ALICE MARIA DE SOUZA KANESHIMA
Microsporogênese em genótipos de
Brachiaria
(Poaceae) tetraploidizados artificialmente
Tese apresentada ao Programa de Pós-Graduação em Ciências Biológicas (área de concentração – Biologia Celular e Molecular) da Universidade Estadual de Maringá, para obtenção do grau de Doutor em Ciências Biológicas.
ALICE MARIA DE SOUZA KANESHIMA
Microsporogênese em genótipos de
Brachiaria
(Poaceae) tetraploidizados artificialmente
Orientadora:
Dra. Maria Suely Pagliarini
Co-Orientadora:
Dra. Cacilda Borges do Valle
Dados Internacionais de Catalogação-na-Publicação (CIP) (Biblioteca Central - UEM, Maringá – PR., Brasil)
Kaneshima, Alice Maria de Souza
K16m Microsporogênese em genótipos de Brachiaria (Poaceae) tetraploidizados artificialmente / Alice Maria de Souza Kaneshima. -- Maringá : [s.n.], 2008. 53 f. : figs., tabs.
Orientadora : Profª. Drª. Maria Suely Pagliarini. Co-Orientadora : Profª. Drª. Cacilda Borges do
Valle.
Dissertação (mestrado) - Universidade Estadual de Maringá, Programa de Pós-Graduação em Ciências Biológicas, 2008.
1. Citogenética vegetal. 2. Brachiaria - Estudo citogenético. 3. Diploidia. 4. Tetraploidia. 5. Meiose.I. Universidade Estadual de Maringá. Programa de Pós-Graduação em Ciências Biológicas. II. Título.
BIOGRAFIA
Alice Maria de Souza-Kaneshima, filha de Benedito Pereira de Souza e Antonia de Souza-Zane (in memorian), nasceu em Londrina-PR em 14 de novembro de 1965.
Aos meus amores, Luís Filipe e Edilson,
por toda paciência, compreensão e apoio.
Minhas desculpas pelo tempo não
compartilhado, em especial ao Luís Filipe,
onde espero que possamos recuperá-lo
Seguindo o exemplo da Virgem Maria:
“Minha alma glorifica ao Senhor, meu espírito exulta de alegria em Deus, meu Salvador, porque olhou para sua pobre serva. Por isto, desde agora, me proclamarão bem-aventurada todas as gerações, porque realizou em mim maravilhas aquele que é poderoso e cujo nome é Santo. Sua misericórdia se estende de geração em geração, sobre os que o temem. Manifestou o poder do seu braço: desconcertou os corações dos soberbos. Derrubou do trono os poderosos e exaltou os humildes. Saciou de bens os indigentes e despediu de mãos vazias os ricos. Acolheu a Israel, seu servo, lembrado da sua misericórdia, conforme prometera a nossos pais, em favor de Abraão e sua posteridade, para sempre. Amém”.
AGRADECIMENTOS
À minha orientadora Dra Maria Suely Pagliarini, pelo incentivo, estímulo, apoio, e tolerância nos momentos de dificuldades e pelos grandes ensinamentos transmitidos. Minha gratidão e o meu respeito pelo seu exemplo de profissionalismo e amizade.
À minha co-orientadora Dra. Cacilda Borges do Valle, pesquisadora da Embrapa Gado de Corte, pelos ensinamentos e sugestões ao longo do desenvolvimento deste trabalho e também por colocar à disposição do Laboratório de Citogenética da UEM os genótipos analisados.
A todos os colegas do Laboratório de Citogenética Vegetal pelo carinho e amizade, em especial à Mariana Ferrari Felismino e Gléia Cristina Laverde Ricci pelo empenho e disposição em várias etapas no desenvolvimento deste trabalho, principalmente na coleta de alguns dos genótipos analisados e também à Veridiana Aparecida Fuzinatto pelo auxílio na montagem das fotos.
À técnica do Laboratório de Citogenética Vegetal da UEM, Neide da Silva, pela dedicação e exemplo de profissionalismo na preparação de materiais utilizados em meus experimentos.
A todos os professores e amigos do Departamento de Análises Clínicas e do Programa de Pós-Graduação em Ciências Biológicas – área de concentração Biologia Celular e Molecular que me ofereceram apoio e incentivo.
APRESENTAÇÃO
Esta tese, intitulada Microsporogênese em genótipos de Brachiaria (Poaceae) tetraploidizados artificialmente, é composta por três artigos.
1. Cytogenetic advances in Brachiaria (Poaceae) breeding: meiotic behavior in the first interspecific hybrid between B. brizantha and B. decumbens, a ser enviado para publicação no periódico Plant Breeding (Alemanha), Fator de Impacto 1.092 (JCR 2007).
2. Abnormal meiosis in tetraploid genotypes of Brachiaria brizantha (Poaceae) induced by colchicine: its implications for the breeding, enviado para publicação no periódico Journal of Applied Genetics (Alemanha), Fator de Impacto 0.967 (JCR 2007).
Microsporogenesis in
Brachiaria
(Poaceae) genotypes
artificially tetraploidized
Alice Maria de Souza Kaneshima Maria Suely Pagliarini (Adviser) Cacilda Borges do Valle (Co-adviser)
Abstract
Many genera of the Poaceae family are economically important forage grasses. An example is the genus Brachiaria, native to the African savannas, that encompasses more
than 100 species. However, the commercially exploited brachiariagrasses around the world belong to four African species: B. brizantha (palisadegrass), B. decumbens
(signalgrass), B. humidicola (koroniviagrass), and B. ruziziensis (ruzigrass). Their
economic importance is greatest in tropical America where extensive adoption over the past several decades has had a revolutionary impact on cattle productivity placing Brazil as the largest beef exporter in the world. Although eight cultivars of Brachiaria are
commercialized in Brazil by a vital seed industry, seven of them were originated as direct selections from germplasm collected in Africa. The Brazilian Brachiaria breeding
program underway at Embrapa Beef Cattle aims at producing new cultivars by intra- and interspecific hybridization. However, Brachiaria hybridization is not easy because
of apomictic reproduction and differences in the ploidy level among species. In the genus Brachiaria diploidy is rare and correlated with sexual reproduction whereas
polyploidy and apomixis predominate. To overcome the barrier between sexual diploid and apomictic tetraploid accessions, it was proposed to create a cross-compatible, sexual tetraploid plant by doubling the chromosome number. The barrier was overcome in Belgium, in 1981, when diploid accessions of B. ruziziensis were tetraploidized with
Agricultura Tropical, Colombia) and in Brazil. In attempts to broaden the genetic basis of Brachiaria breeding in Brazil, the most important Brachiaria species in the Brazilian
pastures (B. brizantha) were also submitted to artificial tetraploidization to allow
intraspecific hybridization among accessions with high resistance to the spittlebugs. The unique diploid accession of B. brizantha (B105) was recently submitted to chromosome
doubling by colchicine treatment and six sexual tetraploid plants were obtained. With the same objective, another approach was used to obtain sexual tetraploid compatible plants in Brachiaria. Two sexual diploid accessions of Brachiaria (B. brizantha, B105 x
B. decumbens, D004) were crossed and the diploid F1 generation (2n = 2x = 18) was
tetraploidized to obtain chromosome homology. Two tetraploid (2n = 4x = 36) plants were obtained. Considering that polyploidy is largely correlated to abnormal meiosis in
Brachiaria and the fact that apomictic plants are pseudogamic, which means that fertile
gametes are necessary for endosperm development, the current research aimed to evaluate the meiotic behavior in progenies of induced tetraploidized plants as a subsidy to the selection of the best plants to act as female genitors in intraspecific and interespecific crosses. All the diploid accessions were collected in the wild African savannas by CIAT, in 1984-1985. Chromosome doubling was accomplished by colchicine treatment of basal segments of in vitro grown plants. Inflorescences for
meiotic studies were collected from individual plants growing at Embrapa Beef Cattle Research Center, located in Campo Grande, Mato Grosso do Sul, Brazil. They were fixed in a mixture of ethanol 95%, chloroform and propionic acid (6:3:2) for 24 hours, transferred to 70% alcohol and stored under refrigeration until use. Microsporocytes were prepared by squashing and staining with 0.5% propionic carmine. Meiotic abnormalities were observed in each phase of microsporogenesis. Photomicrographs were made with a Wild Leitz microscope using Kodak Imagelink – HQ, ISO 25 black and white film. Twenty two accessions of B. ruziziensis available at Embrapa Beef
convergence to the poles (divergent spindle), was recorded. For B. brizantha, six
progenies from the three artificially induced tetraploid sexual genotypes were analyzed. The percentage of abnormal cells among plants ranged from 39.8% to 63.2%. In a plant derived from one progeny, only the common meiotic abnormalities typical of polyploids were observed, while in plants derived from the two other progenies, a chaotic spindle
was formed and the chromosomes remained spread in the cytoplasm in the first division without forming a metaphase plate. The chaotic spindle organization blocked
homologous segregation at anaphase I. Several micronuclei of different sizes were formed and, after the occurrence of an irregular first cytokinesis, the meiocytes progressed normally to the second division, generating polyads with unbalanced microspores. In the diploid accessions of B. brizantha and B. decumbens, meiosis was
quite normal, while in the diploid hybrid plants of the F1 generation, besides some irregular chromosome segregation, parental genome separation was found. This phenomenon was also observed in the amphidiploid hybrids. In the tetraploidized plants, a high number of meiocytes with chaotic spindles was also recorded in both meiosis. The abnormalities compromised the meiotic products that were characterized by polyads. Chromosome association in quadrivalents at diakinesis in the amphidiploid progenies shows that introgression of desirable genes can be obtained. The main objective of chromosome doubling in these sexual diploid plants was to obtain sexual tetraploids to act as female genitors in intra- and intraspecific crosses with natural apomictic tetraploid pollen donors. To accomplish this objective, the meiotic process of tetraploidized plants must be regular to ensure pollen fertility, embryo fertilization and proper seed set. Intraspecific hybridization is expected to yield hybrids with less seed set problems than what has been observed so far in interspecific hybrids. The use of sexual tetraploidized plants in crosses with natural apomictic accessions of other compatible Brachiaria species needs to take into account the results obtained in the
meiotic analysis to avoid carry-over meiotic abnormalities into the progenies. Thus, the analysis of meiotic behavior is the best tool to select progenitors in the Brachiaria
Microsporogênese em genótipos de
Brachiaria
(Poaceae)
tetraploidizados artificialmente
Alice Maria de Souza Kaneshima Maria Suely Pagliarini (Orientadora) Cacilda Borges do Valle (Co-orientadora)
Resumo
Muitos gêneros da família Poaceae são gramíneas forrageiras importantes. Um exemplo é o gênero Brachiaria, nativo das savanas africanas que engloba mais de 100 espécies.
Todavia, as gramíneas forrageiras deste gênero exploradas no mundo pertencem a quatro espécies: B. brizantha, B. decumbens, B. humidicola e B. ruziziensis. Sua
importância econômica é maior na América tropical onde a extensiva adoção nas últimas décadas teve um impacto revolucionário na pecuária, tornando o Brasil o maior exportador de carne do mundo. Embora oito cultivares de Brachiaria sejam
comercializadas no Brasil pela indústria de sementes, sete delas foram originadas por seleção direta do germoplasma coletado na África. O programa de melhoramento de
Brachiaria em desenvolvimento na Embrapa Gado de Corte objetiva produzir novas
cultivares por hibridização intra- ou interespecífica. Todavia, a hibridização em
Brachiaria não é fácil de ser realizada por causa da reprodução apomítica e diferenças
no nível de ploidia entre acessos e espécies. No gênero Brachiaria, diploidia é rara e
está associada com reprodução sexuada, enquanto a poliploidia e a apomixia predominam. Para vencer a barreira entre diplóides sexuais e acessos tetraplóides apomíticos, propôs-se a criação de cruzamentos compatíveis utilizando-se plantas sexuais tetraplóides obtidas por duplicação do número de cromossomos. Esta barreira foi quebrada com colchicina. Seis acessos sexuais tetraplóides foram obtidos e continuam, até hoje, servindo como base para o programa de hibridização interespecífica no Centro Internacional de Agricultura Tropical (CIAT, Colômbia) e no Brasil. Na tentativa de ampliar a base genética no programa de melhoramento de
Brachiaria no Brasil, a espécie mais importante de Brachiaria para as pastagens
brasileiras (B. brizantha) foi submetida à tetraploidização para permitir hibridização
único acesso diplóide de B. brizantha (B105) foi recentemente submetido à duplicação
de cromossomos através de tratamento com colchicina. Seis plantas tetraplóides sexuais foram obtidas. Com o mesmo objetivo, outra abordagem foi usada para obter plantas tetraplóides sexuais em Brachiaria. Dois acessos sexuais de Brachiaria (B. brizantha,
B105 x B. decumbens, D004) foram cruzados e a geração F1 diplóide (2n = 2x = 18) foi tetraploidizada, também com o uso de colchicina, para se obter homologia cromossômica. Duas plantas tetraplóides (2n = 4x = 36) foram obtidas. Considerando que a poliploidia é altamente correlacionada com meiose anormal em Brachiaria e o
fato de que plantas apomíticas são pseudogâmicas, o que significa que gametas viáveis são necessários para o desenvolvimento do endosperma, a presente pesquisa objetivou avaliar o comportamento meiótico em progênies de plantas tetraploidizadas como um subsídio para a seleção das melhores plantas para atuar como genitores femininos em cruzamentos intra ou interespecíficos. Todos os acessos diplóides foram coletados nas savanas da África pelo CIAT, entre 1984-1985. A duplicação de cromossomos foi feita pelo uso de colchicina aplicada em segmentos basais de plantas cultivadas in vitro.
Inflorescências para estudos meióticos foram coletadas em plantas individuais, crescidas em campo, na Embrapa Gado de Corte, localizada em Campo Grande, Mato Grosso do Sul, Brasil. As inflorescências foram fixadas em uma mistura de etanol 95%, clorofórmio e ácido propiônico (6:3:2) por 24 horas, transferidas para álcool 70% e estocadas sob refrigeração até o momento de serem utilizadas. Os microsporócitos foram preparados pela técnica de esmagamento e corados com carmim propiônico 0,5%. As anormalidades foram consideradas em cada fase da microsporogênese. As fotomicrografias foram feitas em microscópio Wild Leitz, usando-se filme branco e preto, Kodak Imagelink – HQ, ISO 25. Vinte e dois acessos de B. ruziziensis
disponíveis na Embrapa Gado de Corte foram citologicamente analisados: 16 eram diplóides (2n = 2x = 18) e seis eram tetraplóides artificiais (2n = 4x = 36). O comportamento meiótico dos 16 acessos diplóides foi variável. A média de anormalidades meióticas por acesso variou de zero a 24,5%. Nos acessos tetraplóides artificiais, a média de anormalidades variou entre 5,2 a 54,7%. As anormalidades mais comuns observada entre os acessos diplóides e tetraplóides foram aquelas relacionadas à segregação irregular de cromossomos em ambas as divisões meióticas. Em um acesso tetraplóide, outra irregularidade envolvendo a orientação dos cromossomos na placa metafásica e convergência de cromossomos para os pólos (fuso divergente) foi
artificiais, foram analisadas. A percentagem de células anormais entre as plantas tetraploidizadas variou de 39,8 a 63,2%. Em uma planta derivada de uma progênie, somente as anormalidades típicas de poliplóides foram observadas, enquanto em plantas derivadas das outras duas progênies, um fuso caótico foi formado e os cromossomos permaneceram espalhados na primeira divisão sem que a placa metafásica fosse formada. A formação do fuso caótico bloqueou a segregação dos cromossomos homólogos na anáfase I. Em conseqüência, muitos micronúcleos de diferentes tamanhos foram formados e, após a ocorrência da primeira citocinese, os meiócitos progrediram normalmente na segunda divisão, gerando políades com micrósporos geneticamente desbalanceados. Nos acessos diplóides de B. brizantha e B. decumbens, a meiose foi
Cytogenetic advances in
Brachiaria
(Poaceae)
breeding: meiotic behavior in the first interspecific
hybrid between
B. brizantha
and
B. decumbens
Alice Maria de Souza-Kaneshima
1, Carine Simioni
2, Mariana Ferrari
Felismino
1, Andréa Beatriz Mendes-Bonato
1, Claudicéia
Risso-Pascotto
1, Maria Suely Pagliarini
1*, and Cacilda Borges do Valle
31. Department of Cell Biology and Genetics, State University of Maringá,
87020-900 Maringá PR Brazil.
2. Federal University of Rio Grande do Sul, Porto Alegre RS Brazil.
3. Embrapa Beef Cattle, P.O. Box 154, 79002-970 Campo Grande MS
Brazil.
* Author for correspondence: Maria Suely Pagliarini (E-mail:
[email protected]
).
ADDRESS FOR CORRESPONDENCE
Dra. Maria Suely Pagliarini
Department of Cell Biology and Genetics
State University of Maringá
87020-900 Maringá, Paraná - Brazil.
Cytogenetic advances in
Brachiaria
(Poaceae)
breeding: meiotic behavior in the first interspecific
hybrid between
B. brizantha
and
B. decumbens
Abstract
. Brachiaria brizantha and B. decumbens constitute the two most widely usedtropical forage species for cultivated pastures and support both beef and dairy cattle production in the tropics. Two apomictics cultivars – B. brizantha cv. Marandu and B.
decumbens cv. Basilisk – cover more than 100 million hectares of cultivated pastures
throughout Latin America and Southeast Asia. This paper discusses the cytogenetic data of the first successful attempt in obtaining interespecific hybrids between these two species. Artificial hybridization between two diploid (2n = 2x = 18) sexual accessions (B. brizantha, B105 x B. decumbens, D004) was performed in the greenhouse. Only
three hybrids were recovered. One of them was treated with colchicine in tissue culture and two amphidiploid plants were obtained. Meiosis in the diploid genitors was quite normal, while in the diploid hybrid parental genome separation was observed together with some irregular chromosome segregation. Genome separation was also observed in several cells in the two 4n amphidiploid hybrids. In these, a high number of meiocytes with chromosomes chaotically spread in the cytoplasm was recorded during meiosis. The abnormalities observed compromised the meiotic products characterized by polyads. Chromosome association as quadrivalents in diakinesis reinforces the proposed taxonomic proximity between these two species. Moreover, it indicates that introgression of desirable genes is feasible between these two most important agronomic species of the genus.
Key words: Brachiaria brizantha, B. decumbens, interspecific hybrid, chromosome
Introduction
Wide hybridization has great potential for transferring valuable traits from allied species to cultivated crops. However, incompatibility barriers between species may pose a major constraint to the effective use of this strategy in breeding programs. Plant breeders have frequently used interspecific hybridization for the transfer of genes from a wild plant species to a related crop variety. If a hybrid plant is recovered, differences in the number or compatibility of parental chromosomes may cause sterility. Cytogenetic manipulations have been useful in obtaining stable gene transfers. Sterility may result from incomplete or unstable chromosome pairing during cell division. Chromosome doubling can facilitate chromosome pairing and restore fertility in amphidiploids (Goodman et al., 1987). However, for a desired gene from the wild species to be incorporated into a chromosome of the crop variety, recombination must take place which implies close relationship between the two species.
Hybridization in the genus Brachiaria is a complex undertaking due to apomixis
and polyploidy. The two major species: B. brizantha and B. decumbens are
predominantly tetraploid and apomictic. Natural sexual compatible genotypes were not identified. Brachiaria ruziziensis, a sexual diploid species belonging to the same agamic
The majority of Brazilian pastures are constituted by only two apomictic cultivars, B. brizantha cv. Marandu and B. decumbens cv. Basilisk, introduced to Brazil
only in the second half of the last century. With their widespread use as pastures, exceptional plant adaptation and productivity, beef cattle production increased greatly and the country became the largest exporter and the second producer of beef in the world. These cultivars cover more than 70 million of hectares in the central region of Brazil and over 100 million hectares considering Latin America and Southeast Asia. New Brachiaria cultivars to diversify pastures in Brazil are urgently required and
comprise the major objective of the ongoing Brachiaria breeding program in Embrapa.
Candidate cultivars are either selected from the natural germplasm or from progenies derived from interspecific hybridization. Brachiaria brizantha and B. decumbens are
considered pivotal species in the genus since the first contains accessions resistant to the major insect, spittlebugs, and the second, although lacking resistance to the insect, is well adapted to infertile, acid soils generally found in the Brazilian savannas (Keller-Grein et al., 1996).
The gene pools in the genus Brachiaria are not yet well defined. The genus
comprises of more than 100 species of African origin. A throrough taxonomic study of a large germplasm collection classified Brachiaria brizantha and B. decumbens in the
same taxonomic group together with four other species (Renvoize et al., 1996). These two species are compatible and closely related and, at times, difficult to distinguish.
Brachiaria breeding is difficult to accomplish because of differences in chromosome
brizantha (B105) out of 220 accessions, and 23 out of 50 accessions of B. decumbens
were identified in the germplasm collection of this species in the Embrapa Beef Cattle Research Center (Valle and Savidan, 1996; Penteado et al., 2000; Mendes-Bonato et al., 2002a). All of those are sexual as determined by embryo-sac analysis using methylsalicilate clearing and interference contrast microscopy (Young et al., 1979).
The Brachiaria hybridization program at Embrapa Beef Cattle produced
interspecific hybrids from sexual artificially tetratploid accessions of B. ruziziensis
obtained in Belgium in the 1980s (Gobbe et al., 1981; Swenne et al., 1981) with apomictic natural tetraploid accessions of B. brizantha or B. decumbens (Valle et al.,
2008) as pollen donors. Several hybrids are under agronomic evaluation to identify future cultivars.
An attempt to produce diploid hybrids from B. brizantha and B. decumbens was
performed by crossing the single diploid accession of B. brizantha available (B105)
with one diploid accession of B. decumbens. In order to create perfect homology for
chromosome pairing to guarantee normal meiosis, the diploid plants of F1 generation were artificially tetraploidized by colchicine. This study reports the meiotic behavior in the diploid parents and the F1 plants, and in the tetraploidized plants as subsidy to understand the low seed production detected in these amphidiploid plants.
Materials and Methods
The Brachiaria germplasm collection existing at Embrapa Beef Cattle Research
(Cenargen/Embrapa, Brasilia, DF, Brazil). After quarantine, the Brachiaria collection
was transferred to Embrapa Beef Cattle (Campo Grande, MS, Brazil) to serve as the basis for the breeding program.
Hybridization between parental genotypes (B. brizantha, B105 and B. decumbens, D004) was performed in the greenhouse, using pollen from the D004 in
prepared flowers of B105. Seeds obtained were individually germinated in Campo Grande (MS) and the three surviving plantlets were later transferred to the field. Basal segments of these hybrids were cultured in vitro and treated with colchicine at 0.01%
applied directly to the culture medium for 48 hours. Afterwards, explants were placed in culture medium without colchicine, and surviving buds were transferred to MS (Murashige and Skoog, 1962) medium for rooting. Only two plantlets (8 and CH4-100) were successfully polyploidized with data confirmed by chromosome countings in root tips (C. Simioni, unpublished data).
Results
Table 1 presents the data obtained from the analysis of microsporogenesis in the diploid and amphidiploid plants. The meiotic behavior was quite normal in the female (B105) and male (D004) genitors. In the first division, some precocious chromosome migration to the poles in metaphase, laggards in anaphases, leading to micronuclei formation in telophase were recorded, but in low frequencies. In the second division, only a few meiocytes displayed some abnormalities, and the meiotic products were characterized by normal tetrads. The average percentage of meiotic abnormalities was 2.6% in B105 and 2.1% in D004.
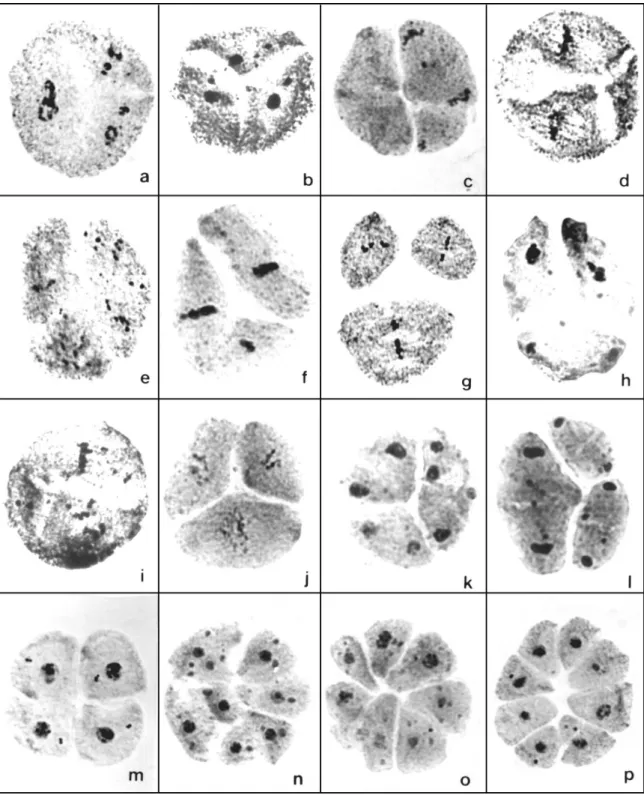
In the diploid hybrid, unfortunately, chromosome association at diakinesis could not be evaluated, because this phase was not found among anthers extensively analyzed. Meiotic abnormalities related to irregular chromosome segregation were recorded in similar frequency to those observed in their genitors. However, in the diploid hybrid, genome separation was detected in a high number of cells since pachytene. In this phase, both parental genomes were distantly positioned in the cytoplasm (Fig. 1a). The inability to share the same territory inside the cell was also detected in metaphase I, when two metaphase plates were organized (Fig. 1b). In the following phases of meiosis, genome separation was not found among meiocytes, but polyads were recorded as meiotic products. These abnormalities increased the frequency of abnormal cells to 17.8% in the diploid F1 generation.
Table 1. Number of normal and abnormal cells recorded in each genotype
Phases Abnormalities Female genitor B105
(2n = 18)
Male genitor D004 (2n = 18)
Hybrid (B105 x D004)
(2n = 18)
Hybrid CH4-8 (2n = 36)
Hybrid CH4-100 (2n = 36)
Pachytene Normal cells 137 -* 75 100 60
Genome separation 0 0 35 62 25
Diakinesis Normal cells 20 - - - 23
Quadrivalents - - - - 13
Genome separation 0 0 - - 10
Metaphase I Normal cells 243 189 67 200 67
Genome separation 0 0 12 20 25
Precocious migration 10 0 0 16 10
Chaotic spindle 0 0 0 204 116
Multiple spindle 0 0 0 15 7
Anaphase I Normal cells 202 20 94 110 92
Genome separation 0 0 0 13 2
Laggards 19 8 7 16 18
Tripolar spindle 0 0 0 34 4
Irregular segregation 0 0 8 0 5
Telophase I Normal cells 179 123 62 89 69
Micronuclei 10 5 0 60 40
Four nuclei 0 0 0 8 17
Three nuclei 0 0 0 10 35
Prophase II Normal cells 65 74 4 2 10
Micronuclei 0 4 - - -
Microcytes 0 - - 1 -
Metaphase II Normal cells 32 87 32 35 31
Multiple spindle 0 0 0 46 11
Microcytes 0 0 0 1 1
Chaotic spindle 0 0 0 43 52
Anaphase II Normal cells 123 34 28 58 18
Multiple spindle 0 0 - 12 3
Laggards 0 1 4 0 0
Irregular segregation 0 0 1 0 0
Telophase II Normal cells 141 181 19 19 26
Multinucleated cells 0 0 0 14 7
Micronuclei 0 2 18 4 0
Tetrads Normal microspores 346 215 56 40 97
Micronuclei 0 0 0 5 1
Microcytes 0 0 1 28 53
Triads 0 0 0 4 5
Polyads 0 0 9 17 21
No. of cells evaluated 1527 943 532 1286 974 No. (%) of abnormal cells 39 (2.6) 20 (2.1) 95 (17.8) 633 (49.2) 468 (50.80)
giving rise to tetranucleated telophase I (Fig. 1d) were recorded. In the second division, four cells, instead the two normal ones, were observed (Fig. 1e). Polyads were also recorded among the meiotic products (Fig. 1f).
In the amphidiploid plants, however, another abnormality related to spindle organization was detected in high frequency (see Table 1). In the affected cells, meiosis was normal until diakinesis. From this phase on, chromosomes were chaotically dispersed in the cytoplasm because of absence of a spindle organizing center in the poles (Fig. 2a). In these cells, anaphase I did not occur and chromosomes, alone or in groups, generated several telophase nuclei of different sizes (Fig. 2b). In these cells, the first cytokinesis was abnormal, dividing the meiocyte in more than two cells (Fig. 2c). The plane of cytokinesis was apparently determined by the position of the telophase nuclei. In the second division, meiocytes with multiple spindle were abundantly recorded (Fig. 2d, e), generating multinucleated cells in telophase II (Fig. 2f). In both amphidiploid hybrids, meiotic products were highly abnormal, with several micronuclei and microcytes in the tetrads (Fig. 2g, h) generating pollen grains of different sizes (Fig. 2i). The percentagem of abnormal cells in the tetraploid progenies was similar, 49.2% in CH4-8 and 50.8% CH4-100.
Discussion
precociously to the poles or those that remained laggard, in general, were not included in the telophase nuclei and formed micronuclei. In the second division, the chromosomes forming micronuclei were included in the second metaphase plate, decreasing the frequency of meiotic abnormalities.
Unfortunately, genome affinity between B. brizantha and B. decumbens could
not be evaluated in the diploid F1 generation because diakinesis was not present in the analyzed anthers. But, considering the data obtained from chromosome association in the amphidiploid progenies, where one to three quadrivalents were recorded on the 13 cells analyzed, we can assert that some affinity exists between them. Chromosome pairing indicates that genetic recombination can occur between both species and thereby gene introgression of desirable traits, such as resistance to spitllebugs and tolerance to infertile soils, can be expected. Genome affinity between these species was never yet evaluated from the cytological view in interspecific hybrids and more studies are necessary to better understand the relationship degree among them. Jauhar and Joppa (1996) have pointed that the analysis of chromosome behavior, mainly during pachytene and metaphase I, can provide valuable evidence of genomic relationship.
Spindle irregularities, like those observed in the amphidiploid progenies obtained from colchicine treatment were also recorded in accessions of B. ruziziensis
(Pagliarini et al., 2008) and in B. brizantha (Mendes-Bonato et al., 2008), also
tetraploidized by this drug. Anaphase movement was not found among meiocytes these meiocytes of the amphidiploid progenies, thus homologous chromosome segregation did not occur in the first division. However, cytokinesis occurred dividing the meiocyte into several cells of different sizes. Similar pattern of cytokinesis was observed during microsporogenesis in an artificially induced tetraploid accession of B. ruziziensis
Bonato et al., 2002b) where spindle disorganization also occurred in the second division, fractionating the genome into polyads with several microcytes.
Genome separation and genome elimination are common among interspecific hybrids. Davies (1974) and Shwarzacher et al. (1992) reported several examples among plants where the phenomenon occurred. According to Luckens et al. (2006), the formation of natural allopolyploids requires the adaptation of two nuclear genomes within a single cytoplasm, which may involve programmed genetic changes during the first generation following genome fusion. The incompatibility of cytoplasm could result from DNA replication and other cell processes (Subrahmanyam and Kasha, 1973). In the present diploid interspecific Brachiaria hybrid and its amphidiploid progenies,
genome separation was visualized since the early meiotic phases until the end of meiosis. The phenomenon was manifested as an inability of the two genomes to share the same territory within the meiocyte, i.e., each genome has its own domain. Thus, the parental genomes tended to lie spatially separated in the cytoplasm. Karyotypic characteristics were not known for the accessions of the B. brizantha and B. decumbens
used as the genitors in the present crossing. However, determination of the DNA content by flow cytometry shows a significative difference between the parental accessions: 1.41 pg of DNA in B105 and 1.0 pg in D004 (Penteado et al., 2000). Genome separation in somatic and premeiotic metaphases was reported in interspecific
Hordeum hybrids where genomes differed substantially in the DNA content
Genome affinity in the genus Brachiaria is not well studied from the cytogenetic
view. Karyological studies in the genus were performed for a few species and accessions (Bernini and Marin-Morales, 2001), and genomes were not yet described. In the last decade, however, evaluation of the meiotic behavior in some hybrids between B.
ruziziensis x B. brizantha or B. decumbens (Risso-Pascotto et al., 2004; Mendes-Bonato
et al., 2006b; Adamowski et al., 2008; Fuzinatto et al., 2008) has revealed that genomes of parental species not always show perfect affinity. Lacking of genome affinity during meiosis was revealed by asynchrony in the meiosis time (Risso-Pascotto et al., 2004) or separation of the two parental genomes (Mendes-Bonato et al., 2006b), like that found in the present diploid and amphidiploid hybrids. Although interspecific tetraploid hybrids among the above species always present a high frequency of meiotic abnormalities, the lack of genome affinity between them seems to be genotype-specific. This is the first attempt to cross B. brizantha with B. decumbens. The result
obtained from the crossing between B105 (2n) with D004 (2n) accession, was not good for breeding purposes. However, since around of 50% of meiotic products was normal in the two amphidiploid hybrids and recombination among the parental genomes could be expected, the progenies of them might be evaluated in order to select promising genotypes. Recently, Pinheiro et al. (2000) duplicated the chromosome number of this diploid accession (B105) of B. brizantha. The 4n sexual progenies are under cytological
Acknowledgements
Authors are grateful to UNIPASTO for financial support.
References
Adamowski, E.V., M. S. Pagliarini, and C. B. Valle, 2008: Meiotic behavior in three interspecific 3-way hybrids between Brachiaria ruziziensis and B. brizantha (Poaceae:
Paniceae). J. Genet. 87, 33-38.
Bernini, C., and M. A. Marin-Morales, 2001: Karyotype analysis in Brachiaria
(Poaceae) species. Cytobios 104,157-171.
Davies D. R., 1974: Chromosome elimination in interspecific hybrids. Heredity 32, 267-270.
Fuzinatto, V. A., M. S. Pagliarini, and C. B. Valle, 2008: Microsporogenesis in sexual
Brachiaria hybrids (Poaceae). Genet. Mol. Res. 6, 1107-1117.
Gleba, Y. Y., A. Parokonny, V. Kotov, I. Negrutiu, and V. Momot, 1987: Spatial separation of parental genomes in hybrids of somatic plant cells. Proc. Nat. Ac. Sci. USA. 84, 3709-3713.
Gobbe, J., A. Swenne, and B. P. Louant, 1981: Diploïdes naturels et autotétraploïdes induits chez Brachiaria ruziziensis Germain et Evrard: critères d’identification. Agron.
Trop. 36, 339-346.
Jauhar, P. P., and L. R. Joppa, 1996: Chromosome pairing as a tool in genome analysis: merits and limitations. In: Jauhar, P.P (ed.), Methods of genome analysis in plants. 9-37. CRC Press, Boca Raton.
Keller-Grein, G., Maass, B. L., and J. Hanson, 1996: Natural variation in Brachiaria
and existing germoplasm collections. In: Miles, J. W., B. L. Maass, and C. B. Valle (eds.) Brachiaria: Biology, Agronomy, and Improvement, 17- 42. CIAT/Embrapa,
Colombia.
Linde-Laursen, I, and J. Jensen, 1991: Genome and chromosome disposition at somatic metaphase in a Hordeum x Psathyrostachys hybrid. Heredity 66, 203-210.
Luckens, L. N., J. C. Pires, G. Leon, R. Vogelzang, L. Oslack, and T. Osborn, 2006: Patterns of sequence loss and cytosine methylation within a population of newly resynthesized Brassica napus allopolyploids. Plant Physiol. 140, 336-348.
Mendes-Bonato, A. B., M. S. Pagliarini, F. Forli, C. B. Valle, and M. I. O. Penteado, 2002a: Chromosome number and microsporogenesis in Brachiaria brizantha
(Gramineae). Euphytica 125, 419-425.
Mendes-Bonato, A. B., R. G. Junqueira Filho, M. S. Pagliarini, C. B. Valle, and M. I. O. Penteado, 2002b: Unusual cytological patterns of microsporogenesis in Brachiaria
decumbens: abnormalities in spindle and defective cytokinesis causing precocious
cellularization. Cell Biol. Int. 26, 641-646.
Mendes-Bonato, A. B., M. S. Pagliarini, C. Risso-Pascotto, and C. B. Valle, 2006a: Chromosome number and meiotic behavior in Brachiaria jubata (Gramineae). J. Genet.
Mendes-Bonato, A. B., C. Risso-Pascotto, M. S. Pagliarini, and C. B. Valle, 2006b. Cytogenetic evidence for genome elimination during microsporogenesis in interspecific hybrid between Brachiaria ruziziensis and B. brizantha (Poaceae). Genet. Mol. Biol. 29,
711-714.
Mendes-Bonato, A. B., M. F. Felismino, V. Calisto, A. M. Souza-Kaneshima, M. S. Pagliarini, and C. B. Valle, 2008: Abnormal meiosis in tetraploid genotypes of
Brachiaria brizantha (Poaceae) induced by colchicine: its implications for the breeding.
J. Appl. Genet. (submitted)
Murashige, T., and F. Skoog, 1962: A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15, 473-497.
Pagliarini, M. S., C. Risso-Pascotto, A. M. Souza-Kaneshima, and C. B. Valle, 2008: Analysis of meiotic behavior in selecting potential genitors among diploid and artificially induced tetraploid accessions of Brachiaria ruziziensis (Poaceae). Euphytica
164: 181-187.
Penteado, M. I. O., A. C. M. Santos, I. F. Rodrigues, C. B. Valle, M. A. C. Seixas, and A. Esteves, 2000: Determinação de poliploidia e avaliação da quantidade de DNA total em diferentes espécies de Brachiaria. Boletim de Pesquisa No. 11, 19p. Campo
Grande-MS, Embrapa Gado de Corte.
Pinheiro, A. A., M. T. Pozzobon, C. B. Valle, M. I. O. Penteado, and V. T. C. Carneiro, 2000: Duplication of the chromosome number of diploid Brachiaria brizantha plants
using colchicine. Plant Cell Rep. 19, 274-278.
Valle (eds.) Brachiaria: Biology, Agronomy, and Improvement, 1-15. CIAT/Embrapa,
Colombia – Brasilia.
Risso-Pascotto, C., M. S. Pagliarini, C.B. Valle, and L, Jank L, 2004: Asynchronous meiotic rhythm as the cause of selective chromosome elimination in an interspecific
Brachiaria hybrid. Plant Cell Rep. 22, 945-950.
Risso-Pascotto, C., M. S. Pagliarini, and C. B. Valle, 2005: Multiple spindles and cellularization during microsporogenesis in an artificially induced tetraploid accession
of Brachiaria ruziziensis (Gramineae). Plant Cell Rep. 23, 522-527.
Risso-Pascotto, C., M. S. Pagliarini, and C. B. Valle, 2006: Microsporogenesis in
Brachiaria dictyoneura (Fig. & De Not.) Stapf (Poaceae: Paniceae). Genet. Mol. Res.
5, 837-845.
Schwarzacher, T., J. S. Heslop-Harrison, and K. Anamthawat-Jónsson, 1992: Parental genome separation in reconstructions of somatic and premeiotic metaphases of
Hordeum vulgare x H. bulbosum. J. Cell Sci. 101, 13-24.
Subrahmanyam, N. C, and K. J. Kasha, 1973: Selective chromosomal elimination during haploid formation in barley following interspecific hybridization. Chromosoma 42, 111-125.
Swenne, A., B. P. Louant, and M. Dujardin, 1981: Induction par la colchicine de formes autotétraploïdes chez Brachiaria ruziziensis Germain et Evrard (Graminée). Agron.
Utsunomiya, K. S., M. S. Pagliarini, and C. B. Valle, 2005: Microsporogenesis in tetraploid accessions of Brachiaria nigropedata (Ficalho & Hiern) Stapf (Gramineae).
Biocell 29, 295-301.
Valle, C. B, C. Simioni, R. M. S. Resende and Jank, 2008: Melhoramento genético de
Brachiaria. In: Melhoramento Genético de Forrageiras Tropicais. Resende R. M. S.
(ed.) 13-53, Embrapa Gado de Corte, Campo Grande, MS.
Valle, C. B., and Y. H. Savidan, 1996: Genetics, cytogenetics, and reproductive biology of Brachiaria. In: Miles, J. W., B. L. Maass, and C. B. Valle (eds.) Brachiaria: Biology,
Agronomy, and Improvement, 147-163. CIAT/Embrapa, Colombia.
Abnormal meiosis in tetraploid genotypes of
Brachiaria
brizantha
(Poaceae) induced by colchicine: its
implications for the breeding
Maria de Souza Kaneshima
1, Andrea Beatriz Mendes-Bonato
1,
Mariana Ferrari Felismino
1, Vergílio Calisto
1, Maria Suely Pagliarini
1and Cacilda Borges do Valle
2.
1.Department of Cell Biology and Genetics, State University of Maringá,
Maringá PR, Brazil
2.Embrapa Beef Cattle, Campo Grande MS, Brazil.
* Author for correspondence: Maria Suely Pagliarini (E-mail:
[email protected]
).
RUNNING TITLE
:
Abnormal meiosis in tetraploidized BrachiariabrizanthaADDRESS FOR CORRESPONDENCE
Dra. Maria Suely Pagliarini
Department of Cell Biology and Genetics
State University of Maringá
87020-900 Maringá – Paraná, Brazil
Abnormal meiosis in tetraploid genotypes of
Brachiaria
brizantha
(Poaceae) induced by colchicine: its
implications for the breeding
Abstract.
Six progenies from the three artificially induced tetraploid (2n = 4x = 36) sexual genotypes (C31, C41, and C48) of B. brizantha were analyzed in their meioticbehavior. These are key plants to allow intraspecific hybridization of this important forage species widely used for pastures in the tropics. The percentage of abnormal cells among plants ranged from 39.8% to 63.2%. In the single plant derived from C48, only the common meiotic abnormalities typical of polyploids were observed, while in plants derived from C31 and C41, a distinct behavior was found. In the majority of cells of these plants, the chromosomes remained spread in the cytoplasm in the first division without forming a metaphase plate. The abnormality blocked chromosome movements at anaphase I. Several micronuclei of different sizes were formed and, after the occurrence of an irregular first cytokinesis, the meiocytes progressed normally to the second division, generating polyads with unbalanced microspores. Pollen viability was not in correlation with meiotic abnormalities. The importance of these findings to
Brachiaria breeding program is discussed.
Key words: Brachiaria brizantha; breeding program; colchicine; forage grass; induced
Introduction
Many genera of the Poaceae family are economically important forage grasses. An example is the genus Brachiaria, native to the African savannas, that encompasses more
than 100 species (Keller-Grein et al., 1996). However, the commercially exploited brachiariagrasses around the world belong to four African species: B. brizantha
(palisadegrass), B. decumbens (signalgrass), B. humidicola (koroniviagrass), and B.
ruziziensis (ruzigrass). Their economic importance is greatest in tropical America where
extensive adoption over the past several decades has had a revolutionary impact on cattle productivity (Miles et al., 2004) placing Brazil as the largest beef exporter in the world (ABIEC, 2008). Although eight cultivars of Brachiaria are commercialized in
Brazil by a vital seed industry, seven of them were originated as direct selections from germplasm collected in Africa (Miles et al., 2004). The Brazilian Brachiaria breeding
program underway at Embrapa Beef Cattle aims at producing new cultivars by intra- and interspecific hybridization. However, Brachiaria hybridization is not easy because
of apomictic reproduction and differences in the ploidy level among species. In the genus Brachiaria diploidy is rare and correlated with sexual reproduction whereas
polyploidy and apomixis prevail (Valle and Savidan, 1996).
Brachiaria brizantha is one of the most important species of the genus and in
Brazil, one cultivar, cv. Marandu, covers more than 40 million hectares. Its widespread use is due to its high production, nutritive value and resistance to spittlebugs (Homoptera:Cercopidae). Resistance to this sucking insect was the major incentive for initiating brachiariagrasses breeding programs in America. Excellent resistance exists in
B. brizantha cv Marandu and several other palisadegrass accessions (Miles at al., 2004).
induced tetraploid accessions of B. ruziziensis (Valle and Savidan, 1996) since no
compatible sexual tetraploid has been found among B. brizantha accessions. Collection
trips done in eastern and southeastern tropical Africa in the mid-1980s by the International Center for Tropical Agriculture (CIAT, Colombia) resulted in the collection of more than 1000 distinct accessions of 33 species (Keller-Grein et al., 1996). Part of the CIAT germplasm of Brachiaria (475 accessions of 13 species) was
sent to Brazil and is maintained in the fields of Embrapa Beef Cattle Research Center (Campo Grande, MS). Analysis of flow cytometry revealed that among 437 accessions evaluated, 13% were diploid and the remaining were polyploid with ploidy ranging from 4n to 7n, and a large predominance of tetraploid accessions (Penteado et al., 2000). The B. brizantha collection, that encompasses 253 accessions, presents only a single
diploid accession (Miles and Valle, 1991; Valle and Savidan, 1996; Penteado et al., 2000).
To overcome the barrier between sexual diploid and apomictic tetraploid accessions, Ferguson and Crowder (1974) proposed to create a cross-compatible, sexual tetraploid plant by doubling the chromosome number. The barrier was overcome a decade latter, in Belgium, when diploid accessions of B. ruziziensis were tetraploidized
with colchicine (Gobbe et al., 1981; Swenne et al., 1981). These accessions continue to be the basis of the interspecific hybridization program both in CIAT and in Brazil. In attempt to overcome seed production failure in interspecific hybrids and also to broaden the genetic basis of Brachiaria breeding in Brazil, the diploid accession was submitted
to chromosome doubling by colchicine treatment applied in plants cultivated in vitro
(Pinheiro et al., 2000). Six tetraploid plants were obtained. Considering that polyploidy is largely correlated to abnormal meiosis in Brachiaria (Mendes-Bonato et al., 2002a;
fact that apomictic plants are pseudogamic, which means that fertile gametes are necessary for endosperm development, the current paper discusses the meiotic behavior in progenies of some induced tetraploidized plants as a subsidy to the selection of the best plants to be used as female genitors in intraspecific crosses.
Material and Methods
Chromosome doubling of the diploid sexual accession B105 (BRA002747) of B. brizantha was accomplished by colchicine treatment of basal segments of in vitro
grown plants at Cenargen/Embrapa, Brasilia-DF, by Pinheiro et al. (2000). Six induced tetraploid plants were obtained (C14, C31, C35, C36, C41, and C48). Progenies were derived from open-pollination in these plants. The sexual progenies derived from the induced tetraploid plants C31, C41, and C48 were cytologically analyzed. The mode of reproduction of each plant was previously determined by examination of embryo-sacs using interference contrast microscopy on methylsalicilate-cleared ovaries (Young et al., 1979). The progeny of C31 was represented by three plants; C41 by two plants; and C48 by a single plant. Chromosome countings in these plants were determined in well spread anaphase I.
carmine in fresh pollen grains of dehiscent anthers in some plants. Photomicrographs were made with a Wild Leitz microscope using Kodak Imagelink – HQ, ISO 25 black and white film.
Results and Discussion
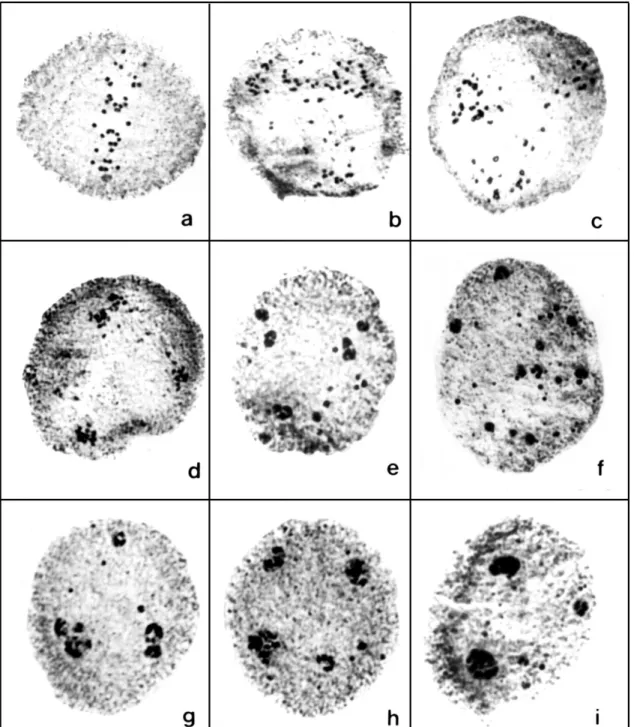
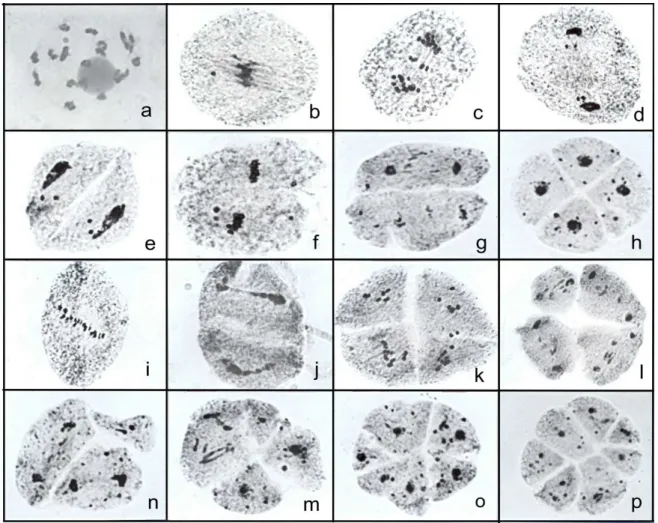
The chromosome countings at anaphase I in the six progenies of the tetraploidized plants (C31, C41, and C48) revealed the presence of 2n = 4x = 36 chromosomes in all of them (Fig. 1a). The results obtained in the analysis of the meiotic behavior and pollen sterility in the sexual induced tetraploidized plants are presented on Table 1. The meiotic behavior of artificially tetraploidized plants was very affected when compared with the original sexual diploid plant B105. The meiotic behavior in B105 was quite normal in the first division and totally normal in the second division, generating 100% of normal tetrads (Mendes-Bonato et al., 2002b). In contrast, the percentage of abnormal cells among the tetraploidized plants ranged from 57.1% to 60.3% in plants of the progeny of C31.
Figure 1. Aspects of meiocytes during the first division in the progenies of C31 and C41. a) Early anaphase I with 2n = 36 chromosomes after normal orientation in the metaphase plate observed in some cells. b and c) Meiocytes in the first division with chromosomes spread in the cytoplasm caused by chaotic spindle formation. d) Group of
Table 1. Number of cells analyzed, number and (percentage) of abnormal cells, and percentage of sterile pollen grains
Genotype
C31 C41 C48
Phase
Plant Plant Plant
1 3 4 1 2 1
M I 66 56 82 134 75 216
7 (10.6) 19 (33.9) 34 (41.5) 86 (64.2) 32 (42.7) 70 (32.4)
A I 39 28 16 66 48 51
17 (43.6) 7 (25.0) 4 (25.0) 27 (40.9) 17 (35.4) 5 (9.8)
T I 72 94 176 118 223 159
32 (44.4) 62 (66.0) 129 (73.3) 47 (39.8) 129 (57.8) 46 (28.9)
P II 140 134 48 212 106 119
102 (72.8) 86 (64.2) 31 (64.6) 54 (25.5) 39 (36.8) 32 (29.9)
M II 111 104 65 190 112 105
69 (62.2) 58 (55.8) 43 (66.1) 88 (46.3) 44 (39.3) 43 (40.9)
A II 70 21 37 50 60 78
43 (61.4) 12 (57.1) 16 (43.2) 18 (36.0) 26 (43.3) 46 (59.0)
T II 142 94 120 138 151 134
90 (63.4) 57 (60.6) 65 (54.2) 49 (35.5) 59 (39.1) 72 (53.7)
Tet 181 125 108 430 294 367
109 (60.2) 94 (75.2) 71 (65.7) 176 (40.9) 107 (36.4) 224 (61.0)
Total 821 656 652 1338 1069 1229
469 (57.1) 395 (60.2) 393 (60.3) 584 (43.6) 453 (42.4) 548 (44.6) Pollen
sterility 46.8 - - 31.1 29.4 11.6
* MI: metaphase I; AI: anaphase I; TI: telophase I; PII: prophase II; MII: metaphase II; AII: anaphase II; TII: telophase II, Tet: tetrade.
2005). Irregular chromosome segregation was also reported in induced tetraploid accessions of B. ruziziensis (Pagliarini et al., 2008). These abnormalities typical of
polyploids have also been reported for several Brachiaria species (Mendes-Bonato et
chromosome segregation, however, varies among accessions and species. The tetraploidized progenies from C31 and C41, on the other hand, had very abnormal meiosis. Only a few cells presented the chromosomes aligned in the metaphase plate or in the early anaphase (Fig. 1 a). In the majority of cells, the chromosomes were dispersed in the cytoplasm either as bivalent or univalent, without forming a metaphase plate (Fig. 1 b to d). Anaphase chromosome movements are blocked. As a consequence of this abnormality in chromosome orientation, a few or several micronuclei of different sizes were observed inside the cells (Fig. 1 e to i). A similar meiotic abnormality was reported in maize by Taschetto and Pagliarini (1993), where the meiotic mutant lacked spindle organization in both divisions, generating polyads as the meiotic product. After telophase, an abnormal cytokinesis occurred, dividing the cytoplasm in generally three parts with different amounts of chromosomes in each cell (Fig. 2 a to l). The second division progressed normally despite of the precocious cellularization. The result was the formation of polyads at the end of meiosis (Fig. 2 n to p). A few tetrads with micronuclei (Fig. 2m) were recorded in these plants. In artificially tetraploidized plants
of B. ruziziensis, some accessions also displayed non-typical meiotic behavior. In one of
them (R50), Risso-Pascotto et al. (2005a) reported a similar meiotic cellularization process compromising the meiotic product, and in another (R46), a meiotic mutation affecting spindle orientation in both divisions was recorded (Pagliarini et al., 2008).
The main objective of chromosome doubling in these sexual diploid plants was to obtain sexual tetraploids to be used as female genitors in intraspecific crosses with natural apomictic tetraploid pollen donors. To accomplish this objective, the meiotic process of tetraploidized plants must be regular to ensure pollen fertility, embryo fertilization and proper seed set. In five accessions of sexual tetraploidized plants of
(Pagliarini et al., 2008) while in two other accessions (R46 and R50), meiotic abnormalities were detected in more than 50% of the meiocytes (Risso-Pascotto et al., 2005b; Pagliarini et al., 2008). Despite these abnormalities, accession R50 was used in crosses with B. brizantha cv. Marandu, but some hybrids exhibited a high frequency of
meiotic abnormalities (Fuzinatto et al., 2007; Adamowski et al., 2008) and could not be advanced to cultivar development. On the other hand, crosses between those tetraploidized accessions of B. ruziziensis with a low frequency of meiotic
abnormalities, with different natural tetraploid accessions of B. brizantha (M. F.
Felismino, pers commun) revealed lesser problems, with abnormalities ranging from 18% to 55%. These results suggested that meiotic behavior in interspecific hybrids seems to be genotype-specific so it becomes very important to select genitors with none to low frequencies of abnormalities.
Intraspecific hybridization in B. brizantha is expected to yield hybrids with less
seed set problems than those observed so far in interspecific hybrids. The use of sexual tetraploidized plants such as C31 and C41 in crosses with natural apomictic B.
brizantha, or other compatible Brachiaria species, such as B. decumbens or B.
ruziziensis, need to take into account the results obtained in the present meiotic analysis
to avoid carry-over of these meiotic abnormalities into the progenies. Thus, considering the results obtained from the meiotic analysis, the sexual progeny C48 is more adequated to be used as female genitor in intra- and interspecific hybrids.
Pollen viability evaluated with propionic carmine was also performed in some plants in an attempt to find quicker means of selecting the best plants for crosses. Previous pollen viability tests in Brachiaria did not correlate well with the amount of
plants as may be seen on Table 1. Thus, the analysis of meiotic behavior still remains as the best parameter to select progenitors in Brachiaria breeding.
Acknowledgements
Authors are grateful to UNIPASTO for financial support.
References
ABIEC- Associação Brasileira das Indústrias Exportadoras de Carne. Site consulted on March 11, 2008. http/www.abiec.com.br/institutional.asp
Adamowski EV, Pagliarini MS, Valle CB, 2008. Meiotic behavior in three interspecific three way hybrids between Brachiaria ruziziensis and B. brizantha (Poaceae: Paniceae).
J Genet 87: 33-38.
Araujo ACG, Nóbrega JM, Pozzobon MT, Carneiro VTC, 2005. Evidence of sexuality in induced tetraploids of Brachiaria brizantha (Poaceae). Euphytica 144: 39-50.
Ferguson JE, Crowder LV, 1974. Cytology and breeding behaviour of Brachiaria ruziziensi. Crop Sci 14: 893-895.
Fuzinatto VA, Pagliarini MS, Valle CB, 2007. Microsporogenesis in sexual Brachiaria
Gobbe J, Swenne A, Louant BP, 1981. Diploïdes naturels et autotétraploïdes induits chez Brachiaria ruziziensis Germain et Evrard: critères d’identification. Agron Trop 36:
339-346.
Keller-Grein G, Maass BL, Hanson J, 1996. Natural variation in Brachiaria and
existing germoplasm collections. In: Miles JW, Maass BL, Valle CB, eds. Brachiaria:
Biology, Agronomy, and Improvement. Colombia: CIAT/Embrapa: 17-42.
Mendes-Bonato AB, Junqueira Filho RG, Pagliarini MS, Valle CB, Penteado MIO, 2002a. Unusual cytological patterns of microsporogenesis in Brachiaria decumbens:
abnormalities in spindle and defective cytokinesis causing precocious cellularization. Cell Biol Int 26: 641-646.
Mendes-Bonato AB, Pagliarini MS, Forli F, Valle CB, Penteado MIO, 2002b. Chromosome number and microsporogenesis in Brachiaria brizantha (Gramineae).
Euphytica 125: 419-425.
Mendes-Bonato AB, Pagliarini MS, Risso-Pascotto C, Valle CB, 2006. Chromosome number and meiotic behavior in Brachiaria jubata (Gramineae). J Genet 85: 83-87.
Miles JW, Valle CB, 1991. Assessment of reproductive behavior of interspecific Brachiaria hybrids. Apomixis Newslett 3: 9-10.
Miles JW, Valle CB, Rao IM, Euclides VPB, 2004. Brachiariagrasses. In: Sollenberger
L. editor. Warm-Season (C4) Grasses Madison: ASA, CSSA, SSSA: 745-760.
Penteado MIO, Santos ACM, Rodrigues IF, Valle CB, Seixas MAC, Esteves A, 2000. Determinação de poliploidia e avaliação da quantidade de DNA total em diferentes espécies de Brachiaria. Boletim de Pesquisa No. 11. Campo Grande-MS, Embrapa
Gado de Corte 19p.
Pinheiro AA, Pozzobon MT, Valle CB, Penteado MIO, Carneiro VTC, 2000. Duplication of the chromosome number of diploid Brachiaria brizantha plants using
colchicine. Plant Cell Rep 19: 274-278.
Risso-Pascotto C, Pagliarini MS, Valle CB, 2005a. Multiple spindles and cellularization during microsporogenesis in an artificially induced tetraploid accession of Brachiaria
ruziziensis (Gramineae). Plant Cell Rep 23: 522-527.
Risso-Pascotto C, Pagliarini MS, Valle CB, 2005b. Meiotic behavior in interspecific hybrids between Brachiaria ruziziensis and Brachiaria brizantha (Poaceae). Euphytica
145:155-159.
Risso-Pascotto C, Pagliarini MS, Valle CB, 2006. Microsporogenesis in Brachiaria
dictyoneura (Fig. & De Not.) Stapf (Poaceae: Paniceae). Genet Mol Res 5: 837-845.
Swenne A, Louant BP, Dujardin M, 1981. Induction par la colchicine de formes autotétraploïdes chez Brachiaria ruziziensis Germain et Evrard (Graminée). Agron Trop
36:134-141.
Utsunomiya KS, Pagliarini MS, Valle CB, 2005. Microsporogenesis in tetraploid accessions of Brachiaria nigropedata (Ficalho & Hiern) Stapf (Gramineae). Biocell
29:295-301.
Valle CB, Savidan YH, 1996. Genetics, cytogenetics, and reproductive biology of
Brachiaria. In: Miles JW, Maass BL and Valle CB, editors. Brachiaria: Biology,
Agronomy, and Improvement, Colombia: CIAT/Embrapa: 147-163.
Analysis of meiotic behavior in selecting potential
genitors among diploid and artificially induced
tetraploid accessions of
Brachiaria ruziziensis
(Poaceae)
Alice Maria de Souza-Kaneshima
1, Claudicéia Risso-Pascotto
1, Maria
Suely Pagliarini
1*, and Cacilda Borges do Valle
21. Department of Cell Biology and Genetics, State University of Maringá,
87020-900 Maringá PR Brazil.
2. Embrapa Beef Cattle, P.O. Box 154, 79002-970 Campo Grande MS
Brazil.
* Author for correspondence: Maria Suely Pagliarini (E-mail:
[email protected]
).
RUNNING TITLE
:
Microsporogenesis in Brachiaria ruziziensis 2n and 4nADDRESS FOR CORRESPONDENCE
Dra. Maria Suely Pagliarini
Department of Cell Biology and Genetics
State University of Maringá
87020-900 Maringá, Paraná - Brazil.
Analysis of meiotic behavior in selecting potential
genitors among diploid and artificially induced
tetraploid accessions of
Brachiaria ruziziensis
(Poaceae)
Abstract.
Sexuality is correlated with diploidy and apomixis with polyploidy in theBrachiaria genus. Brachiaria ruziziensis is a key species in Brachiaria breeding due to
its obligate sexuality and intrinsic agronomic qualities. Interspecific crosses in the genus became feasible only when a few diploid accessions of B. ruziziensis were artificially
tetraploidized and remained sexual. Hybridization has been done since, using natural tetraploid apomictic accessions of B. brizantha or B. decumbens as pollen donors.
Twenty two accessions of B. ruziziensis from the Embrapa Beef Cattle germplasm
collection (Campo Grande, MS, Brazil) were cytologically analyzed: 16 are natural diploids (2n = 2x = 18) and six are artificially induced tetraploids (2n = 4x = 36). The meiotic behavior in the 16 diploid accessions varied. The mean of meiotic abnormalities per accession ranged from zero to 24.46%. Meiotic behavior in the induced tetraploid accessions also varied with the mean of meiotic abnormalities ranging from 5.20% to 54.71%. The most common abnormalities observed in both the diploid and the tetraploid accessions, were those related to irregular chromosome segregation. In one tetraploid accession, with a high frequency of those, other irregularities involving chromosome orientation at metaphase plate and chromosome convergence to the poles, a meiotic mutation known as divergent spindle, were recorded. Meiotic behavior should
be considered in selecting potential genitors for breeding.
Key words: Brachiaria ruziziensis, microsporogenesis, diploidy, tetraploidy, forage
Introduction
Some African species of Brachiaria have been introduced into the Americas and
became the most important forage for pastures in the tropics. Accessions of five species
– B. brizantha, B. decumbens, B. dictyoneura, B. humidicola, and B. ruziziensis – have
been released as commercial cultivars in different countries, including Brazil (Keller-Grein et al. 1996). New cultivars can be obtained either from direct selections from the natural existing variability in the germplasm collections or from interspecific hybridizations. There are seven germplasm collections of Brachiaria in ex situ
conservation in the world, encompassing almost 1000 distinct accessions of 33 species (Keller-Grein et al. 1996). In Brazil, the Brachiaria collection is maintained by
Embrapa Beef Cattle, at Campo Grande, MS, and encompasses 475 accessions of 13 species. Brachiaria ruziziensis is represented by 25 accessions in that collection (Valle
and Savidan 1996). It is native to the region of the Ruzi Valley between the Democratic Republic of Congo, Rwanda and spreads into Burundi. The cultivar Kennedy is widely distributed in tropical countries. It is known as ruzigrass or Congo signalgrass. Ruzigrass provides palatable forage of high nutritive quality. It requires fertile, well-drained soils and it is highly susceptible to most important insect pest in pastures, the spittlebugs (Keller-Grein et al. 1996). It flowers in the autumn and produces good seed crops.
Polyploidy is predominant in the genus Brachiaria. Analysis by flow cytometry
revealed that among 437 accessions of Brachiaria of the Embrapa Beef Cattle collection