w ww.e l s e v i e r . c o m / l o c a t e / b j p
Original
Article
Antinociceptive
effect
of
semi-purified
petroleum
ether
partition
of
Muntingia
calabura
leaves
Zainul
Amiruddin
Zakaria
a,c,∗,
Mohd
Hijaz
Mohd
Sani
a,
Arifah
Abdul
Kadir
b,
Teh
Lay
Kek
c,d,
Mohd
Zaki
Salleh
c,daDepartmentofBiomedicalSciences,FacultyofMedicineandHealthScience,UniversitiPutraMalaysia,Serdang,Malaysia
bDepartmentofVeterinaryPreclinicalSciences,FacultyofVeterinaryMedicine,UniversitiPutraMalaysia,Serdang,Malaysia
cIntegrativePharmacogenomicsInstitute,UniversitiTeknologiMARA,Selangor,Malaysia
dFacultyofPharmacy,UniversitiTeknologiMARA,Selangor,Malaysia
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received23August2015 Accepted7December2015 Availableonline2April2016
Keywords:
Muntingiacalabura
Petroleumetherpartition Antinociceptiveactivity Mechanismsofantinociception
a
b
s
t
r
a
c
t
MuntingiacalaburaL.,Muntingiaceae,isamedicinalplantforvariouspain-relateddiseases.Theaimsof thepresentstudyweretodeterminetheantinociceptiveprofileandtoelucidatethepossible mech-anismsofantinociceptionofpetroleumetherpartitionobtainedfromcrudemethanolextractofM. calaburaleavesusingvariousanimalmodels.Theantinociceptiveprofileofpetroleumetherfraction (givenoral;100,250and500mg/kg)wasestablishedusingtheinvivochemicals(aceticacid-induced abdominalconstrictionandformalin-inducedpawlickingtest)andthermal(hotplatetest) models ofnociception.Theroleofglutamate,TRPV1receptor,bradykinin,proteinkinaseC,potassium chan-nels,andvariousopioidandnon-opioidreceptorsinmodulatingthepartition’santinociceptiveactivity wasalsodetermined.Theresultsobtaineddemonstratedthatpetroleumetherpartitionexerted signif-icant(p<0.05)antinociceptioninallthechemicals-,thermal-,capsaicin-,glutamate-,bradykinin,and phorbol12-myristate13-acetate(PMA)-inducednociceptionmodels.Theantinociceptiveactivitywas reversedfollowingpretreatmentwithopioidantagonists(i.e.naloxone,-funaltrexamine,naltrindole andnor-binaltorphimine),andthenon-opioidreceptorantagonists(i.e.pindolol(a-adrenoceptor), haloperidol(anon-selectivedopaminergic),atropine(anon-selectivecholinergicreceptor),caffeine(a non-selectiveadenosinergicreceptor),andyohimbine(an␣2-noradrenergic)).Inaddition,pretreatment withl-arginine(anitricoxide(NO)donor),NG-nitro-l-argininemethylesters(l-NAME;aninhibitorof
NOsynthase(NOS)),methyleneblue(MB;aninhibitorofcyclic-guanosinemonophosphate(cGMP) path-way),ortheircombinationfailedtoinhibitpetroleumetherpartition’santinociception.Inconclusion, petroleumetherpartitionexertsantinociceptiveactivityattheperipheralandcentrallevelsviathe mod-ulationof,partly,theopioid(i.e.,and␦)andseveralnon-opioids(i.e.-adrenergic,dopaminergic, cholinergic,adenosinergic,and␣2-noradrenergic)receptors,glutamatergic,TRPV1receptors,PKCand K+channelssystems,butnotl-arg/NO/cGMPpathway.
©2016SociedadeBrasileiradeFarmacognosia.PublishedbyElsevierEditoraLtda.Thisisanopen accessarticleundertheCCBY-NC-NDlicense(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Introduction
Variousneurotransmittersand receptorsystemstakepartin themodulationofpainprocessesinthecentralnervoussystem (CNS)andperipheralnervoussystem(PNS),suchasthevanilloid (Cui et al., 2006), opioid (Craig and Sorkin, 2011), glutamate (Osikowiczetal.,2013),proteinkinaseC(PKC;Velázquezetal., 2007),potassiumion(K+)channels(Ocanaetal.,2004),andnitric
oxide/cyclic guanosine monophosphate (NO/cGMP) pathways
∗ Correspondingauthor.
E-mail:[email protected](Z.A.Zakaria).
(Schmidtko etal.,2009).Despitetherapidadvancementsinthe developmentofvarioustreatmentsforpain,theclinicalefficacy andtolerabilityofconventionalanalgesicscanbeovershadowed byitsunwantedadverseeffects.Somepatientshavetaken alterna-tiveapproachtotreatpain,includingcomplementaryalternative medicine(CAM)inordertoavoidthesesideeffects(Astin,1998). Theuseofnaturalproductasalternativetherapiesisan increas-inglypopularmethodtotreatsuchdiscomfort,eitheraloneoras acomplementtoconventionalmedicalapproaches(Wirthetal., 2005).HoltandChandra(2002)categorized naturalproductsas herbs,herbalconcoctions,traditionalChinesemedicines,dietary supplements and alternative medicines. The research in natu-ral product based on its ethnopharmacological knowledge has
http://dx.doi.org/10.1016/j.bjp.2015.12.007
provided substantial contributions to drug innovation through the discovery of novel chemical structure and/or mechanism of actions (Rates, 2001). Muntingia calabura L., Muntingiaceae, the sole species in the genus, has been traditionally used in the Southeast Asia and tropical America (Kaneda et al., 1991; Nshimoet al., 1993)to treat headaches and gastric ulcer, and asan emmenogogue, antidyspeptic,antispasmodic, diaphoretic, tranquillizerandtonic(Kanedaetal.,1991;Perez-Arbelaez,1975). Various medicinal properties have been scientifically reported from M. calabura leaves, including anti-tumor (Kaneda et al., 1991;Suetal.,2003),hypotensive(Shihetal.,2006),antibacterial (Zakaria etal.,2006a,b;Zakariaetal.,2010;Sufianetal.,2013; Isnariantietal.,2013),antiplateletaggregation(Chenetal.,2007), anti-inflammatory,antipyreticandantinociceptive(Zakariaetal., 2007a,b), anti-myocardial infarction (Nivethetha et al., 2009), antihypertensive(Shih,2009),antioxidant(Siddiquaetal.,2010; Zakariaet al.,2011; ArunaSindhe etal., 2013)cytotoxic(Chen etal.,2005;Sufianetal., 2013), antiproliferative(Zakariaetal., 2011), antihyperglycemic (Aruna Sindhe et al., 2013), antiulcer (Balanetal.,2014;Zakariaetal.,2014a,b),andhepatoprotective (Mahmood et al., 2014) activities. Other parts of M. calabura have also been reported to exert insecticidal (Neto Bandeira et al., 2013), anti-inflammatory (Preethi et al., 2012; Gomathi et al., 2013), antioxidant (Preethi et al., 2010; Gomathi et al., 2013) activities. With regards to the antinociceptive studies, themethanolextractofM.calaburahasbeenreportedtoshow antinociceptive activity (Mohd Sani et al., 2012), which was later shown to involve activation of the non-selective opioid (i.e. -, ␦- and -opioid) and non-opioid (i.e. adenosinergic,
␣2-noradrenergic, and -adrenergic) receptors, modulation of the ATP-sensitive K+ channel, and inhibition of bradikinin and
proteinkinaseCactions(Zakariaet al.,2014a,b).Moreover,the authorsalsosuggestedthattheobservedantinociceptiveactivities anditsmechanismofactionswereresultedfromthesynergistic effectofthebioactivecompounds,flavonoids, saponins,tannins andsteroids, possessedbytheplantextracts.Recentlaboratory researchprovedthatthecrude methanolextractofM.calabura (MEMC)leavespossessesgoodtherapeuticeffectinreducing noci-ceptiveresponse(MohdSanietal.,2012;Zakariaetal.,2014a,b) andfurtherstudybyMohamad Yusofetal. (2013)leadstothe isolationoffourflavonoid-basedantinociceptive-bearingbioactive compounds,ofwhichoneisanewcompoundcalledcalaburone (8-hydroxy-6-methoxyflavone)and threewereknowncompounds, namely 5-hydroxy-3,7,8-trimethoxyflavone,3,7-dimethoxy-5-hydroflavone and 2′,4′-dihydroxy-3′-methoxychalcone. In this study,we furtherevaluate thepossible antinociceptiveactivity and its mechanism of action of the petroleum ether partition extractedfromMEMC.
Materialsandmethods
Plantcollection
TheleavesofMuntingiacalaburaL.,Muntingiaceae,were col-lectedfromitsnaturalhabitatinShahAlam,Selangor,Malaysia. Plantidentificationhasbeenmadeearlier(Balanetal.,2014)and avoucherspecimen(SK964/04)hasbeendepositedatthe Herbar-iumofInstituteofBiosciences,UniversitiPutraMalaysia,Serdang, Selangor,Malaysia.
PreparationofmethanolextractofM.calabura(MEMC)and itspetroleumetherfraction(PEMC)
ThemethodforpreparationofMEMCwasdescribedindetail byBalanet al.(2014). TheobtainedMEMCwasthen extracted
subsequentlywithpetroleumetherusingthemethoddescribed in detail by Mohamad Yusof et al. (2013). MEMC (2g) dis-solvedinasolventwitharatioof1:7:2whichconsistofMeOH (100ml),petroleumether (700ml)anddistilled water (200ml). Themixturewasrigorouslyshakenusingaseparatoryfunneland thenlefttoseparate.Thesupernatantwascollectedandfiltered whiletheresidue wassoakedagainwithpetroleumetheruntil thesupernatantbecomecolorless.Thecollectedpetroleumether supernatantwasalsosubjectedtorotaryevaporatorat40◦Cunder reduced pressuretoobtaina concentratedextractofpetroleum etherofM.calabura(PEMC).
Experimentalanimals
MaleICRmice(25–30g)andSpragueDawleyrat(150–180g) wereobtainedfroma privatesupplier (ChenurSupplier, Selan-gor, Malaysia) and acclimatized for three days in the Animal Holding Unit, Faculty of Medicine and Health Sciences, UPM. Details on the handled and cared of these animals have been described by Balan et al. (2014) and was in compliance with currentUPMguidelinesforthecareoflaboratoryanimals (Ethi-calapprovalno.:UPM/FPSK/PADS/BR-UUH/00404)andtheethical guidelinesforinvestigationsofexperimentalpaininconscious ani-mals(Zimmermann,1983).Allexperiments(n=6)wereconducted between09:00and16:00htominimizetheeffectofenvironmental changes.
Drugsandchemicals
Capsaicin,glutamate,naloxone,-funaltrexamine,naltrindole, nor-binaltorphimine, apamin, charybdotoxin, tetraethylammon-ium,atropine,haloperidol,pindolol,yohimbine,caffeine, gliben-clamide were purchased from Sigma–Aldrich (USA) while bradykininwaspurchasedfromTocrisBioscience,UK.Theywere preparedatthedesireddosebydissolvingthemindistilledwater (dH2O). Acetyl salicylic acid (ASA), morphine and capsezepine
(Sigma–Aldrich,USA)weredissolvedin10%DMSOwhilephorbol 12-myristate 13-acetate (PMA; Sigma–Aldrich, USA) was dis-solved in PBS solution.Acetic acid, petroleum ether, methanol anddimethylsulfoxide(DMSO)werepurchasedfromFisher Sci-entific (U.K.). All drugs and PEMC were given in a 10ml/kg basis.
Antinociceptiveanalysis
Aceticacid-inducedabdominalconstrictiontest
Theaceticacid-inducedabdominalconstrictionwascarriedout withslightmodifications accordingtothemethoddescribed in detailbyMohdSanietal.(2012).Themicewerepretreatedorally with10%DMSO(vehicle,negativecontrol),100mg/kgASA (posi-tivecontrol)orPEMC(100,250and500mg/kg)and60minlater subjectedtotheabdominalconstrictiontest.
Hotplatetest
Formalintest
Theformalintestwascarriedoutasdescribedindetailin previ-ousresearchbyMohdSanietal.(2012).Ratswereadministered orallywiththetestsolutions (i.e.10%DMSO(negativecontrol), 100mg/kgASA(peripherallyactingpositivecontrol),5mg/kg mor-phine(centrallyactingpositivecontrol)orPEMC (100,250and 500mg/kg))60minpriortotheformalininjection.The nocicep-tiveresponsedescribedastheearlyphaseandthelatephasewas recordedattherespective0–5minand15–30minfollowingthe formalinadministration.
Determinationofthemotorcoordinationandsedative effectsofPEMC
EffectofPEMConmotorcoordination
Rota-rodtest
Arota-rodtreadmilldevice(Ugo Basil,Italy)wasemployed toassesstheeffectofPEMConmotorcoordination(Meloetal., 2011)withslightmodifications. In thebeginning,micecapable ofremainingontherota-rodapparatuslongerthan210s(9rpm) wereselected12and24hpriortothetest.Thirtyminutesafterthe administrationofeither10%DMSO(negativecontrol),diazepam (2.5mg/kg, i.p.) or PECM (100, 250 and 500mg/kg, p.o.), each mousewastestedontherota-rodapparatusat0.5,1and2h post-treatment.Thetime(s)themicewereabletoremainontopof thebarwasrecordedforupto210s.Decreaseinfallofftimeis suggestiveofdepressionofthecentralnervoussystem(CNS).
SedativeeffectofPEMC
Sodiumpentobarbital-inducedsleepingtimetest
ThesedativeeffectofPEMCincombinationwithsodium pen-tobarbital(SP)wasevaluated(Martínez-Vázquezetal.,2012).For thispurpose,atotalof30miceweredividedintofivegroups(n=6) andorallyadministeredwitheither10%DMSO(negativecontrol), diazepam(2.5mg/kg,i.p.)orPECM(100,250and500mg/kg,p.o.) 30minbeforetheintraperitonealadministrationofsodium pento-barbital(42mg/kg,i.p.).Eachmousewasplacedonawarmtable andcautiouslymonitoredfortheonsetofuncoordinated move-mentssubsequenttothesedativephaseofthetest. Lossofthe rightingreflexrelatedtothehypnosisphaseandthedurationof sleepwerealsoobserved.Theeffectswererecordedasfollows: timepassedbetweentheadministrationsofpentobarbitaluntilloss ofrightingreflexwasrecordedasoftheonsetofsleep,whilstthe timefromthelosstorecoverywasconsideredasthedurationof sleep.
Investigationonthepossiblemechanismsofantinociceptionof PEMC
Capsaicin-inducedpawlickingtest
AntinociceptiveactionofPEMConvanilloidreceptors(TRPV1) were investigated using the proceduredescribed by Goncalves etal.(2005)withslightmodifications.Ratswerepretreatedorally withvehicle,capsazepine(0.17mmol/kg)orPEMC(100,250and 500mg/kg)60minbeforecapsaicininjection(1.6g/paw,50l) intothei.pl.regionof therat’srighthindpaw. Theinducedrat individuallyobservedinatransparentglasschamberfor5min,and theamountoftimetheanimalsspentlickingtheinjectedpawwas recordedwithachronometer.
Glutamate-inducedpawlickingtest
AslightmodificationfromtheproceduredescribedbyBeirith etal.(2002)wasusedtostudytheroleofglutamatergicsystem
inthemodulationofPEMCantinociceptiveaction.Ratswere pre-treatedorallywithvehicle,ASA(100mg/kg)orPEMC(100,250and 500mg/kg)60minpriortoglutamateinjection.Avolumeof50l ofglutamate(10mol/paw)wasinjectedintothei.pl.regionon therighthindpawofeachrat.Immediatelyafterthephlogistic agentadministration,theinducedratwereplacedinatransparent glasscageobservationchamberandindividuallyobservedfrom0 to15min.Theamountoftimetheanimalsspentlickingorbiting theinjectedpawwasrecordedwithachronometer.
InvolvementofproteinkinaseC
The experiment was conducted based on the previously describedmethodbySavegnagoetal.(2007).Avolumeof50l ofPMA(aproteinkinaseCactivator)solution(0.05g/paw)was injectedintotheventralsurfaceoftherighthindpawoftherat 60minaftertheoraladministrationofvehicle,ASA(100mg/kg)or PEMC(100,250and500mg/kg).Theanimalswereobserved indi-viduallyfromminute15to45following PMAinjectionandthe amountoftimetheratspentlickingtheinjectedpawwasrecorded usingachronometer.
Bradykinin-inducednociception
Based onthemethodpreviously described byFerreiraet al. (2004),bradykinin(10nmol/pawin50l)wasinjectedintothe intraplantarsurfaceoftherighthindpawofeachrat60minafter theoraladministrationofvehicle,ASA(100mg/kg)orPEMC(100, 250and 500mg/kg). The bradykinin-inducedrat was observed individuallyfor10min,andtheamountoftimetheyspentlicking theinjectedpawwasrecorded.
Involvementofl-arginine/nitricoxide/cyclicguanosine
monophosphatepathway
To determine the role of nitric oxide/cyclic-guanosine monophosphate (NO/cGMP) pathway in the modulation of PEMCantinociceptiveactivitythemethoddescribedindetailby
Zakariaetal.(2006a,b)wasadopted.Micewerepretreatedwith 20mg/kgl-arginine,l-NAME,MBortheirrespectivecombination
(l-argininewithl-NAMEorl-argininewithMB)followed5min
laterbypretreatmentwithvehicleorPEMC(500mg/kg), respec-tively.Sixtyminutesaftertheadministrationoftestsolutions,the miceweresubjectedtotheabdominalconstrictiontest.
Involvementofpotassiumchannels
Todeterminetheeffectof PEMC-inducedantinociception on K+channels,amethodpreviouslydescribedbyAlvesandDuarte (2002)wasused.Rats werepre-treated withglibenclamide(an ATPsensitiveK+channelinhibitor;10mg/kg,i.p.),apamin(small
conductanceCa2+-activatedK+channels,0.04mg/kg,i.p.),
charyb-dotoxin (an inhibitor of large conductance Ca2+-activated K+
channels,0.02mg/kg, i.p.)and tetraethylammonium chloride (a non-selective voltage dependant K+ channel inhibitor, 4mg/kg, i.p.)15minbeforeoraladministrationofeithervehicleorPEMC (500mg/kg).Sixtyminuteslater,painwasinducedusing0.6%acetic acid.Thenumberofwrithingwillberecordedfor25min,5min followingaceticacidinjection.
EffectofvariousreceptorantagonistsonPEMC-induced antinociception
pre-treatedwithcaffeine(3mg/kg,i.p.),atropine(10mg/kg,i.p.), haloperidol(20mg/kg,i.p.),pindolol(1mg/kg.i.p.),oryohimbine (0.15mg/kg,i.p)15minbeforetheoraladministrationofvehicleor PEMC(500mg/kg).Sixtyminuteslateraftertheadministrationof PEMCorvehicle,themiceweresubjectedtotheabdominal con-strictiontest.
Analysisofopioidreceptorsubtypes
Evaluationontheinvolvementofopioidreceptorsubtypewas doneusingtheabdominalconstrictiontestwhichissimilarwith previouslydescribed.Thepreparation oftheopioidantagonists’ dosesandtimingofadministrationwerebasedonpreviously con-ductedstudiesbyChoietal.(2003),Reetaetal.(2006)andZakaria etal.(2014a,b).Theopioidantagonist,-funaltraxamine(-FNA; 10mg/kg, i.p.), ␦opioid receptor antagonist, naltrindole (NALT; 1mg/kg.i.p.)oropioidreceptorantagonist,nor-binaltorphimine (nor-BNI; 1mg/kg, i.p.) were administered 90min, 15min and 30min respectively, beforeoral administration of500mg/kg of PEMC.Thenociceptivestimuluswasinjected60minafterPEMC administration.
HPLCprofileofPEMCatvariouswavelengths
The HPLCprofile of PEMC wasestablished according tothe methods described in detail for MEMC by Mohd Sani et al. (2012).Inaddition,PEMCwasalsospikedwithalistof flavonoid-based compounds, namely pinostrobin, hesperetin, flavanone, 4′,5,7-trihydroxyflavanone,2,4,4′-trihydroxychalcone,quercitrin, dihydroquercitin,fisetin,quercetin,rutin,quercitrin,naringenin, silibinin,andgenistein,todeterminetheirpresenceintheextract. The HPLC analysis was carried out in the Laboratory of Phy-tomedicine,MedicinalPlantsDivision,ForestResearchInstituteof Malaysia(FRIM),Kepong,Malaysia.
Statisticalanalyses
Theresultsare presented as mean±standard errorof mean (SEM).Theone-wayANOVAtestwithDunnett’smultiple compar-isonandtwo-wayANOVAwithBonferroniposthoctestwereused toanalyzeandcomparethedata.p<0.05wassetasthelimitof significance.
Result
AntinociceptiveprofileofPEMC
Fig.1 showsthat orallyadministered PEMCproduced a sig-nificant(p<0.001)anddose-dependentantinociceptiveeffectin micesubjectedtotheaceticacid-inducedabdominalconstriction test.Thepercentageanalgesiaproducedrangedbetween26and 70%,while thepositive controltreatment(100mg/kg ASA,p.o.) producedananalgesiceffectof59.31%.
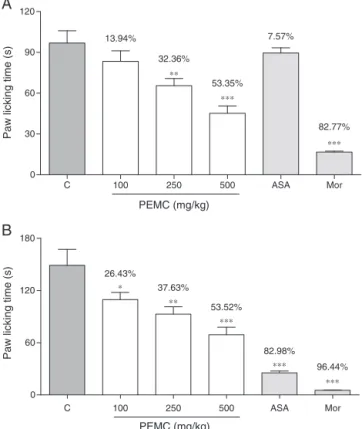
Fig.2showsthecentrallyandperipherallymediated antinoci-ceptiveprofileof orallyadministeredPEMCas indicatedbythe formalintest.Theextractexertedsignificant(p<0.05)and dose-dependentantinociceptiveactivityinboththeearlyandlatephases oftheassay.Theantinociceptiveactivity,inbothphases,waslower thanthatof5mg/kgmorphine.
ThecentralantinociceptiveprofileofPEMC,performedusing hotplatetest,athermal-inducednociceptionmodelwasshownin
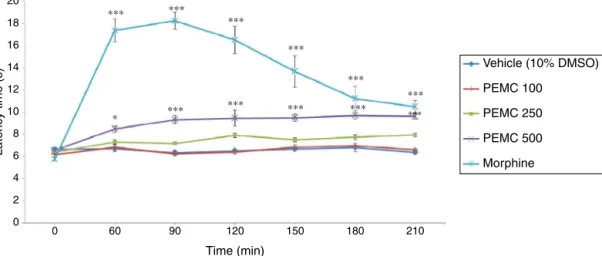
Fig.3.PEMCexertedsignificant(p<0.001)antinociceptiveactivity onlybythe500mg/kgwhichisthehighestdosetested.However, theantinociceptioninducedbyPEMCwasagainlowerthanthat inducedby5mg/kgmorphineadministeredviai.p.injection.
0 20 40 60 80 100
26.05% ∗∗∗
49.05% ∗∗∗
70.15% ∗∗∗
59.31% ∗∗∗
ASA 500 250 100 C
PEMC (mg/kg)
Number of constrictions
Fig.1.Numberofabdominalconstrictionsinmiceduringtheaceticacid-induced abdominalconstrictiontestaftertreatmentwithPEMC,acetylsalicylicacid(ASA; positivecontrol),orvehicle([C]negativecontrol).Aceticacidwasadministered intraperitoneally60minbeforeoraladministrationoftreatment.n=6ineachgroup. Numbersabovethebarsindicatethepercentageofanalgesia.***p<0.001as com-paredtonegativecontroltreatmentbyone-wayanalysisofvariancefollowedby Dunnett’sposthoctest.
MotorcoordinationandsedativeeffectsofPEMC
EffectofPEMConmotorcoordination
Intherota rodtest, alldoses ofPECMdidnotimpairmotor coordinationat0.5,1and2hpost-administrationincontrastto 2.5mg/kgdiazepam,whichcausedasignificant(p<0.05)decrease intimethattheanimalsstayedontherota-rodapparatuscompared tothecontrolgroup(Table1).
A
0 30 60 90 120
∗∗∗ ∗∗∗
∗∗
Mor 500
250 100
C ASA
PEMC (mg/kg)
13.94%
32.36%
53.35% 7.57%
82.77%
Paw licking time (s)
B
0 60 120 180
∗∗∗ ∗∗∗
∗∗
Mor 500
250 100
C ASA
∗∗∗ ∗
PEMC (mg/kg)
26.43%
37.63%
53.52%
82.98%
96.44%
Paw licking time (s)
Fig.2.EffectofPEMCinformalin-inducedpawlickingtest.Datarepresentsthe mean±SEMofsixrats.Theratswerepretreatedwithvehicle(10%DMSO),PEMC (100,250,and500mg/kg,p.o.),acetylsalicylicacid(ASA,p.o.),ormorphine(5mg/kg,
p.o.),60minbeforei.pl.injectionofformalin.Theasterisksdenotethesignificance
∗∗∗ ∗∗∗
∗∗∗
∗∗∗
∗∗∗
∗∗∗
∗∗∗ ∗∗∗ ∗∗∗ ∗∗∗
∗∗∗ ∗
20
18
16
14
12
10
8
Latency time (s)
Time (min) 6
4
2
0
0 60 90 120 150 180
Vehicle (10% DMSO)
PEMC 100
PEMC 250
PEMC 500
Morphine
210
Fig.3. AntinociceptiveprofileofPEMCassessedusingthehot-platetestinmice.*p<0.05whencomparedwiththecontrolgroupatsametherespectiveinterval.Dataare themean±SEM;n=6micepergroup.
Table1
EffectofPEMConmotorcoordinationassessedbytherota-rodtest.
Treatment Dose(mg/kg) Time(s)onRota-rod/210sa
30min 60min 90min
10%DMSO – 210.0±0.0 207.18±2.11 210.0±0.0
Diazepam 3 41.15±6.32a 28.18
±7.35a 56.92
±7.43a
PEMC
100 205.49±4.26 204.75±3.64 210.0±0.0
250 202.35±6.14 207.03±2.08 206.47±2.52
500 208.08±1.12 205.87±3.02 210.00±0.00
Dataarethemean±SEM;n=6micepergroup.Theratswerepretreatedwith10%DMSO(negativecontrol),2.5mg/kgdiazepam(i.p.)orPEMC(100,250,and500mg/kg, p.o.)30minbeforesubjectiontothetest.
aSignificantlydifferedatP<0.05whencomparedtothecontrolgroupasassessedbytheone-wayANOVAfollowedbyDunnett’sposthoctest.
Table2
EffectofPEMConmicesleepingactivityassessedusingthesodiumpentobarbital-inducedsleepingtest.
Treatment Dose(mg/kg) Onsetofsleep(min) Durationofsleep(min)
10%DMSO – 3.48±0.41 41.12±0.33
Diazepam 1 1.72±0.28a 94.33±0.66a
PEMC
100 3.08±0.17 68.66±0.47a
250 3.26±0.39 71.66±0.49a
500 2.98±0.32 88.83±0.55a
Dataarethemean±SEM;n=6micepergroup.
aSignificantlydifferedatP<0.05whencomparedtothecontrolgroupatthesamerespectiveintervalasassessedbytheone-wayANOVAfollowedbyDunnett’sposthoc test.
Sodiumpentobarbital-inducedsleepingtimetest
AdministrationofPEMCdidnotcausechangesinneitherthe latencysedationnorthesleepingtimeinducedbysodium pento-barbitalsuggestingthatPEMCdidnotexertsedativeeffectsatany ofthedosestested(Table2).
RoleofTRPV1,glutamatergic,B2receptorsandPKCpathway
TheabilityofPEMCtoinhibitthevanilloidreceptor-induced nociceptivetransmissionisshowninFig.4.The250and500mg/kg PEMC significantly (p<0.001) attenuated the capsaicin-induced pawlickingtestinadose-dependentmannerwiththepercentage ofanalgesiarecordedrangingbetween30and64%,respectively. Theinhibitionpercentageof63.52%producedby500mg/kgPEMC washigherthanthepositivecontrolcapsazepine(59.72%).
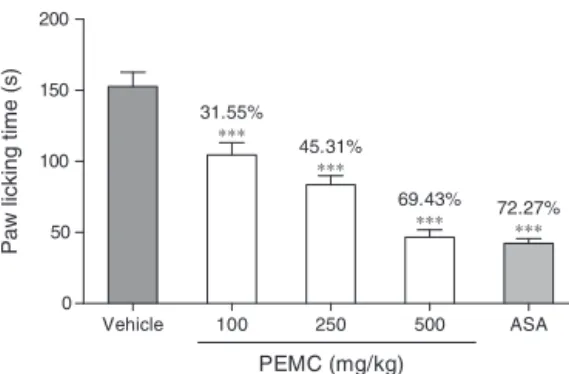
Fig. 5 shows the antinociceptive profile of PEMC against glutamate-induced paw licking test. PEMC significantly (p<0.001) attenuated the nociceptive effectassessed using the
0 20 40 60
Vehicle Capz
PEMC (mg/kg)
500 250 100
63.52% ∗∗∗
59.72% ∗∗∗ 30.74%
∗∗∗ 12.72%
Paw licking time (s)
0 50 100 150 200
Vehicle ASA
PEMC (mg/kg)
500 250 100
69.43% ∗∗∗ 72.27%∗∗∗ 45.31%
∗∗∗ 31.55%
∗∗∗
Paw licking time (s)
Fig.5.EffectofPEMConglutamate-inducedpawlickingtestinrats.Eachcolumn representsthemean±SEMofsixrats.Theratswerepretreatedwithvehicle (con-trol,10%DMSO)orPEMC(100,250,and500mg/kg,p.o.)60minbeforeinjectionof glutamate(10mol/paw,50l,i.pl.).Theasterisksdenotethesignificancelevelsas comparedtocontrol,***p<0.001byone-wayANOVAfollowedbyDunnett’spost hoctest.
glutamate-inducedpawlickingtestinadose-dependentmanner. Thepercentageofanalgesiaobtainedfromtheadministrationof 100,250and500mg/kgPEMCrangingbetween32and69%.
Fig.6showsthattheoraladministrationof250and500mg/kg PEMCproducedsignificant(p<0.001)inhibitionofPMA-induced nociception in rat. Interestingly, the 500mg/kg PEMC exerted slightlybetterantinociceptiveactivitycomparedto100mg/kgASA indicatedbythecalculatedpercentage ofanalgesia(66.38%and 55.36%respectively).
AsseeninFig.7,PEMCgivenorallyexhibitssignificant inhi-bitioninadose-dependentmanneronthenociceptioncausedby intra-plantarinjectionofbradykinininrat.Themaximalinhibition observedwas53.92%fordose500mg/kg.Similarinhibitoryeffect wasobservedfor100mg/kgaspirin.
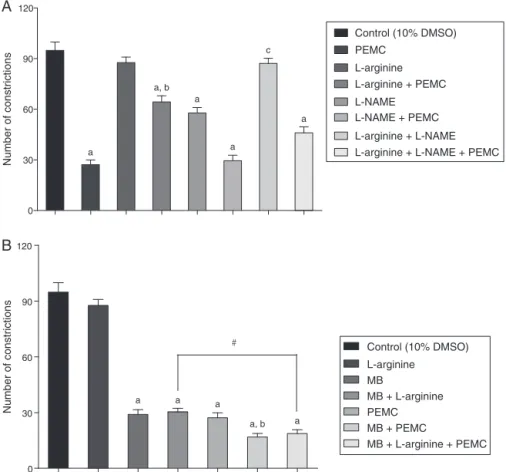
Evaluationofl-arginine/NO/cGMPpathway
Fig.8aandbshowtheroleoftheabdominalconstrictiontests incorporatingl-arginine,l-NAMEandMBtoexploretheroleofthe l-arginine/NO/cGMPpathwayinthemodulationofPEMC-induced
antinociceptiveactivity.InFig.8a,pretreatmentwith20mg/kgl
-argininedidnotproduceanysignificanteffectwhencomparedto thenegativecontrol.Meanwhile,pre-treatmentwith20mg/kgl
-NAMEaloneexertedasignificantantinociceptiveeffect.However, thepositiveeffectwasdiminishedwhenl-NAMEgivenin
combina-tionwithl-arginine.InthegroupstreatedwithPEMC,pretreatment
withl-NAMEdidnot affecttheextractantinociceptive activity,
whereaspretreatmentwithl-arginine significantlyreduced,but
0 20 40 60 80
ASA 500 250 100 Vehicle
PEMC (mg/kg)
36.52% ∗∗∗
66.38% ∗∗∗
55.36%
∗∗∗ 9.57%
Paw licking time (s)
Fig.6.EffectofPEMConnociceptioninducedbyphorbol12-myristate13-acetate (PMA)onrats.Eachcolumnrepresentsthemean±SEMtimespentonpaw-licking behaviorofsixanimals.Animalswereadministeredwithvehicle(negativecontrol), PEMC(100,250,and500mg/kg,p.o.)oracetylsalicylicacid(ASA;100mg/kg,p.o.) 60minbeforeinjectionofPMA(50mlsolutioncontaining0.05mgPMA/paw)inthe righthindpaw.Numbersabovebarsindicatepercentageofanalgesia.***p<.001 ascomparedtothecontrolgroupbyone-wayanalysisofvariancefollowedby Dunnett’sposthoctest.
0 20 40 60
Vehicle
PEMC (mg/kg) 500 250
100 ASA
23.82% ∗
53.92% ∗∗∗
51.1% ∗∗∗ 8.16%
Paw licking time (s)
Fig.7. TheantinociceptiveeffectofPEMCagainstbradykinin-inducedpawlicking. Eachcolumnrepresentsthemean±S.E.M.ofsixrats.Vehicle(negativecontrol; 10%DMSO,p.o.),PEMC(100,250and500mg/kg,p.o.)andacetylsalicylicacid(ASA: 100mg/kg,p.o.).***p<0.001whencomparedtocontrolgroup.
didnotcompletelyinhibit,theantinociceptiveeffectsofPEMC. Pre-treatmentwithacombinationofl-arginineandl-NAMEalsofailed
toaffecttheintensityofPEMCantinociception.
Pretreatment with 20mg/kg MB alone (Fig. 8b) exerted significant (p<0.05) antinociceptive activity and co-treatment with 20mg/kg l-arginine failed to reverse MB antinociception.
Pre-treatment with MB significantly (p<0.05) enhanced PEMC antinociceptionwhileco-treatmentwithMBandl-argininefailed
toaffecttheantinociceptiveintensityofPEMC.
Involvementofpotassiumchannels,non-opioidandopioid receptorsonPEMC-inducedantinociception
Fig. 9 shows the involvement of potassium channelsin the modulation of PEMC-induced antinociceptive activity. Pretreat-mentwithglibenclamide(10mg/kg,i.p.),Apamin(0.04mg/kg,i.p.), charybdotoxin(0.02mg/kg,i.p.)andtetraethylammoniumchloride (4mg/kg,i.p.),significantly(p<0.01)reversedtheantinociceptive activity of PEMC (500mg/kg) when assessed using acetic acid-inducedabdominalwrithingtest.
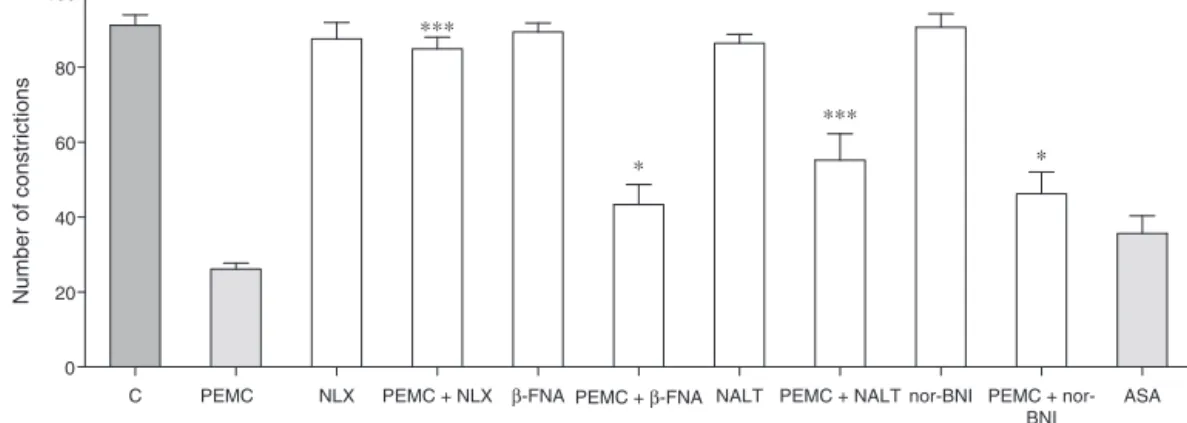
The effect of antinociceptive activity of PEMC was signifi-cantly (p<0.05) reversed following intraperitoneal administra-tion of several receptor antagonists (Fig. 10) namely caffeine (3mg/kg),atropine(10mg/kg),yohimbine(0.15mg/kg), haloperi-dol (0.2mg/kg), pindolol (0.2mg/kg), or naloxone, naltrindole, nor-binaltorphimineand-funaltrexamine(Fig.11).
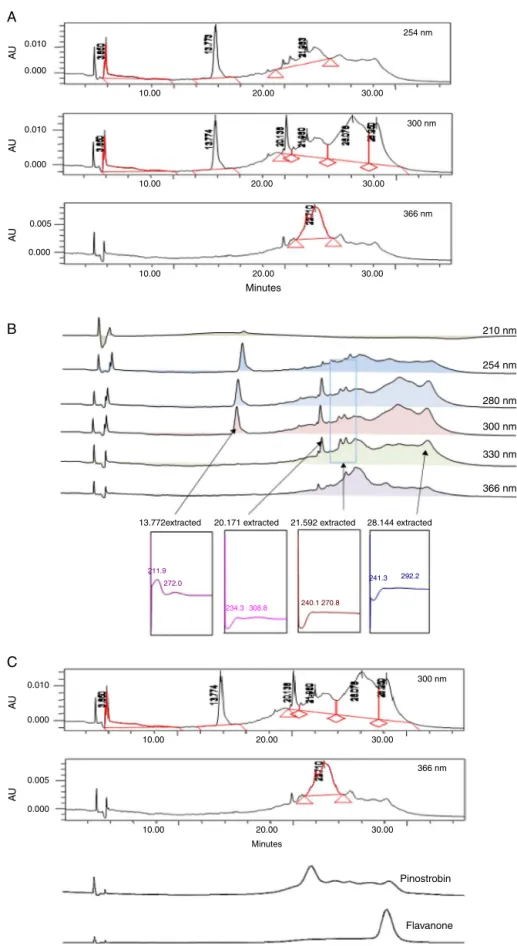
HPLCprofileofPEMC
TheHPLCanalysisofPEMCwasmeasuredatvariousdifferent wavelengths(e.g.210,254,280,300,330and366nmandthebest separationwasobtainedat300nm(Fig.12a).Fourmajorpeaks wereseparatedatthesewavelengths,whichwerelabeledasP1 (RT13.772min),P2(RT20.171min),P3(RT21.592min),andP4(RT
28.144min).Furtheranalysisdemonstratedthatthesefourpeaks showedmax values in theregionof 211.9–272.0, 234.3–308.8,
240.1–270.8,241.3–292.2nm,respectively(Fig.12b),suggesting, in part, the presence of flavonoid-based compounds. Compari-sonbetweenchromatogram ofPEMCand chromatogramof the standard compoundsdemonstrated thepresenceof pinostrobin andflavanone(Fig.12c).
Discussion
Thepresentstudywasthecontinuationofpreviousstudiesby
0 30 60 90 120
A
B
L-arginine + L-NAME Control (10% DMSO)
PEMC
L-arginine L-arginine + PEMC
L-NAME L-NAME + PEMC
L-arginine + L-NAME + PEMC
a
a, b a
a
a c
Number of constrictions
0 30 60 90 120
Control (10% DMSO)
PEMC MB L-arginine
MB + L-arginine
MB + PEMC
MB + L-arginine + PEMC
a
a a
a, b a #
Number of constrictions
Fig.8.(A)Effectsofl-arginine,l-NAME,andtheircombinationonPEMCantinociceptionasassessedbyaceticacid-inducedabdominalconstrictiontest.Thelettersrepresent thesignificancelevelsbetweencomparedgroups;ap<0.001whencomparedwithcontrolgroup;bp<0.001whencomparedwithPEMC-treatedgroup;cp<0.001when comparedwithl-NAME-treatedgroup,byone-wayANOVAfollowedbyDunnett’sposthoctest.(B)Effectsofl-arginine,methyleneblue,andtheircombinationonPEMC antinociceptionasassessedbyaceticacid-inducedabdominalconstrictiontest.Thelettersandsymbolabovethecolumnsrepresentsignificancelevelsbetweencompared groups;ap<0.001whencomparedwithcontrolgroup,bp<0.05whencomparedwithMB-treatedgroup,#p<0.05whencomparedbetweenselectedgroup.
ofM.calabura(MEMC).Recentlaboratorystudyconducted focus-ingontheantinociceptivepotential ofpartition extractedfrom theMEMC,wherethecrude extractpartitionedusingthree sol-ventwithdifferentpolaritynamelypetroleumether,ethylacetate andaqueoususingthemethoddescribedabove.Usingacetic acid-induced abdominal writhing test to screen for antinociceptive activity,petroleumether partitionproduced significantlybetter activity(datanotshown).Hence,thePEMCselectedtobefurther testedforitsantinociceptivepropertiesaswellasthemechanism ofactions.
OurdatashownthePEMCattenuatedthenociceptiveresponse inducedbychemicalandthermalstimuli,suggestingthatthe com-poundexertsperipheralandcentralantinociceptiveactivities.Its abilitytoinfluencebothperipheraland centralnociception was furtherconfirmedbyourobservationofitsantinociceptiveeffects in bothphases of theformalin test.Overall, theseobservations suggestthattheantinociceptiveactivityofPEMCresemblesthat ofopioid analgesics,suchas morphine.Moreover,theabilityof PEMCtoattenuatenociception inboth theabdominal constric-tionandhotplatetestssuggeststhat thecompoundiseffective
0 20 40 60 80 100
dH2O GLIB APA CHAR TEA dH2O GLIB APA CHAR TEA PEMC
Vehicle
∗∗ ∗
∗∗
Number of constrictions
0 20 40 60 80 100
V PE P P + PE H H + PE A A + PE C C + PE
a
a, b
a
a, b
Y + PE Y
a, b
a, b
Number of constrictions
Fig.10.Theinvolvementofseveralnon-opioidreceptorantagonistsinPEMC-inducedantinociceptionagainstaceticacid-inducedabdominalwrithingtestinmice.Vehicle (V:10%DMSO,p.o.),PEMC(PE:500mg/kg,p.o.),atropine(A:10mg/kg,i.p.)haloperidol(H:20mg/kg,i.p.),pindolol(P:1mg/kg,i.p.),caffeine(C:3mg/kg,i.p.),yohimbine(Y: 0.15mg/kg,i.p.).ap<0.001significantlydifferentwhencomparedtovehicle-treatedgroup;bp<0.001significantlydifferentwhencomparedtoPEMC-treatedgroup.
withbothinflammatory-inducedandnon-inflammatory-induced nociception,respectively(Zakariaetal.,2007a,b),whichalso sup-ported byourdata fromtheformalin-induced pawlicking test wherebothphases,theearly(neurogenic)andlate(inflammatory) phase,attenuatedbyPEMCtreatment.
Theaceticacid-inducedabdominalconstrictiontest,described asatypicalmodelforinflammatory pain,haslongbeenwidely usedasatooltoscreenforanalgesicoranti-inflammatory prop-ertiesof newagents(Collier etal.,1968; Vongtauet al.,2004). Theacetic acid-inducedabdominalconstriction testreflectsthe inflammatory-mediatednociceptiveresponse (MohdSani etal., 2012)where theinjected acetic acidinto theperitoneal cavity increases thelevel of cyclooxygenase(COX) and lipooxygenase (LOX)intheperitonealfluid andindirectly leadstotherelease ofendogenousmediatorssuchasprostaglandinE2(PGE2),PGF2␣,
serotonin,histamine,cytokines,andeicosanoids. These endoge-nousmediators,inturn,excitetheperipheralnociceptiveneurons withintheperitonealcavitythataresensitivetonon-steroidal anti-inflammatory drugs (NSAID) (Ikeda et al., 2001). Theincreased levelsofPGE2 duetoprolongirritationofperitonealcavitylead
toenhancedcapillarypermeability(Deraedtetal.,1980;Vogeland Vogel,1997)aswellasthereleaseofglutamateandsubstanceP fromperipheralafferentfiber(Millan,1999),indicatingthe stimu-lationofperipheralnociceptivemechanisms.TheabilityofPEMC toinhibitaceticacid-inducednociceptionsuggeststhatthe com-poundattenuates the levelsof peripheralCOXand LOX, which indirectlydecreasesPGE2 synthesisand obstructsthe
transduc-tionofpaininprimaryafferentnociceptors.However,theacetic
acid-inducedabdominalconstructiontestisnotspecific,thus fur-therstudiesusingothernociceptivemodels,namelythehotplate andformalintestsarerequiredtodeterminewhetherthe antinoci-ceptiveeffectsofPEMCarecentrallyorperipherallymediated.
The hot plate test, predominantly a spinal reflex test that involvesactivationofthesupraspinalnociceptiveprocessing,has beenappliedin variousresearches toshowthat onlythe anal-gesicactingcentrally,butnotperipherally,haveaneffectonthis test (Chen et al.,1995; Le Barset al., 2001; Hosseinzadeh and Younesi,2002;Giglioetal.,2006).TheabilityofPEMCtoinhibit boththeearlyandlatephasesoftheformalintestinthepresent studyconfirmedtheextract’sabilitytoexertcentrallyand periph-erally mediatedantinociceptive effects(MohdSani etal.,2012; MohamadYusofetal.,2013).Theearlyphase,classifiedasa neuro-genicpain,isanacutereaction,lastedfor5minwhichimmediately occuraftertheinjectionofformalinwhereitdirectlyactivatethe nociceptors.Thelatephase,appearsbetween15and30min fol-lowingformalininjection,isclassifiedasaninflammatorypainas itiscausedbyinflammatorymediatorsreleasedsuchashistamine, serotonin,bradykininandPGE2 (Vermaetal.,2005).Ourresults
inthepresentstudyindicatethatPEMCpossessesmildcentrally mediatedantinociceptivecharacteristicsasseeninboth thehot platetestandearlyphaseofformalintest.
Overthepastdecade,tremendousresearchbeingdevotedin developing thebestmedicationforpain reliever,and extensive studieshasbeenperformtoexploretheirprecisemechanismof actioninordertounderstandandlimitanyunwantedsideeffect. However,theneuronalbasisofpaintransmissionandmodulation
0 20 40 60 80 100
PEMC + NALT NALT
PEMC
C β-FNA PEMC + β-FNA PEMC + nor- ASA
BNI nor-BNI
∗∗∗
∗
PEMC + NLX NLX
∗∗∗
∗
Number of constrictions
AU
AU
AU
AU
AU
0.010
A
B
C
254 nm
300 nm
366 nm
300 nm
366 nm 0.000
0.010
0.000
10.00
10.00
10.00 0.010
0.005
0.000 0.000
20.00
Minutes
30.00
10.00 20.00 30.00
20.00
Minutes
13.772extracted
211.9
272.0
234.3 308.8
240.1270.8
Pinostrobin
Flavanone
241.3 292.2
20.171 extracted 21.592 extracted 28.144 extracted
210 nm
254 nm
280 nm
300 nm
330 nm
366 nm 30.00
10.00 20.00 30.00
20.00 30.00
0.005
0.000
Kv channel
Nociceptor terminal
Nitric oxide + L-citrulline
NOS PKC Post-synaptic nerve cell
Noxious stimulus
5-HTR
5-HTR
Opioid receptor
TRPV1
L-arginine
AMPA
NMDA SP
NO
PGE2 EP1
A1R
Glut
Ca2+
Ca2+
Ca2+
α2R
A1R
PGE2
B2
Primary afferent fiber
Opioid receptor KATP channel
sK Ca channel
Fig.13.SchematicdiagramoftheproposedmechanismsofantinociceptiveactiondemonstratedbyPEMC.PEMCexertsantinociceptiveactivityatperipheralandcentral levelsviamodulationofseveralreceptorsandneurotransmitterssystems.Ca2+:calciumion;cGMP:cyclicguanosinemonophosphate;GC:guanylatecyclase;GTP:guanosine triphosphate;Kv:voltage-gatedpotassiumchannel;KATP:ATP-sensitivepotassiumchannel;SKCa:smallconductancecalcium-activatedpotassiumchannel;5-HTR: sero-tonergicreceptor;␣2R:␣2-adrenergicreceptor;A1R:adenosinereceptor;Glut:glutamate;PGE2:prostaglandinE2;EP1:prostaglandinreceptor;B2:bradykininreceptor; NMDA:N-methyl-d-aspartate;PKC:proteinkinaseCenzyme;NOS:nitricoxidesynthase;NO:nitricoxide;TRPV1:transientreceptorpotentialcationchannelsubfamilyV member1.
isstillnotcompletelyunderstood.Numerouschemicalsandfactors suchasopioids,substanceP,glutamate,nitricoxide,serotonin,and otherinflammatorymediators,hasbeendocumentedtoengaged intheactivationofthePNSandCNSnociceptivepathways(Mohd Sanietal.,2014).Theexistencedescriptionofthesemodulatory mechanismssuchastheroleofvanilloidreceptors, glutamater-gicsystem,opioidandnon-opioidreceptors,nitricoxide,PKCas wellasK+channelspathways,havebenefitedandprovideguidance
forresearchersinthedevelopmentofnewanalgesiccandidates (Rodellaetal.,2010).
Our previous study (Mohd Sani et al., 2012; Zakaria et al., 2014a,b)usingthecrudemethanolextractofM.calabura(MEMC) revealedtheparticipationof mostthemodulatorymechanisms. Inthepresentstudy,weemployedthesameapproachtoexplore thepossiblemechanismsof actionresponsiblefortheobserved antinociceptiveactivityofPEMC.Basedonourfindings,PEMC;(1) attenuatethenociceptioninducedbycapsaicinindicatingpossible involvementofvanilloidreceptors,(2)inhibitglutamate-induced nociceptionwhichinturninhibitthereleaseofNOandNO-related substance (Beirith et al., 2002), (3) significantly inhibit PMA-inducednociceptionsuggestingitsinvolvementinPKC-activated phosphorylationofvanilloidreceptorpathway(Vellanietal.,2001), (4)inhibitbradykinin-inducednociceptionshowingitsabilityto blockB2receptorfrombeingactivated,(5)affectedbyl-arginine,
but not l-NAME and MB,suggesting PEMC antinociception did
notinvolvemodulationviathel-arginine/NO/cGMPpathway,(6)
involved in the activation of ATP-sensitive K+ channels, small
conductanceCa2+-activatedK+channelsandnon-selective
voltage-dependantK+channels,butnotlargeconductanceCa2+-activated
K+channels,(7)playsignificantroleinantinociceptionmodulated
byadenosinergic,␣2-noradrenergic,cholinergicand-adrenergic
receptors,(8)involvedintheactivationofopioidreceptorsubtypes asPEMCantinociceptiveeffectwassignificantlyreducedbythe pretreatmentof,␦andopioidreceptorsubtypeinhibitors.
Ourfindings,apartfromthepossibleparticipationof choliner-gicreceptorandinabilityofPEMCtoactivatelargeconductance Ca2+-activatedK+channels,wereinlinewithourresultsobtained
fromthecrudemethanolextractofM.calabura.Theinvolvementof cholinergicreceptorseeninPEMC-inducedantinociceptivemight probably due tohigher purity of theexpressed bioactive com-poundsextractedfromthecrudeextract.
EarlierstudiesimpliedthatthedepressionofCNSandtheeffect of nonspecific muscle relaxation can diminish the response of motorcoordinationand,accordingly,maynullifythe antinocicep-tiveresults(Quintans-Júnioretal.,2010).Interestingly,thepresent findingsdemonstratedthatPEMC-treated(100,250and500mg/kg, orally)miceexertednoperformancealterationsinthe rota-rod-andpentobarbital-inducedsleeping-testindicatingthatPEMCdid notinterfereinthemiceanxietybehaviororinthemotorfunction, whencomparedtothecontrolgroup.Incontrast,diazepaminduced anxietyreductiondemonstratedbyanincreaseinthenumberof visitsandtimespentintheopenarmsandbyanincreaseinthetotal numberofentries.Overall,thesefindingssuggestedthatthereisno alterationinmotorcoordinationamongthetreatedanimals, there-fore,eliminatinganonspecificmusclerelaxationaswellassedative effectsofPEMCatthedosesused.Theseresultssuggestthatthe antinociceptiveeffectofPEMCisnotrelatedtosedativeeffectorto alterationinmotorfunction(deVasconcelosetal.,2011).
Furtheranalysisoftheidentifiedpeaksbasedontherecorded UVspectrarevealedthatthefourpeaksfallsbetweenthemax
of 210and 310nm suggesting thepresence of flavonoid-based bioactivecompoundsasreportedbyAznametal.(2012).There arefivemajorsubgroupsofflavonoidsnamelyflavones,flavonols, flavanonols,flavanonesanddihydroflavonols(Tsimogiannisetal., 2007).Thepresenceofflavanones-basedbioactivecompounds,in particular,canbesuggestedbasedontherecordedUV–Visspectra obtainedforeachofthepeak(Aznametal.,2012).Accordingto
Aznametal.(2012),severalflavonoidsthathave methoxyland hydroxylgroups,detectedattheUVspectrathatrangebetween 210and290,havebeenisolatedfromfractionsofmethanolextract ofKaempferiarotunda. Recentstudiesby MohamadYusofet al. (2013)tendtosupportthepresenceofflavanonesor methoxyl-andhydroxyl-containedflavonoidsinPEMCastheauthorshave successfully isolatedseveralantinociceptive-bearing flavanones, namely 5-hydroxy-3,7,8-trimethoxyflavone, 3,7-dimethoxy-5-hydroyflavone, 2′,4′-dihydroxy-3′-methoxychalcone, and 8-hydroxy-6-methoxyflavone(calaburone).
Itisreasonabletoconcludefromourfindingsinthepresent studythattheantinociceptivepropertiesofPEMCoccurvia interac-tionofavarietyofphysiologicalpathwayswithinthePNSandCNS. PEMCappearstoactasanagonistforpotassiumchannels,opioid, cholinergic,adenosinergic,adrenergic,andserotonergicreceptors, whileatthesametimeinhibitingthePKC,vanilloidreceptorsand glutamatergicsystem.Fig.13representsamodelofthe mecha-nismsof PEMC’santinociceptiveactionbased onthesefindings. Theseactivitiescouldbeduetothesynergisticeffectofflavonoids, triterpenes,tanninsandsteroids.Theknowledgegainedfromsuch studiesinthepresentandfuturewillbeusefulforthedesignor dis-coveryofnewchemicalentitiesthatexhibitsimilarantinociceptive activity.
Ethicaldisclosures
Protectionofhumanandanimalsubjects. Theauthorsdeclare thattheproceduresfollowedwereinaccordancewiththe regula-tionsoftherelevantclinicalresearchethicscommitteeandwith thoseoftheCodeofEthicsoftheWorldMedicalAssociation (Dec-larationofHelsinki).
Confidentialityofdata. Theauthorsdeclarethatnopatientdata appearinthisarticle.
Righttoprivacyandinformedconsent. Theauthorsdeclarethat nopatientdataappearinthisarticle.
Authors’contribution
ZAZconceivedofthestudy,participatedinitsdesignandhelped todraftthemanuscript.Allauthorsreadandapprovedthefinal manuscript.MHMScarriedouttheexperimentsanddraftedthe manuscript. AKA participated in the experimental design, and helpedtodraftthemanuscript.TLKandMZSinvolvedinthe statis-ticalanalysisandmanuscriptpreparation.
Conflictsofinterest
Theauthorsdeclarenoconflictsofinterest.
Acknowledgements
This study was supported by the Science Fund Research Grant (Reference no. 06-01-04-SF1127) awarded by the Min-istryofScienceTechnologyandInnovation(MOSTI),Malaysiaand
theResearch UniversityGrantScheme(Referenceno. 04-02-12-2019RU)fromtheUniversitiPutraMalaysia,Malaysia.Theauthors thankedtheFaculty ofMedicineandHealthSciences,Universiti PutraMalaysia,Malaysiaforproviding thefacilitiestocarryout thisstudy.
References
Alves,D.,Duarte,I.,2002.InvolvementofATP-sensitiveK(+)channelsinthe periph-eralantinociceptiveeffectinducedbydipyrone.Eur.J.Pharmacol.444,47–52. ArunaSindhe,M.,Bodke,Y.D.,Chandrashekar,A.,2013.Antioxidantandinvivo
anti-hyperglycemicactivityofMuntingiacalaburaleavesextracts.DerPharmacia Lettre5,427–435.
Astin,J.A.,1998.Whypatientsusealternativemedicine:resultsofanationalstudy. J.Am.Med.Assoc.279,1548–1553.
Aznam,N.,Atun,S.,Arianingrum,R.,Nurestri,S.,2012.Isolation,identificationand antiviralactivityofbioactivecompoundsofKampheriarotunda.Int.Proc.Chem. Biol.Environ.Eng.38,27–30.
Balan,T.,MohdSani,M.H.,Suppaiah,V.,Mohtarrudin,N.,Suhaili,Z.,Ahmad,Z., Zakaria,Z.A.,2014.AntiulceractivityofMuntingiacalaburaleavesandthe pos-siblemechanismsofactioninvolved.Pharm.Biol.52,410–418.
Beirith,A.,Santos,A.R.S.,Calixto,J.B.,2002.Mechanismsunderlyingthenociception andpawoedemacausedbyinjectionofglutamateintothemousepaw.Brain Res.924,219–228.
Beirith,A.,Santos,A.R.S., Calixto,J.B.,Hess,S.C.,Messana,I.,Ferrari,F.,Yunes, R.A.,1999.Studyoftheantinociceptiveactionoftheethanolicextractand thetriterpene24-hydroxytormenticacidisolatedfromthestembarkofOcotea suaveolens.PlantaMed.65,50–55.
Chen,J.J.,Lee,H.H.,Duh,C.Y.,Chen,I.S.,2005.Cytotoxicchalconesandflavonoids fromtheleavesofMuntingiacalabura.PlantaMed.71,970–973.
Chen,J.J.,Lee,H.H.,Shih,C.D.,Liao,C.H.,Chen,I.S.,Chou,H.T.,2007.New dihydrochal-conesandanti-plateletaggregationconstituentsfromtheleavesofMuntingia calabura.PlantaMed.73,572–577.
Chen,Y.F.,Tsai,H.Y.,Wu,T.S.,1995.Anti-inflammatoryandanalgesicactivitiesfrom rootsofAngelicapubescens.PlantaMed.61,2–8.
Choi,S.S.,Han,K.J.,Lee,H.K.,Han,E.J.,Suh,H.W.,2003.Possibleantinociceptive mechanismsofopioidreceptorantagonistsinthemouseformalintest. Pharma-col.Biochem.Behav.75,121–124.
Collier,H.O.J.,Dinneen,J.C.,Johnson,C.A.,Schneider,C.,1968.Theabdominal con-strictionresponseanditssuppressionbyanalgesicdrugsinthemouse.Br.J. Pharmacol.Chemother.32,295–310.
Cui,M.,Honore,P.,Zhong,C.,Gauvin,D.,Mikusa,J.,Hernandez,G.,Faltynek,C.R., 2006.TRPV1receptorsintheCNSplayakeyroleinbroad-spectrumanalgesiaof TRPV1antagonists.J.Neurosci.26,9385–9393.
Craig,A.D.,Sorkin,L.S.,2011.Painandanalgesia.In:EncyclopediaofLifeSciences. JohnWiley, Chichester, England, http://dx.doi.org/10.1002/9780470015902. a0000275.pub3.
deVasconcelos,D.I.B.,Leite,J.A.,Carneiro,L.T.,Piuvezam,M.R.,deLima,M.R.V., de Morais, L.C.L., Rumjanek,V.M., Rodrigues-Mascarenhas, S., 2011. Anti-inflammatoryandantinociceptiveactivityofouabaininmice.Med.Inflamm. 2011,1–11,http://dx.doi.org/10.1155/2011/912925.
Deraedt,R.,Jouquey,S.,Delevallee,F.,Flahaut,M.,1980.Releaseofprostaglandins EandFinanalgogenicreactionanditsinhibition. Eur.J.Pharmacol.61, 17–24.
Ferreira,J.,daSilva,G.L.,Calixto,J.B.,2004.Contributionofvanilloidreceptorsto theovertnociceptioninducedbyB2kininreceptoractivationinmice.Br.J. Pharmacol.141,787–794.
Giglio,C.A.,Defino,H.L.A.,da-Silva,C.A.,de-Souza,A.S.,delBel,E.A.,2006. Behav-ioralandphysiologicalmethodsforearlyquantitativeassessmentofspinalcord injuryandprognosisinrats.Braz.J.Med.Biol.Res.39,1613–1623.
Gomathi,R.,Anusuya,N.,Manian,S.,2013.Adietaryantioxidantsupplementation ofJamaicancherries(MuntingiacalaburaL.)attenuatesinflammatoryrelated disorders.FoodSci.Biotechnol.22,787–794.
Goncalves,C.E.,Araldi,D.,Panatieri,R.B.,Rocha,J.B.,Zeni,G.,Nogueira,C.W.,2005. Antinociceptivepropertiesofacetylenicthiopheneandfuranderivatives: evi-denceforthemechanismofaction.LifeSci.76,2221–2234.
Holt,G.A.,Chandra,A.,2002.Herbsinthemodernhealthcareenvironment:an overviewofuses,legalitiesandtheroleofthehealthcareprofessional.Clin.Res. Regul.Aff.19,83–107.
Hosseinzadeh,H., Younesi,H.M.,2002. Antinociceptiveandanti-inflammatory effectsofCrocussativusL.stigmaandpetalextractsinmice.BMCPharmacol. 2,7,http://dx.doi.org/10.1186/1471-2210-2-7.
Ikeda,Y.,Ueno,A.,Naraba,H.,Oh-Ishi,S.,2001.Involvementofvanilloidreceptor VR1andprostanoidsintheacid-inducedwrithingresponsesofmice.LifeSci. 69,2911–2919.
Isnarianti,R.,Wahyudi,I.A.,Puspita,R.M.,2013.MuntingiacalaburaL.leavesextract inhibitsglucosyltransferaseactivityofStreptococcusmutans.J.Dent.Indonesia 20,59–63.
Karumi,Y.,Onyeyili,P.,Ogugbuaja,V.O.,2003.Anti-inflammatoryand antinocicep-tive(analgesic)propertiesofMomordicalbalsaminaLinn.(Balsamapple)leaves inrats.Pak.J.Biol.Sci.6,1515–1518.
LeBars,D.,Gozariu,M.,Cadden,S.W.,2001.Animalmodelsofnociception. Pharma-col.Rev.53,597–652.
Mahmood, D., Mamat, S.S., Kamisan, F.H., Yahya, F., Kamarolzaman, M.F.F., Nasir, N., Suhaili, Z., Mohtarrudin, N., Zakaria, Z.A., 2014. Amelioration of paracetamol-induced hepatotoxicity in rat by the administration of methanolextract of Muntingiacalabura L. leaves.Biomed. Res. Int. 2014, 1–10,http://dx.doi.org/10.1155/2014/695678.
Martínez-Vázquez, M., Estrada-Reyes, R., Martínez-Laurrabaquio, A., López-Rubalcava,C.,Heinze,G.,2012.NeuropharmacologicalstudyofDracocephalum moldovicaL.(Lamiaceae)inmice:sedativeeffectandchemicalanalysisofan aqueousextract.J.Ethnopharmacol.141,908–917.
Melo, M.S.,Santana, M.T., Guimarães,A.G., Siqueira,R.S., Sousa, D.P., Santos, M.R.V.,Bonjardim,L.R.,Araújo,A.A.S.,Onofre,A.S.C.,Lima,J.T.,Almeida,J.R.G.S., Quintans-Júnior,L.J.,2011.Bioassay-guidedevaluationofcentralnervous sys-temeffectsofcitronellalinrodents.Braz.J.Pharmacogn.21,697–703. Millan,M.J.,1999.Theinductionofpain:anintegrativereview.Prog.Neurobiol.57,
1–164.
MohamadYusof,M.I.,Salleh,M.Z.,Teh,L.K.,Ahmat,N.,NikAzmin,N.F.,Zakaria,Z.A., 2013.Activity-guidedisolationofbioactiveconstituentswithantinociceptive activityfromMuntingiacalaburaL.leavesusingtheformalintest.Evid.Based Complement.Altern.Med.,715074,http://dx.doi.org/10.1155/2013/715074. MohdSani,M.H.,Zakaria,Z.A.,Balan,T.,Teh,L.K.,Salleh,M.Z.,2012.Antinociceptive
activityofmethanol extractofMuntingiacalabura leavesandthe mecha-nismsofactioninvolved.Evid.BasedComplement.Altern.Med.2012,890361, http://dx.doi.org/10.1155/2012/890361.
MohdSani,M.H.,Taher,M.,Susanti,D.,The,L.K.,Salleh,M.Z.,Zakaria,Z.A.,2014. Mechanismsof␣-mangostin-inducedantinociceptioninarodentmodel.Biol. Res.Nurs.17,68–77.
Musa,A.M.,Aliyu,A.B.,Yaro,A.H.,Magaji,M.G.,Hassan,H.S.,Abdullahi,M.I.,2009. Preliminaryphytochemical,analgesicandanti-inflammatorystudiesofthe methanolextractofAnisopusmanniiinrodents.Afr.J.Pharm.Pharmacol.3, 374–378.
NetoBandeira,G.,daCamara,C.A.G.,deMoraes,M.M.,Barros,R.,Muhammad,S., Akhtar,Y.,2013.InsecticidalactivityofMuntingiacalaburaextractsagainst lar-vaeandpupaeofdiamondback,Plutellaxylostella(Lepidoptera,Plutellidae).J. KingSaudUniv.25,83–89.
Nivethetha, M., Jayasari, J., Brindha, P., 2009. Effects of Muntingia calabura L. on isoproterenol-induced myocardial infarction. Singapore Med. J. 50, 300–302.
Nshimo,C.M.,Pezzuto,J.M.,Kinghorn,A.D.,Farnsworth,N.R.,1993.Cytotoxic con-stituentsofMuntingiacalaburaleavesandstemscollectedinThailand.Int.J. Pharmacogn.31,77–81.
Ocana,M.,Cendan,C.M.,Cobos,E.J.,Entrena,J.M.,Baeyens,J.M.,2004.Potassium channelsandpain:presentrealitiesandfutureopportunities.Eur.J.Pharmacol. 500,203–219.
Osikowicz,M.,Mika,J.,Przewlocka,B.,2013.Theglutamatergicsystemasatarget forneuropathicpainrelief.Exp.Physiol.98,372–384.
Patel,N.K.,Bhutani,K.K.,2014.PinostrobinandcajanuslactoneisolatedfromCajanus cajan(L.)leavesinhibitsTNF-␣andIL-1production:invitroandinvivo exper-imentation.Phytomed.21,946–953.
Perez-Arbelaez,E.,1975.In:Salazar,H.(Ed.),PlantasMedicinalesyVenenosasde Colombia.,p.192,Medellin,Colombia.
Preethi,K.,Vijayalakshmi,N.,Shamna,R.,Sasikumar,J.R.,2010.Invitroantioxidant activityofextractsfromfruitsofMuntingiacalaburaLinn.fromIndia. Pharma-cogn.J.14,11–18.
Preethi,K.,Premasudha, P.,Keerthana, K.,2012. Anti-inflammatoryactivityof Muntingiacalaburafruits.Pharmacogn.J.4,51–56.
Quintans-Júnior,L.J.,Silva,D.A.,Siqueira,J.S.,Araújo,A.A.S.,Barreto,R.S.S., Bon-jardim,L.R., DeSantana,J.M.,DeLuccaJúnior,W.,Souza,M.F.V., Gutierrez, S.J.C.,Barbosa-Filho,J.M.,Santana-Filho,V.J.,Araújo,D.A.M.,Almeida,R.N.,2010. Bioassay-guidedevaluationofantinociceptiveeffectofN-salicyloyltryptamine: abehavioralandelectrophysiologicalapproach.J.Biomed.Biotechnol.2010, 1–6,http://dx.doi.org/10.1155/2010/230745.
Rates,S.M.K.,2001.Plantsassourceofdrugs.Toxicon29,603–613.
Reeta,K.,Mediratta,P.K.,Rathi,N.,Jain,H.,Chugh,C.,Sharma,K.K.,2006.Roleof kappaanddeltaopioidreceptorsintheantinociceptiveeffectofoxytocinin formalin-inducedpainresponseinmice.Regul.Pept.135,85–90.
Rodella,L.F.,Borsani,E.,Rezzani,R.,2010.Newperspectiveinpaintreatment.Open Conf.Proc.J.1,61–65.
Savegnago,L.,Pinto,L.G.,Jesse,C.R.,Alves,D.,Rocha,J.B.T.,Noguira,C.W.,Zeni, G.,2007.Antinociceptivepropertiesofdiphenyldiselenide:evidencesforthe mechanismofaction.Eur.J.Pharmacol.555,129–138.
Schmidtko,A.,Tegeder,I.,Geisslinger,G.,2009.NoNO,nopain?Theroleofnitric oxideandcGMPinspinalpainprocessing.TrendsNeurosci.32,339–346. Shih,C.D.,2009.Activationofnitricoxide/cGMP/PKGsignalingcascademediates
antihypertensiveeffectsofMuntingiacalaburainanesthetizedspontaneously hypertensiverats.Am.J.Chin.Med.37,1045–1058.
Shih,C.D.,Chen,J.J.,Lee,H.H.,2006.Activationofnitricoxidesignalingpathway mediateshypotensiveeffectofMuntingiacalaburaL.(Tiliaceae)leafextract.Am. J.Chin.Med.34,857–872.
Siddiqua,A.,Premakumari,K.B.,Sultana,R.,Savitha,V.,2010.Antioxidantactivity andestimationoftotalphenoliccontentofMuntingiacalaburabycalorimetry. Int.J.ChemTech.Res.2,205–208.
Starec,M.,Waitzov’a,D.,Elis,J.,1988.Evaluationoftheanalgesiceffectof RG-tanninusingthe“hotplate”and“tailflick”methodinmice.Cesk.Farm.37, 319–321.
Sufian,A.S.,Ramasamy,K.,Ahmat,N.,Zakaria,Z.A.,Mohd.Yusof,M.I.,2013.Isolation andidentificationofantibacterialandcytotoxiccompoundsfromtheleavesof MuntingiacalaburaL.J.Ethnopharmacol.146,198–204.
Su,B.N.,JungPark,E.,Vigo,J.S.,Graham,J.G.,Cabieses,F.,Fong,H.H.,Pezzuto,J.M., Kinghorn,A.D.,2003.Activity-guidedisolationofthechemicalconstituentsof Muntingiacalaburausingaquinonereductaseinductionassay.Phytochemistry 63,335–341.
Tsimogiannis,D.,Samiotaki,M.,Panayotou,G.,Oreopoulou,V.,2007. Character-ization offlavonoid subgroupsandhydroxy substitution byHPLC-MS/MS. Molecules12,593–606.
Tuchinda,P.,Reutrakul,V.,Claeson,P.,Pongprayoon,U.,Sematong,T.,Santisuk, T.,Taylor,W.,2002.Anti-inflammatorycyclohexenylchalconederivativesin Boesenbergiapandurata.Phytochemistry59,169–173.
Velázquez,K.T.,Mohammad,H.,Sweitzer,S.M.,2007.ProteinkinaseCinpain: involvementofmultipleisoforms.Pharmacol.Res.55,578–589.
Vellani,V.,Mapplebeck,S.,Moriondo,A.,Davis,J.B.,McNaughton,P.A.,2001.Protein kinaseCactivationpotentiatesgatingofthevanilloidreceptorVR1bycapsaicin, protons,heatandanandamide.J.Physiol.Lond.543,813–825.
Verma,P.R.,Joharapurkar,A.A.,Chatpalliwar,V.A.,Asnani,A.J.,2005.Antinociceptive activityofalcoholicextractofHemidesmusindicusR.Br.inmice.J. Ethnophar-macol.102,298–301.
Vogel,H.G.,Vogel,W.H.,1997.Pharmacologicalassays.In:DrugDiscoveryand Eval-uation.J.A.MajorsCompany,Lewisville,TX,USA,pp.360–418.
Vongtau,H.O.,Abbah,J.,Ngazal,I.E.,Kunle,O.F.,Chinbo,B.A.,Otsapa,P.B.,Gamaniel, K.S.,2004.Antinociceptiveandanti-inflammatoryactivityofthemethanolic extractofParinaripolyandrastembarkinratsandmice.J.Ethnopharmacol.90, 115–121.
Wirth,J.H.,CraigHudgins,J.,Paice,J.A.,2005.Useofherbaltherapiestorelievepain: areviewofefficacyandadverseeffects.PainManage.Nurs.6,145–167. Zakaria,Z.A.,Balan,T.,Suppaiah,V.,Ahmad,S.,Jamaludin,F.,2014a.Mechanism(s)
ofactioninvolvedinthegastroprotectiveactivityofMuntingiacalabura.J. Ethnopharmacol.3,1184–1193.
Zakaria,Z.A.,Fatimah,C.A.,MatJais,A.M.,Zaiton,H.,Henie,E.F.P.,Sulaiman,M.R., Somchit,M.N.,Thenamuha,M.,Kasthuri,D.,2006a.Theinvitroantibacterial activityofMuntingiacalaburaextracts.Int.J.Pharmacol.2,439–442. Zakaria,Z.A.,Kumar,G.H.,MohdZaid,S.N.H.,AbdulGhani,M.,Hassan,M.H.,Mohd
NorHazalin,N.A.,Khamis,M.M.,Devi,R.G.,2007a.Analgesicandantipyretic actionsofMuntingiacalaburaleaveschloroformextractinanimalmodels. Ori-ent.Pharm.Exp.Med.7,34–40.
Zakaria, Z.A.,Mohammad, A.M.,Mohd Jamil,N.S.,Rofiee,M.S.,Hussain, M.K., Sulaiman,M.R.,Teh,L.K.,Salleh,M.Z.,2011.Invitroantiproliferativeand antiox-idantactivitiesoftheextractsofMuntingiacalaburaleaves.Am.J.Chin.Med.39, 183–200.
Zakaria,Z.A.,MohdNorHazalin,N.A.,MohdZaid,S.N.H.,AbdulGhani,M.,Hassan, M.H.,Gopalan,H.K.,Sulaiman,M.R.,2007b.Antinociceptive,anti-inflammatory andantipyreticeffectsofMuntingiacalaburaaqueousextractinanimalmodels. J.Nat.Med.61,443–448.
Zakaria,Z.A.,MohdSani,M.H.,Cheema,M.S.,Arifah,A.K.,Teh,L.K.,Salleh,M.Z., 2014b.AntinociceptiveactivityofmethanolicextractofMuntingiacalabura leaves:furtherelucidation ofthepossible mechanisms.BMCComplement. Altern.Med.63,http://dx.doi.org/10.1186/1472-6882-14-63.
Zakaria,Z.A.,Sufian,A.S.,Ramasamy,K.,Ahmat,N.,Sulaiman,M.R.,Arifah,A.K., Zuraini,A.,Somchit,M.N.,2010.InvitroantimicrobialactivityofMuntigia cal-aburaextractsandfractions.Afr.J.Microbiol.Res.4,304–308.
Zakaria,Z.A.,Sulaiman,M.R.,MatJais,A.M.,Somchit,M.N.,Kogilla,V.J.,Ganesh,R., Fatimah,C.A.,2006b.TheantinociceptiveactivityofMuntingiacalabura aque-ousextractandtheinvolvementofl-arginine/nitricoxide/cyclic guanosine monophosphatepathwayinitsobservedactivityinmice.Fundam.Clin. Pharma-col.20,365–372.