 (M.foetida)) wereprepared and deposited totheDepartmentofBotany,UniversityofJohannesburg.Unless statedotherwise,allthechemicalswereofanalytical gradeand obtainedfromvariousinternationallyreputablesuppliers.Boththe methanoland acetonitrile(Romil, MicroSep,South Africa)were usedforUHPLC-qTOF-MSanalyses.Waterwaspurifiedwitha Milli-Q Gradient A10system (Millipore, Billerica, MA, USA). Leucine encephalin, rutin and formic acidwere purchased from Sigma Aldrich,Germany.Quercetin-3-glucose,quercetin-4′-glucoseand quercetin-7-glucosewerepurchasedfromPhytolab (Vestenbergs-greuth,Germany).
Metaboliteextraction
TheleavesofthethreeMomordicaspecieswereair-driedat37◦C forthreeconsecutivedays.Metaboliteswereextractedfromthe fourindependentlycrushedleafsamples(2g),representingfour independentbiological replicates, using80% aqueous methanol (20ml).Formaximumextraction,thehomogenatewasplacedon anorbitalshakeratroomtemperature(25◦C)foratleast30min. Aftertheextraction,thetissue debriswasremovedby centrifu-gation at5000×g for 10min.The supernatant was driedto at least1mlusingarotaryevaporatoroperatingat55◦Cunder nega-tivepressurevacuum.The1mlextractwassubsequentlydriedto completenessusingavacuumconcentratorcentrifuge(Vacufuge, Eppendorf,Germany)operatingat55◦C.PriortoUHPLC–MS analy-ses,thepelletedextractresidueswerere-constitutedin1mlof50% aqueousmethanolandfilteredthrough0.22mnylonfilters.
UHPLC–MSanalyses
For UHPLC–MS analyses, a previously described method (Madala et al., 2014a) was used. Briefly, one (1) l of the extracts was analysed on a Waters Acquity BEH C8 column (150mm×2.1mm,1.7mparticlesize)andthetemperature con-trolledat60◦C.Here,threetechnicalreplicateswereanalyzedand randomizedduringtheUHPLC–MSanalysesusingonline random-izingsoftware(www.random.org/lists/)toavoidtechnicalbias.A binarysolventmixturewasusedconsistingofwatercontaining 10mMformicacid(pH2.3)(eluentA)andacetonitrilecontaining 10mMformicacid(eluentB).Theinitialconditionswere98%Aata flowrateof0.4ml/minandmaintainedfor1min,followedby mul-tiplegradientsto5%Aat26min.Theconditionswerekeptconstant for1minandthenchangedtotheinitialconditions.Thetotal chro-matographicruntimewas30min.Chromatographicelutionwas monitoredwiththeaidofaphotodiodearray(PDA)detectorand MS.
ForMSdetection,ahighresolutionmassspectrometer(Waters SYNAPTG1Q-TOFsystem),operatinginV-opticsandelectrospray negativemode,wasused.Leucineenkephalin(50pg/ml)wasused asreferencelockmasscalibranttoobtaintypicalmassaccuracies between1and 5mDa.Theoptimalconditionsforanalysiswere asfollows:capillaryvoltageof2.5kV,thesamplingconeat30V and theextraction cone at4V. Thescan time was0.1s cover-ing the 100–1000Da mass range. The source temperature was 120◦Candthedesolvationtemperaturewassetat450◦C. Nitro-genwasused as thenebulizationgasat a flow rateof 700l/h. Toobtainbettermetabolitecoverageandfragmentationpatterns thereof,theMSwasoperatedatdifferentcollisionenergy(CE) lev-elsasreportedelsewhere(Madalaetal.,2012).For comparison purposes,authenticstandards(quercetin-3-glucose,quercetin-4′ -glucose and quercetin-7-glucose) werealso analyzed usingthe sameconditions.Alltheacquisitionandanalysisofdatawere con-trolledbyWatersMassLynxTMv4.1software(SCN704).
Multivariatedataanalyses
Primaryrawdatawasanalyzedbydataalignment,peakfinding, peakintegrationandretentiontime(Rt)correctionusinga Mark-erlynxXSTMsoftware(WatersCorporation,Milford,USA)withthe followingprocessingparameters:Rtrangeof7–12min,massrange of100–1000Da,masstoleranceof0.02Da,Rtwindowof0.2min. The resulting datasets were exportedto theSIMCA-P software version13.0(Umetrics,Umea,Sweden)forPrincipalComponent Analysis(PCA)andHierarchicalClusterAnalysis(HCA).Beforethe models were computed, all the data were mean centered and
Pareto-scaled.ForHCAanalysis,theWarddistancealgorithmwas usedtocalculatethedistancebetweenthedifferentgenerated clus-ters(Madalaetal.,2014b).
Resultsanddiscussion
ClassificationofMomordicaspeciesbasedontheirflavonoid content
100
0
6.00
9.28 489.1061
8.85; 447.0978
7.68 300.0248
8.15
8.56 285.0367
8.85; 447.0908 11.32 335.1167 463.0836
7.20 625.1361
5.47; 293.1082
8.15 463.0778
4.81 285.0603
8.56 285.0327
9.61
315.0475 12.60 301.1333 489.0988
9.88 9.32 489.1041
12.62 285.0372
13.12
327.2150 285.035815.93 17.55 285.0416
8.00 10.00 12.00 14.00 16.00 18.00 20.00
6.00 8.00 10.00 12.00 14.00 16.00 18.00 20.00
6.00 8.00 10.00 12.00
Retention time (min)
Momordica balsamina Momordica charantia Momordica foetida
14.00 16.00 18.00 20.00
%
100
0
%
100
0
%
Fig.1. Selectedionchromatograms(XIC)showingrelativechromatographicelutionratesofdifferentflavonoidisomersinthethreeMomordicaspecies.TheXICwere extractedusingtheaglyconem/zof301,285and315representingquercetin,kaempferolandisorhamnetin.
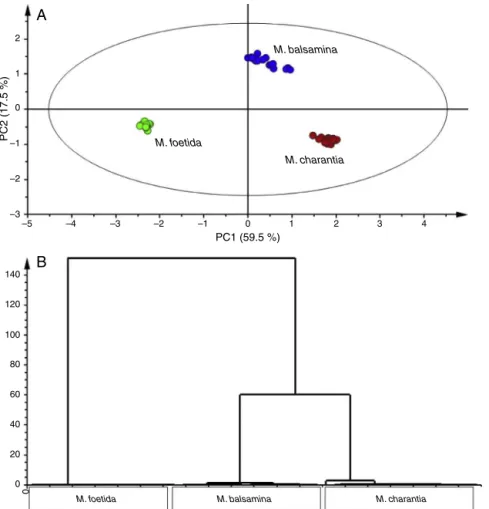
taxonomically-relatedplantstoproduceverysimilar phytochemi-calsand,assuch,flavonoidshavebeenusedforchemo-taxonomical classification (El Shabrawy et al., 2014; Martucci et al., 2014). Toevaluatethetaxonomicalconnection/relationshipbetweenthe threespecies,PCAwasperformedandthescoreplotshowedthat
thethree speciesare distinctfromoneanother (Fig.2A). How-ever,PCAmodelsareknowntobelesssuperiorinshowingsample relationships(Madalaetal.,2014b).Toovercomethischallenge, HCA results (Fig. 2B) were evaluated and it was seen that M. foetida forms a distinctive group whilst the other two species
2
A
B
1
0
–1
–2
–3
140
120
100
80
60
40
20
0
0
–5 –4 –3 –2 –1 0
PC1 (59.5 %)
M. charantia
M. charantia M. foetida
M. foetida
M. balsamina
M. balsamina
PC2 (17.5 %)
1 2 3 4
Aglycone R1 R2 R3 [Y0–H]– [Y0–
Y0
– H
O O aglycone
OH OH OH
OR1 R3
O
O OH R2O
HO
H]–
/Y0–
Quercetin H H OH 300/301
Kaempferol H H H 284/285
Isorhamnetin H H OCH3 314/315
O
Scheme1. Thestructureofcommonflavonoidaglyconesandcharacteristicfragmentsions([Y0−H]−andY0−)formedduringhomolyticandheterolyticsugarcleavage,
respectively.
formaverycloseclade.Overall,theseresultssuggestarelatively close relationship between M. balsaminaand M. charantia, but thesetwo (asa group/clade)have a distance relationshipwith
M.foetida.
Generalapproachforidentificationofflavonoids
FromUHPLC-qTOF-MSbaseionpeak(BIP)chromatograms,ions representingtypicalflavonoidaglyconefragmentswereselected andusedtogeneratethesingleionchromatograms(Fig.1).From the MS spectra, molecular formulae of the pseudo-molecular ions ([M−H]−) representing quercetin (m/z 300.021/301.029), kaempferol (m/z 284.025/285.040) and isorhamnetin (m/z
315.045) were generated and selected based on the criterion that the mass difference between the measured and calcu-latedmass is below 5ppm. The generated molecular formulae were further used for compound identification searches using
the Dictionary of Natural Products (DNP) online database (http://dnp.chemnetbase.com/) and the KNapSAcK database (http://kanaya.naist.jp/knapsackjsp/top.html).Massspectra gen-eratedatdifferentCEswerealsousedtoelucidate theposition andnumberofsugarmoleculesattachedtotheaglyconeskeleton. The proposed identities of the flavonoids and their respective MS fragmentation patterns were also compared to literature (CuyckensandClaeys,2004;Gobbo-Netoetal.,2008).
Structurally, flavonoids are polyphenolic compounds with a nuclear structure base of C6–C3–C6 (Scheme 1, Cuyckens and Claeys,2004;Tanetal.,2014).Thesecompoundsexistaseither aglyconesorremainglycosylatedwithdifferentsugarmoieties.It isalsoworthmentioningthatsugarattachmentontheflavonoid aglyconemoietycanhappenatdifferentpositions and,assuch, deducingthesugar position becomesa difficult analytical task. Thisadds tothestructural complexitywhich makes flavonoids difficultto identify. However, it hasbeen reportedthat during
[M-H]– Rt
(min)
R1 R2 R3 M.
balsamina M. charantia
M. foetida
Molecular Identity
OR1 R3
OH
O
O OH R2O
formula (< 5 ppm)
625 7.2 gly gly OH x x x quercetin-3,7-O
-diglucopyranoside
C27H30O17
595 7.46 di-gly H OH x x x quercetin 3-O
-sambubioside
C26H28O16
609 7.83 di-gly H OH x x x quercetin 3-O-rutinoside C27H30O16
579 8.12 di-gly H OH x x x quercetin-3-O-
arabinoside-rhamnopyranoside
C26H27O15
609 8.3 gly gly OCH3 x isorhamnetin-3-O
-glucoside-7-O -arabinopyranoside
C27H30O16
593 8.48 di-gly H OH x x x quercetin-3,7-di-
rhamnopyranoside
C27H30O15
593 8.56 di-gly H H x x kaempferol-3-O-rutinoside C27H30O15
447 8.85 gly H H x x x kaempferol-3-O-glucoside C21H20O11
533 9.32 H gly H x x kaempferol-7-O
-malonylglucoside
C24H22O14
533 9.5 gly H H x kaempferol-3-O
-malonylglucoside
C24H22O14
477 9.6 gly H OCH3 x x isorhamnetin 3-O-glucoside C22H22O12
533 9.87 H H gly x kaempferol-4’-O
-malonylglucoside
C24H22O14
489 10.01 gly H H x kaempferol-3-O
-acetylglucoside
C23H22O12
100
0
%
100
300.0211
A
B
C
302.0401
609.1453
610.1559
608.2439 367.0691
173.0092
173.0069 301.0376 367.0576 521.1978 608.2651
609.1472
610.1554 611.1740
609.1450
173.0046
284.0276 255.0241 227.0287
285.0442 286.0307 361.1481
447.0893
448.1083
450.1090 547.1911 633.2510 832.2695 895.2112 950.3019
949.3366 895.2121
832.2465 633.2555
537.2037 452.1095 448.1003 447.0902
361.1922 285.0374 255.0372
227.0689 285.0339 361.1808
447.0959
448.0982
450.1372 537.1966 605.2307723.2059832.2444 895.2068 949.3102
949.3387 895.2114 832.2424
624.1481 547.2236 448.1128
447.0903
361.1722 261.0999
178.8829
315.0449
314.0433 316.0509
463.0815 517.0784
609.1399 610.1476
729.1909 814.1890 846.2151 930.4323
822.3264 789.3167 612.2260
610.1514 609.1422
537.2060 463.0984 315.0446
297.0554 167.0610
193.0323 315.1687 463.0785 579.1420
609.1438
610.1492
677.2391 789.3088
961.1688
939.3214
3 eV 10 eV 20 eV 30 eV 3 eV 10 eV 20 eV
30 eV 10 eV 20 eV 30 eV
853.2057 789.3243 677.2489
610.1359 609.1395
491.0126 463.0932 315.1480
193.0589 167.0249
262.1064 367.0659 521.1878
609.1459
610.1506
677.1432 916.2618 977.2323
367.0724 262.1056
173.0146
610.1495
612.1707 776.2944 916.2798 977.2341
733.1944 916.2770 977.2595
612.1740 733.1738 863.2634 916.2659
0
%
100
0
%
100
0
100 150 200 250 300 350 400 450 500 550 600 650 700 750 800 850 900 950 1000
%
100
0
%
100
0
%
100
0
%
100
0
100 150 200 250 300 350 400 450 500 550 600 650 700 750 800 850 900 950 1000
%
100
0
%
100
0
%
100
0
%
100
0
100 150 200 250 300 350 400 450 500 550 600 650 700 750 800 850 900 950 1000
%
Fig.4. Massspectrageneratedatdifferentcollisionenergylevelsshowingtherespectivefragmentationpatternsofquercetin(A),kaempferol(B)andisorhamnetin(C) containingflavonoids.
negativemodeESI,deprotonatedflavonoid-O-glycosides[M−H]− ionsundergoaseriesoffragmentationstages duringMS analy-ses.Thisfragmentationistypicallycharacterizedbytheimmediate lossofsugarwhichcanhappenintwodifferentways.Asshown
2014).Theratioof[Y0−H]•−/Y0•−hasbeenusedwithsome suc-cessforthedeterminationofthesugarposition.Forinstance,Geng etal.(2009)couldpositivelydistinguishthedifferentisomersof quercetinbasedonthesugarpositionusingthreedifferentmass analyzers.However,theseobservationswerefoundtobe instru-ment(MS analyzer) specific.As such, in thecurrent study,the proposalmadeby Genget al.(2009) wasre-affirmedwithour Q-TOF-MSusingthreepositionalisomersofquercetinglycosides, namelyquercetin-3-glucose,quercetin-4′-glucoseand quercetin-7-glucose. Our results (supplementary file 1) suggest that our Q-TOF-MSinstrumentisalsocapableofshowingthesugar posi-tionontheaglyconeflavonoidmoieties.Furthermore,itisknown thatflavonoidscanbeglycosylatedatdifferentpositionswithtwo ormoresugarsattachinginthesameordifferentpositions(Zhou etal.,2014).Here,atmoderateCElevels,moleculesshowingan abundant([Y0−H]•−)wereidentifiedasthoseglycosylated at3′ positionandthosewithanintenseY0•−wereregardedasthose glycosylatedat7′or4′position(Gengetal.,2009).Furthermore, moleculesexhibiting equalintensitiesof both radicalaglycones ([Y0−H]•−/Y0•−)wereregardedasthoseglycosylatedinthetwo possibleglycosylationsites(Gengetal.,2009;AblajanandTuoheti, 2013;Zhouetal.,2014).Therefore,withtheaidoftheabove infor-mationandthoroughvisualinspectionofthemass spectra,the flavonoidcompositionof thethree Momordicaspeciescouldbe preciselydetermined(Fig.3).
Identificationofquercetinflavonoids
For flavonoids containing the quercetin aglycone, m/z
300.021/301.029 was used togenerate the extracted ion chro-matograms representing possible quercetin-bearing molecules (Fig.1).BycomparingtheMSchromatogramsgeneratedusingaCE of3eVandthatof30eV,atleastfivequercetinflavonoidisomers were positively identified from all three plant species (Fig. 3). However,notallthreespecieswerefoundtocontainallthe iso-mers.Moreinterestingly,thehigherCE(30eV)MSchromatograms werecapableofshowingthepositionofthesugarmoreefficiently thanthelowerCE(3eV)(Fig.4A–C).Thisobservationisconsistent withpreviouslypublisheddata(Gengetal.,2009)whereitwas shownthat [Y0−H]•−/Y0•− ratioincreases concomitantwithan increaseinCE.
Identificationofkaempferolflavonoids
Similarly,kaempferol-bearingflavonoidswerealsoidentified usingtheapproachas describedfor quercetinflavonoids. How-ever, m/z 284.025 ([Y0−H]−) and 285.040 (Y0−), representing thedeprotonatedforms of kaempferol aglycones,were usedto generatetheextractedionchromatogram(Fig.1)showingthe dif-ferentretentiontimesofkaempferolflavonoidsisomers.Atleast six kaempferol-containing flavonoids were positively identified betweenthethreespecies,withM.foetidacontaininglarge quan-titiesofthesecompounds,incomparisontotheothertwospecies (Fig.3).
Identificationofisorhamnetinflavonoids
In comparison to the former two types of flavonoids, the isorhamnetinclasswasfoundtobetheleastabundantinthethree
Momordicaspeciesinvestigated.Here,onlytwodifferentisomers (Fig.3)wereidentified,withtheM.balsaminahavingthree iso-mersandM.foetidaonlyone.Surprisingly,theM.charantiaspecies wasfoundtonotcontainanyisorhamnetin-bearingflavonoids.Itis thesedifferencesintheflavonoidcontentwhichmakesthecurrent studyimportantbecauseitrevealsinterestingunderlying biochem-icaldifferencesbetweenthethesespecies which,in part,could
possiblybeusedfortaxonomicalandotherbiologicalclassification systems.
Conclusion
WhilesomereportsontheflavonoidsfromM.charantiaandM. foetidaexist,tothebestofourknowledgethisisthefirstreport ontheflavonoidcompositionofM.balsamina.Theresultssuggest thatthethreespeciesarechemo-taxonomicallyrelatedandcontain verysimilarflavonoidcompositions.Fromtheresultsitcanalsobe seenthatM.charantiaandM.balsaminaaremorecloselyrelatedto eachotherand,asagroup,aredistantlyrelatedtoM.foetida.Above all,theresultsofthecurrentstudyconfirmtheMomordicaspecies asarichsourceofstructurallydiverseflavonoids.Theresultsalso reaffirmtheuseofLC–MSincombinationwithmultivariatedata modelstobeafeasibleapproachtostudymetabolitedistribution patternsbetweencloselyrelatedplantspecies.
Authors’contributions
NEM, IAD and LAP planned the study. NEM collected the plants and extracted the metabolites. NEM, LAP, IAD and PAS conductedtheUHPLC-qTOF-MS,multivariatedatamodelsand exe-cutedmetaboliteidentification.Allauthorsreadandapprovedthe finalmanuscript.
Conflictsofinterest
Theauthorsdeclarenoconflictsofinterest.
Acknowledgements
TheauthorswouldliketothanktheUniversityofJohannesburg andtheNRFforfinancialsupport.MrMuadeisthankedforthe donationofM.charantiaplants.
AppendixA. Supplementarydata
Supplementarymaterialrelatedtothisarticlecanbefound,in theonlineversion,atdoi:10.1016/j.bjp.2016.03.009.
References
Ablajan,K.,Tuoheti,A.,2013.Fragmentationcharacteristicsandisomeric differen-tiationofflavonolO-rhamnosidesusingnegativeionelectrosprayionization tandemmassspectrometry.RapidCommun.MassSpectrom.27,451–460.
Chen,J.,Chiu,M.,Nie,R.,Cordell,G.,Qiu,S.,2005.Cucurbitacinsandcucurbitane glycosides:structuresandbiologicalactivities.Nat.Prod.Rep.22,386–399.
Cuyckens,F.,Claeys,M.,2004.Massspectrometryinthestructuralanalysisof flavonoids.J.MassSpectrom.39,1–15.
Dey,S.S.,Singh,A.K.,Chandel,D.,Behera,T.K.,2006.Geneticdiversityofbittergourd MomordicacharantiagenotypesrevealedbyRAPDmarkersandagronomictraits. Sci.Hortic.109,21–28.
ElShabrawy,M.O.,Hosni,H.A.,ElGarf,I.A.,Marzouk,M.M.,Kawashty,S.A.,Saleh, N.A.,2014.FlavonoidsfromAlliummyrianthuBoiss.Biochem.Syst.Ecol.56, 125–128.
Gaikwad,A.B.,Behera, T.K.,Singh,A.K.,Chandel,D.,Karihaloo, J.L.,Staub,J.E., 2008.Amplifiedfragmentlengthpolymorphismanalysisprovidesstrategiesfor improvementofbittergourd(MomordicacharantiaL.).HortScience43,127–133.
Geng,P.,Sun,J.,Zhang,R.,He,J.,Abliz,Z.,2009.Aninvestigationofthefragmentation differencesofisomericflavonol-O-glycosidesunderdifferentcollision-induced dissociationbasedmassspectrometry.RapidCommun.MassSpectrom.23, 1519–1524.
Gobbo-Neto,L.,Gates,P.J.,Lopes,N.P.,2008.Negativeion‘chip-based’nanospray tandemmassspectrometryfortheanalysisofflavonoidsinglandulartrichomes ofLychnophoraericoidesMart.(Asteraceae).RapidCommun.MassSpectrom.22, 3802–3808.
Horax,R.,Hettiarachchy,N.,Islam,S.,2005.Totalphenoliccontentsand pheno-licacidconstituentsin4varietiesofbittermelons(Momordicacharantia)and antioxidantactivitiesoftheirextracts.J.FoodSci.70,C275–C280.
Kenny,O.,Smyth,T.J.,Hewage,C.M.,Brunton,N.P.,2013.Antioxidantproperties andquantitativeUPLC–MSanalysisofphenoliccompoundsfromextractsof fenugreek(Trigonellafoenum-graecum)seedsandbittermelon(Momordica cha-rantia)fruit.FoodChem.141,4295–4302.
Kubola,J.,Siriamornpun,S.,2008.Phenoliccontentsandantioxidantactivitiesof bittergourd(MomordicacharantiaL.)leaf,stemandfruitfractionextractsin vitro.FoodChem.110,881–890.
Lin,J.Y.,Tang,C.Y.,2007.Determinationoftotalphenolicandflavonoidcontents inselectedfruitsandvegetables,aswellastheirstimulatoryeffectsonmouse splenocyteproliferation.FoodChem.101,140–147.
Martucci,M.E.P.,DeVos,R.C.,Carollo,C.A.,Gobbo-Neto,L.,2014.Metabolomicsas apotentialchemotaxonomicaltool:applicationinthegenusVernoniaSchreb. PLoSONE9,e93149.
Madala,N.E.,Steenkamp,P.A.,Piater,L.A.,Dubery,I.A.,2012.Collisionenergy alter-ationduringmassspectrometricacquisitionisessentialtoensureunbiased metabolomicanalysis.Anal.Bioanal.Chem.404,367–372.
Madala, N.E.,Tugizimana, F.,Steenkamp,P.A., 2014a. Developmentand opti-mizationofanUPLC-QTOF-MS/MSmethodbasedonanin-sourcecollision induceddissociationapproachforcomprehensivediscriminationof chloro-genic acids isomers from Momordica plant species. J. Anal. Met. Chem.,
http://dx.doi.org/10.1155/2014/650879.
Madala,N.E.,Piater,L.A.,Steenkamp,P.A.,Dubery,I.A.,2014b.Multivariatestatistical modelsofmetabolomicdatarevealsdifferentmetabolitedistributionpatterns inisonitrosoacetophenone-elicitedNicotianatabacumandSorghumbicolorcells. SpringerPlus,http://dx.doi.org/10.1186/2193-1801-3-254.
Nagarani,G.,Abirami,A.,Siddhuraju,P.,2014a.Foodprospectsandnutraceutical attributesofMomordicaspecies:apotentialtropicalbioresources–areview. FoodSci.Hum.Wellness3,117–126.
Nagarani,G.,Abirami,A.,Siddhuraju,P.,2014b.Acomparativestudyonantioxidant potentials,inhibitoryactivitiesagainstkeyenzymesrelatedtometabolic syn-drome,andanti-inflammatoryactivityofleafextractfromdifferentMomordica species.FoodSci.Hum.Wellness3,36–46.
Rios,J.,Escandell,J.,Recio,M.,2005.Newinsightsintobioactivityofcucurbitacins. In:Atta-ur-Rahman(Ed.),StudiesinNaturalProductsChemistry.Amsterdam, pp.429–469.
Singh,A.K.,Behera,T.K.,Chandel,D.,Sharma,P.,Singh,N.K.,2007.Assessinggenetic relationshipsamongbittergourd(MomordicacharantiaL.)accessionsusing inter-simplesequencerepeat(ISSR) markers.J.Hortic. Sci.Biotechnol. 82, 217–222.
Singh,J.,Cumming,E.,Manoharan,G.,Kalasz,H.,Adeghate,E.,2011.Suppl2: Medicinalchemistryoftheanti-diabeticeffectsofMomordicacharantia:active constituentsandmodesofactions.OpenMed.Chem.J.5,70.
Tan,S.P.,Parks,S.E.,Stathopoulos,C.E.,Roach,P.D.,2014.Extractionofflavonoids frombittermelon.FoodNutr.Sci.5,458–465.
Wu,S.J.,Ng,L.T.,2008.Antioxidantandfreeradicalscavengingactivitiesofwild bittermelon(MomordicacharantiaLinn.var.abbreviateSer.)inTaiwan.LWT– Food.Sci.Technol.41,323–330.
Zhou,H.,Tang, W.,Zeng,J.,Tang,C.,2014. Screeningofterpene lactonesand flavonoidglycosidesinGingkobilobacapsulebyUPLC-OrbitrapHighResolution MS,withemphasisonisomerdifferentiation.J.FoodNutr.Res.2,369–376.
Zhang,Q.C.,1992.PreliminaryreportontheuseofMomordicacharantiaextractby HIVpatients.J.Naturpath.Med.3,65–69.
Zhu,Y.,Dong,Y.,Qian,X.,Cui,F.,Guo,Q.,Zhou,X.,Wang,Y.,Zhang,Y.,Xiong,Z.,2012.