UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE QUÍMICA E BIOQUÍMICA
Study of pH-dependent misfolding of surfactant protein C
using state-of-the-art molecular modelling techniques
Catarina Azevedo Carvalheda dos Santos
Mestrado em Bioquímica
UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE QUÍMICA E BIOQUÍMICA
Study of the pH-dependent misfolding of
surfactant protein C using state-of-the-art
molecular modelling techniques
Catarina Azevedo Carvalheda dos Santos
Mestrado em Bioquímica
Dissertação de Tese de Mestrado orientada por
Doutor António M. Baptista e Doutor Miguel Machuqueiro
Lisboa
2011
“A field plays out when our imagination fails and we can no longer think of good questions to ask.”
vii
Abstract
P-C is a highly hydrophobic protein with two covalently linked fatty acyl chains that adopts a mainly helix structure while associated with the membrane. However, it misfolds into a β-rich structure under certain environmental conditions, which suggests that SP-C is in a metastable state. The slow α→β transitions are likely to be caused by the membrane-dissociation, deacylation and exposure to the neutral pH of the alveolar subphase, leading to the formation of amyloid aggregates associated with Pulmonary Alveolar Proteinosis (PAP).
Here, we present the first study of the pH-dependent conformational behaviour of the deacylated SP-C (dSP-C) in a chloroform/methanol/water mixture using state-of-the-art molecular modelling methods, with the aim of getting insight into several structural properties of dSP-C that might be relevant for the formation of amyloid fibrils in PAP. We performed 60 simulations of 100 ns each, considering 6 different pH conditions and using the GROMOS 43A1 and the recently released GROMOS 54A7 force fields.
Although the results for both force fields are contradictory and neither of them reproduces the experimental results for the dSP-C under similar conditions, they identify the increase of pH as a promoter of protein-protein interactions, in this type of organic mixtures. The possibility of undergoing inter-protein interactions due to the presence of other dSP-C molecules may be the principal contributor factor for the α→β transitions observed at pH 8 in the experimental results and that were not captured in our simulations.
Our simulations show that the loss of structure is more prone to begin at the terminal region and that the β motifs start to form simultaneously in both N and C-terminal regions of the dSP-C with their approximation.
These observations reinforce the necessity of using more than one force field to address a scientific question and validate the newest GROMOS 96 version, the 54A7.
Keywords: SP-C, misfolding, constant-pH molecular dynamics, organic solvent
mixtures, pH
viii
Resumo
proteína C do surfactante pulmonar (SP-C) é um dos constituintes proteicos do surfactante pulmonar, uma interface lipídica que delimita os alvéolos pulmonares e que tem como função manter uma baixa tensão superficial durante a inspiração e prevenir o colapso alveolar durante a expiração. A SP-C é particularmente importante para uma expansão eficaz da interface aquando do aumento do volume pulmonar e para a estabilização das reservas de surfactante aquando da diminuição do volume pulmonar.
A SP-C é constituída por 35 aminoácidos e possui uma região estruturalmente desordenada no segmento N-terminal (1-8), uma região em hélice rica em resíduos de aminoácidos de cadeias laterais apolares (9-34), um resíduo C-terminal sem estrutura definida e dois grupos acilo ligados covalentemente a dois resíduos de cisteína nas posições 5 e 6. A proteína adopta uma orientação transmembranar que lhe permite permanecer ancorada ao surfactante pulmonar durante os períodos de expansão e compressão do ciclo respiratório.
A Proteinose Alveolar Pulmonar (PAP) é uma doença amilóide caracterizada pela formação de agregados de material do surfactante nos pulmões, estando associada a disfunções respiratórias. 90% dos casos de PAP estão associados à forma idiopática, para a qual não se conhecem ainda causas. Contudo, análises ao material agregado revelaram quantidades significativas de SP-C desacilada características de doentes com PAP [45].
Embora estável em ambiente membranar, a SP-C é metaestável em solução, sofrendo alterações conformacionais que resultam na perda da estrutura em hélice e no ganho de motivos β, culminando na formação de fibrilhas amilóides. A remoção dos grupos acilo da proteína leva a uma perda de cerca de 20% do conteúdo total em hélice num ambiente membranar a pH neutro [32, 62] e promove a sua dissociação da membrana [56], sendo um factor contributivo para a exposição da proteína a um ambiente aquoso. Para além destes factores, variações de pH parecem influenciar a estabilidade estrutural da forma desacilada em misturas de clorofórmio, metanol e
ix
água [56]. Todos estes factores afectam a estabilidade da SP-C conduzindo à formação de agregados fibrilhares característicos da PAP.
Dado o seu carácter extremamente hidrófobo e rápida formação de agregados proteicos quando em solução, a SP-C é difícil de estudar experimentalmente. A sua estabilidade estrutural é especialmente sensível ao ambiente químico que a rodeia, sendo afectada pelas interacções proteína-solvente e pelo arranjo das moléculas de solvente à sua volta. Estas características tornam a SP-C num candidato perfeito para estudar usando técnicas de simulação molecular que permitem o estudo conformacional da proteína na ausência de outras moléculas de SP-C a um nível atómico.
Existe um grande número de técnicas computacionais que podem ser aplicadas ao estudo de sistemas biomoleculares e que permitem analisar diferentes aspectos do comportamento molecular. De particular interesse para o trabalho desenvolvido durante a tese de mestrado são os métodos de Mecânica Molecular (MM) e de Electrostática Contínua (EC) utilizados para os cálculos energéticos associados a diferentes estados do sistema que podem ser amostrados através de métodos de Dinâmica Molecular (DM) e de Monte Carlo (MC). Quando conjugados, os métodos de MM/DM permitem seguir a dinâmica conformacional do sistema a nível atómico e os métodos de EC/MC permitem amostrar diferentes estados de ionização para um sistema rígido. Estas metodologias podem ser usadas para estudar o efeito do pH em proteínas mas ambas apresentam desvantagens significativas: os métodos de MM/MD amostram o espaço conformacional mas tendo em conta um estado de protonação fixo, enquanto os métodos de EC/MC amostram diferentes estados de protonação mas para uma estrutura rígida. No primeiro caso perdemos a amostragem de diferentes estados de protonação e no segundo caso não incluímos alterações conformacionais que possam influenciar ou ser influenciadas por um novo estado de protonação atribuído ao sistema (acoplamento conformação-protonação).
Dada a importância do pH para a estrutura e função de proteínas, o avanço na área da simulação molecular contou com um esforço para ultrapassar as desvantagens inerentes aos métodos disponíveis para estudar efeitos associados ao pH. Este esforço
x
resultou na criação dos métodos de dinâmica molecular a pH constante que introduzem o pH como um parâmetro externo que é controlado ao longo do período de simulação (como a temperatura e a pressão). Vários destes métodos resultam da complementaridade entre métodos de MM/DM e EC/MC, superando as suas desvantagens: simulações MM/DM amostram o estado conformacional para um dado estado de protonação que é alterado com uma frequência pré-definida através de cálculos EC/MC. Desta forma, é possível seguir o comportamento estrutural do sistema tendo em conta os diferentes estados de protonação acessíveis a um dado pH e tendo em conta o acoplamento conformação-protonação. De entre os métodos de dinâmica molecular a pH constante que usam métodos de MM/DM e de EC/MC, o método de titulação estocástica do grupo de Simulação Molecular do ITQB [107, 128, 129] apresenta-se como a alternativa mais realista, incluindo o efeito da configuração das moléculas do solvente na dinâmica e o efeito da força iónica do meio no cálculo da probabilidade de um estado de protonação.
Dada a importância dos efeitos do pH e da organização e natureza das moléculas do solvente para a estabilidade estrutural da SP-C desacilada, propusemo-nos a estudar o seu comportamento conformacional numa gama de valores de pH de 3 a 8, considerando uma mistura de solvente orgânico que visa mimetizar o ambiente membranar e utilizando o método de titulação estocástica de dinâmica molecular a pH constante. Este estudo tem como objectivo compreender as propriedades que podem ser relevantes para a perda de estrutura da SP-C e consequente formação das fibras amilóides características da PAP.
Para tal foram realizadas um total de 60 simulações de 100 ns cada, com a SP-C desacilada numa mistura de clorofórmio, metanol e água, semelhante à usada na determinação da sua estrutura com técnicas de Ressonância Magnética Nuclear [26]. Foram utilizadas 5 estruturas iniciais diferentes, simuladas a 6 valores de pH (3, 4, 5, 6, 7 e 8) e utilizando 2 campos de forças alternativos da família GROMOS96, o 43A1 e, a mais recente versão desta família, o 54A7, para o cálculo da energia potencial nos métodos de MM/DM. A utilização de dois campos de forças deve-se ao facto de evidências recentes apontarem para a tendência de alguns dos mais utilizados campos de forças estabilizarem determinados motivos de estrutura secundária em detrimento
xi
de outros, tornando a escolha do campo de forças dependente do sistema que se pretende simular.
Os resultados obtidos para a estabilidade estrutural da SP-C desacilada são contraditórios: a região da hélice é mais estável a pH 5, 6 e 7 com a versão 43A1 e a pH 3 e 8 com a versão 54A7. Para além disso, a formação de estrutura β é apenas significativa nos resultados obtidos com a versão 43A1 a valores de pH acídicos. Nenhum dos resultados está, contudo, de acordo com os resultados experimentais obtido para a SP-C desacilada numa mistura de solvente orgânico semelhante, em que a proteína mantém uma estrutura em α-hélice a pH baixo (≈2), semelhante à estrutura nativa a pH neutro, que é perdida para uma estrutura rica em motivos β com o aumento progressivo do pH até aproximadamente 8 [56].
No entanto, a discrepância entre os nossos resultados e os dados experimentais pode advir de dois factores. Em primeiro lugar, do uso de diferentes proporções de metanol/clorofórmio nas misturas de solvente adoptadas nos dois trabalhos. Neste tipo de solventes existe normalmente uma preferência de solvatação das moléculas de metanol em relação a regiões polares e das moléculas de clorofórmio em relação a regiões apolares [154, 155], que é observada nas nossas análises às interacções proteína-solvente. Este efeito torna a proporção clorofórmio/metanol importante para garantir que a quantidade de moléculas de clorofórmio em solução é suficiente para manter a estabilidade da região em hélice da SP-C desacilada. Nas nossas simulações a proporção clorofórmio/metanol é cerca de 0.5 em termos de volume (tal como na mistura de solvente utilizada para obter a estrutura da SP-C), enquanto no estudo experimental em que a titulação da SP-C desacilada foi estudada a proporção clorofórmio/metanol é de, aproximadamente, 0.14 também em termos de volume. Em segundo lugar, a estabilidade da SP-C desacilada no trabalho experimental pode também estar a ser influenciada pela presença de outras moléculas de SP-C em solução. Ao longo das nossas simulações foi possível observar interacções intra-proteína de natureza electrostática que resultarão da incapacidade das moléculas de solvente para estabilizar a carga dos resíduos tituláveis, tornando interacções proteína-proteína mais favoráveis em relação a interacções proteína-solvente. Algumas destas interacções tornam-se mais declaradas com o aumento do pH,
xii
causando desvios significativos nas curvas de titulação obtidas por nós para alguns dos resíduos tituláveis. Este efeito é normalmente observado em solventes orgânicos e poderá estar a promover interacções inter-proteína e, consequentemente, as transições α→β observadas para valores mais elevados de pH no estudo experimental em causa. Tendo em conta os resultados obtidos para o campo de forças 54A7, esta alteração conformacional para uma estrutura rica em motivos β a pH elevado é a única particularidade que não está a ser captada nas nossas simulações, provavelmente devido à inexistência de outras moléculas de SP-C nos sistemas simulados. A comparação de ambos os estudos, identifica o pH como um factor contributivo para a formação das fibrilas amilóides a partir da SP-C desacilada na mistura de solvente orgânico considerada, mas demonstra que este não será suficiente para promover as transições α→β que antecedem a formação dos agregados pré-fibrilares.
Embora a falta de mais estudos experimentais não permita uma análise mais aprofundada das discrepâncias entre os nossos resultados e os resultados obtidos experimentalmente, abre as portas a uma possível via alternativa de investigação em que a presença de várias moléculas de SP-C desacilada poderá ser estudada, considerando a nossa mistura de solvente, por métodos de simulação molecular.
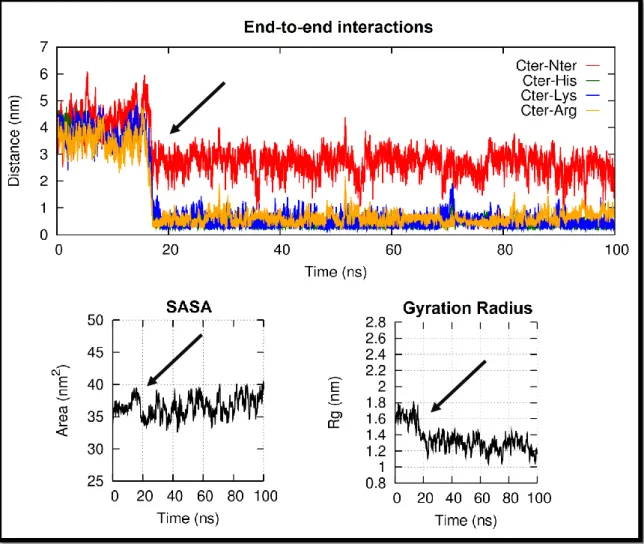
Algumas das interacções electrostáticas observadas nas nossas simulações envolvem resíduos das regiões extremas da proteína, fazendo com que esta se dobre sobre si própria e promovendo, assim, uma diminuição da área superficial acessível ao solvente da proteína e do raio de giração da proteína, observados na maior parte dos replicados simulados com o campo de forças 43A1 e nalguns replicados simulados com o campo de forças 54A7.
Para ambos os campos de forças utilizados, a perda de estrutura em hélice começa, na maior parte dos casos, na região C-terminal e a formação de motivos β acontece em simultâneo nas regiões N e C-terminal, podendo ter um carácter persistente ou transitório.
O nosso estudo é inovador em três principais aspectos: consiste nas primeiras simulações longas da SP-C desacilada, nas primeiras simulações em que esta proteína é simulada numa mistura de solvente orgânico e na primeira vez em que é amostrado
xiii
não só o seu espaço conformacional como diferentes estados de protonação acessíveis a diferentes valores de pH. Em particular, o uso da mistura de solvente em causa permitiu identificar alguns aspectos estruturais que se relacionam com a utilização de solventes orgânicos e que podem afectar a estabilidade e o comportamento estrutural do sistema em estudo, e a utilização de dois campos de força permitiu reforçar a ideia da necessidade de usar diferentes conjuntos de parâmetros quando pretendemos estudar o comportamento conformacional de um sistema usando técnicas de simulação molecular e permitiu validar a mais recente versão dos campos de força da família GROMOS96, o 54A7.
O facto de não termos obtido resultados semelhantes aos do estudo experimental da SP-C desacilada numa mistura de solvente orgânico semelhante à por nós utilizada, permitiu-nos identificar a presença de outras moléculas de SP-C como factor adicional que poderá ser relevante para a ocorrência de transições α→β e consequente formação das fibrilhas amilóides características da PAP, embora este factor possa ser apenas uma consequência da utilização de uma mistura de solvente orgânico para mimetizar o ambiente membranar.
De forma a compreender o efeito estabilizador da presença dos grupos acilo na SP-C estamos, neste momento, a realizar um estudo semelhante para a SP-C acilada, considerando a mesma mistura de solvente orgânico, os mesmos campos de força e o mesmo intervalo de valores de pH. Posteriormente pretendemos simular a SP-C acilada e desacilada num ambiente membranar sob condições de pH similares e utilizando os mesmos campos de força. Estes últimos estudos serão os mais relevantes para compreender o efeito da perda dos grupos acilo e de ligeiras variações de pH no comportamento conformacional da SP-C in vivo e permitirão responder a algumas das questões deixadas em aberto pelo trabalho decorrido durante a tese de mestrado.
Palavras-chave: SP-C, misfolding, dinâmica molecular a pH constante, misturas de
xiv
Acknowledgements
t all started with my growing interest in the use of informatic tools to study scientific problems and in the subject of misfolding diseases, which were combined in the master thesis proposed by Dr. António Baptista and Dr. Sara Campos and led me to integrate the Molecular Simulation group at ITQB. By that time I did not know what to expect, but now I can say that I feel lucky and grateful to have been given this opportunity. The work was always stimulating, the people were always friendly and helpful and the work environment was always informal and pleasant.
Starting with the main people responsible for this great experience, I would like to thank Dr. António Baptista and Dr. Sara Campos for their guidance, support, time and for all the things they taught me during this year. In particular, I want to thank António for his constructive criticism, correctness and rigor and Sara for all her attention, devotion and advice, which were major contributors for this work from the beginning.
I would also like to acknowledge Dr. Miguel Machuqueiro, whom I was lucky to have as co-supervisor, for his sympathy, support and availability to help and discuss the work developed during this master thesis. I am also grateful for his contributions for the preparation of this work.
I cannot forget to thank my colleagues in the Molecular Simulation Group and everyone in the Protein Modelling group, for being an important part of this experience and for all the good moments, fruitful discussions, help and fellowship.
I want to thank my parents for their moral and financial support without which this would not have been possible to accomplish, and for always believe in my capabilities and support my decisions.
Last but not the least, I want to thank all my family and friends for their support. In particular, I want to acknowledge two unique human beings that always kept me mentally sane in the most stressful moments. Bruno Narciso for all the love
xv
and patience and for not letting me sink into my worries and fears, and Inês Torcato for all her time, attention and care and for keeping me always informed and updated.
xvii
Index
Abstract ... vii Resumo ... viii Acknowledgements ... xiv 1. Surfactant Protein C ... 1 1.1. Lung Surfactant ... 1 1.1.1. The discovery ... 2 1.1.2. Synthesis ... 41.1.3. Functions and properties ... 6
1.2. Surfactant Protein C: Biochemistry ... 10
1.2.1. Biosynthesis and sorting ... 10
1.2.2. Structure ... 12
1.2.3. Transmembranar structure ... 15
1.2.4. Functions ... 16
1.3. pH-dependent misfolding of SP-C ... 17
1.3.1. Pulmonary Alveolar Proteinosis ... 19
1.3.2. The stability of SP-C structure ... 21
1.4. Studying the Misfolding of SP-C by Molecular Modelling Methods ... 24
2. Molecular Simulation ... 25
2.1. Statistical Mechanics – From microscopic data to macroscopic properties ... 26
2.1.1. The Ergodic Hypothesis ... 27
2.2. Periodic Boundary Conditions ... 29
2.3. Molecular Mechanics and Force Fields... 30
2.3.1. Empirical Potential Energy Functions ... 31
xviii
2.3.3. Some force field families for protein modelling ... 37
2.3.4. Choosing an appropriate force field ... 39
2.4. Molecular Mechanics/Molecular Dynamics ... 40
2.4.1. How does it work? ... 40
2.4.2. Choosing an appropriate time step for integration ... 41
2.4.3. The Lyapunov-instability... 42
2.5. Continuum electrostatics (CE) methods ... 42
2.5.1. Continuum electrostatics and Poisson Boltzmann Equation ... 44
2.5.2. Acid-base equilibrium in proteins using PB methods ... 47
2.6. Poisson-Boltzmann/Monte Carlo (PB/MC) ... 49
2.6.1. Monte Carlo methods... 49
2.6.2. PB/MC methods, how does it work? ... 50
2.7. Constant pH Molecular Dynamics ... 51
2.7.1. MM/MD or PB/MC methods? Both! ... 51
2.7.2. The stochastic titration method ... 52
3. Methods... 55
3.1. Box Shape ... 55
3.2. Constraints vs Restraints... 55
3.3. Non-bonded Interactions ... 56
3.4. Energy minimisation and search algorithms ... 57
3.4.1. Steepest Descent ... 57
3.4.2. L-BFGS ... 58
3.5. Temperature and pressure control: NPT ensemble ... 60
3.6. The procedure ... 62
3.6.1. Preparation phase ... 62
xix
3.6.3. Production phase ... 68
3.7. General Analysis ... 73
3.7.1. Protein analysis ... 73
3.7.2. Solvent analysis ... 76
3.7.3. Plots and images ... 77
4. Protein analysis: results and discussion ... 79
4.1. Global Structural Analysis ... 79
4.1.1. The stability of the helix structure ... 79
4.1.2. Hydrogen bonds ... 91
4.1.3. Root Mean Square Deviation ... 92
4.1.4. Solvent Accessible Surface Area ... 93
4.1.5. Gyration Radius ... 95
4.2. The missing factor: end to end interactions ... 96
4.3. Where does the loss of structure occur? ... 103
4.4. Titration Behaviour ... 108
4.4.1. Global Titration ... 109
4.4.2. Titration of the individual groups ... 110
4.5. Residue-Residue Interactions and Titration Shifts ... 113
4.5.1. Cysteine interactions ... 113
4.5.2. Histidine interactions... 122
4.5.3. Intra-protein interactions and organic solvent mixture ... 128
4.6. Summing up ... 129
5. Protein – Solvent interactions: results and discussion ... 133
5.1. Radial distribution function ... 135
5.1.1. Histidine 9 ... 137
xx
5.1.3. Valine residues 17 and 26 ... 142
5.2. Contacts persistency ... 144
5.2.1. Summing up ... 147
6. Concluding Remarks ... 149
6.1. Conclusions ... 149
6.2. Ongoing work and future perspectives... 150
Appendix I: Solvent mixture calculations ... 151
Proportion of solvent molecules ... 151
Dielectric constant calculation ... 153
Isothermal Compressibility calculation ... 155
Methanol and Chloroform Hydration Shells For the Contacts Persistency ... 156
Appendix II: Structural Content Analysis ... 159
Dssp ... 159
Root Mean Square Deviation (RMSD) ... 219
Solvent Accessible Surface Area (SASA) ... 226
Hydrogen Bonds ... 233
Gyration Radius (Rɡ) ... 240
Root Mean Square Fluctuation (RMSF) ... 247
Appendix III: Residue-Residue interactions ... 255
End-to-End Interactions ... 255
Histidine interactions... 262
1
1. Surfactant Protein C
1.1. L
UNGS
URFACTANTir breathing consists, generally, in the reciprocal movement of air into and out of the lungs. The vertebrate lungs are complex networks of tubes and sacs, which structure varies considerably among species. In the case of mammals, the trachea subdivides to form bronchi, which branch repeatedly in bronchioles, each of which connected to blind-ending, interconnected sacs – alveoli – that form the termination point of the respiratory tract [1] – Figure 1.
Figure 1 – The bronchial tree. Adapted from [2].
The alveolar system of the lung is the largest surface of the body in contact with the environment and it is composed of two types of epithelial cells, pneumocytes I and II, and macrophages [3] – Figure 2. The alveoli are tiny respiratory exchange units that allow for the generation of an enormous respiratory surface area, maximizing the amount of gas exchange during the breathing cycle – Figure 2.
2
Figure 2 – Alveolar cellular components. Adapted from [2].
The entire alveolar surface is lined with a thin fluid continuum, called the alveolar lining layer, which consists of an aqueous hypophase covered by a monolayer of pulmonary surfactant material that creates an air/water interface in the lungs [4]. This air/water surface film is formed at the first air inspiration of the newborn and it is the site of surfactant’s major action on lung mechanics [5] being responsible for biophysical stabilizing activities and innate defence mechanisms [6, 7].
1.1.1. The discovery
For over a century it was taught that the breathing cycle consisted in an alternated balance between the work of the respiratory muscles to expand the alveoli and the elastic recoil of the chest wall and lung tissue to contract it [8]. It was only in the 1920s that Kurt von Neergaard questioned the correctness of this simple concept and related the surface tension with the lung’s work [9].
The surface tension is the force per unit length in the plane of the interface between a liquid and a gas, which has the tendency to contract due to cohesive forces between the molecules in and near that liquid surface [8, 10]. According to Laplace’s Law and considering a curved surface, like the alveoli surface, the pressure difference
3
across the surface is directly proportional to the tension and inversely proportional to the radius of curvature, which means that the surface tension raises the pressure inside the alveoli and that the smaller the alveoli radius the higher the pressure inside it [8]. Any substance that acts on lowering the surface tension of a liquid surface is called a surfactant [8, 10].
The properties, function and origin of the alveolar lining layer were later described by Richard Pattle in 1955 [11], who suggested that the airspaces are lined by an insoluble layer composed of lipid-protein complexes that eliminates the surface tension in the alveoli lining [8, 12]. In 1957, and after making some changes in the design of the Langmuir-Wilhelmy balance1, John Clements published the first direct surface tension measurements on lung extracts [8, 12, 13]. In 1958, Mary Avery and Jere Mead established the connection between neonatal respiratory distress syndrome and a deficiency of surface active material, proving its clinical relevance [8].
Further work was done to get insight into this surface film composition. In the 1960s the important role of dipalmitoyl phosphatidilcholine (DPPC) in lowering the surface tension was described by Klaus and co-workers and its synthesis and secretion into the alveolar space by Gluck and co-workers [12]. The first indication for the presence of proteins in the surfactant film came in the 1970s from Robert King and co-workers [14], but it was only with the first successful treatment of infants with respiratory distress syndrome using surfactant replacement therapy that the importance of the surfactant proteins for surfactant efficiency was accepted [12]. Since then more attention has been given to surfactant proteins.
1
One of the oldest but still most used devices to study the spread of films of different compositions that consists of a trough containing the aqueous hypophase whose surface area can be varied, and a Wilhelmy dipping plate attached to an electro-balance which permits to deposit precise amounts of surface active material [6].
4
1.1.2. Synthesis
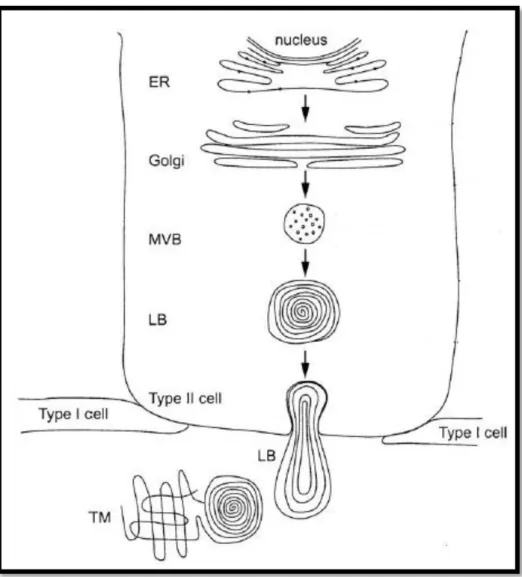
The pulmonary surfactant is a complex mixture of phospholipids, neutral lipids and proteins that is assembled in the endoplasmatic reticulum and the Golgi apparatus of alveolar pneumocyte II cells in the form of tightly packed membranes, which are stored in specialized organelles called lamellar bodies2 (LB) until exocytosis [7, 15].
After lamellar bodies being released into the alveolar subphase, the surfactant complexes experiment different environmental conditions: increase in hydration, change in calcium ion concentration and an increase in pH. All these changes seem to trigger the unpacking of lamellar bodies and their transformation into a characteristic cross-hatched structure termed tubular myelin (TM) [16]. The TM then supplies the material for surface film, transferring lipids and proteins into the alveolar interface. Two cartoon illustrations of this process are depicted in Figure 3 and Figure 4.
2
Dense proteinaceous core with lipid bilayers arranged concentric, stacked lamellae surrounded by a limiting membrane.
5
Figure 3 - Surfactant synthesis. ER: endoplasmatic reticulum; MVB: multivesicular body; LB: lamellar body; TM: tubular myelin. Adapted from [17].
The exact mechanism by which the lipids and proteins are transferred to the interface is not known. Still, there is evidence suggesting that the surfactant components are transferred from TM to incorporate the air-water interface by rapid adsorption [4]. The TM has been considered to be associated with highly surface active material as a precursor, a reservoir, or both, for the surface film [16] – Figure 4.
6
Figure 4 – Transference of surfactant to the surface. Adapted from [16].
Electrostatic interactions and perturbations in the phospholipid packing are believed to contribute to the insertion of pulmonary surfactant components into the surface by enhancing protein-lipid interactions and the exchange of molecules between the TM and the interface [16].
Once in the interface, the surfactant is repeatedly expanded and compressed during the breathing cycle and, after completed their mission, the surfactant components are removed from the surface film and taken up by the epithelium type II cells or degraded by alveolar macrophages [18].
1.1.3. Functions and properties
The main functions of the pulmonary surfactant are overcoming the problems in the respiratory mechanics due to the volume variations, allowing the dynamic film behaviour and reducing the work of breathing during the respiratory cycle. When inspiration takes place, the expansion of the lungs demands a great increase in the pulmonary surface that is achieved with the spreading of the air/water interface as a
7
result of the inclusion of more surfactant material. This lowers the surface tension and reduces the energy required to inflate the lungs, increasing the lungs compliance3. During the expiration some of the lipids and proteins are excluded from the surface film to surfactant reservoir bilayers, decreasing the elastic recoil and thereby preventing the alveolar system collapse due to the increase of the surface pressure. The material excluded upon compression stays attached to the interface preventing the loss of material to the aqueous subphase [4, 19, 20].
To accomplish these functions the pulmonary surfactant has to possess, at least, three biophysical properties [4, 6]:
i. Rapid adsorption of surfactant material into the surface film during inspiration;
ii. Effective film replenishment upon film expansion; iii. Very low surface tension upon film compression.
The ability to lower the surface tension with changing surface area is attributed to the interactions between surfactant components: lipid-lipid and lipid-protein interactions.
In addition, due to its continuous exposure to the environment, lungs have specific and non-specific defence mechanisms capable of eliminating microorganisms before they cause a disease and some of them involve surfactant components [6].
Surfactant Lipids
The surfactant lipids consist mainly of phospholipids (PL, ≈90-95%) with a small amount of neutral lipids (≈5-10%), primarily cholesterol. The most prevalent phospholipid class is phosphatidylcholine (PC), primarily dipalmitoylphosphatidil-choline (DPPC, ≈36%) which contributes largely to surface tension reduction, and unsaturated PC (≈33%) which can provide the required fluidity during the dynamic breathing process [4, 16, 21]. Besides PC, there are anionic lipids, mainly phosphatidilglycerol (PG, ≈10%), which seem to be important for lipid-protein interactions [2, 21].
8
Despite being widely accepted that the surfactant components responsible for lowering the surface tension are the phospholipids, namely DPPC, none of the phospholipid components is able to behave as a good surfactant by itself. Nowadays, there are sufficient experimental data that point to the requirement of the surfactant proteins, particularly the hydrophobic ones, for a suitable efficiency in surface tension reduction during expansion cycles of breathing [6].
Surfactant Proteins
8-10% of total surfactant mass is constituted by specific surfactant proteins (SPs) without which the surfactant mixture would not be fluid at body temperature
[16, 22]. Surfactant comprises two types of specific proteins: hydrophilic A and
SP-D, and hydrophobic SP-B and SP-C, named after their chronologic order of discovery
[12, 22].
SP-A and SP-D are large, multimeric, glycosylated proteins with several structural features in common, being members of the family of C-type (Ca2+ -dependent) lectins [22] – Figure 5. They are believed to be molecules involved in the immune response system due to their ability to recognize a broad spectrum of pathogens and bind alveolar macrophages. Recent work has related SP-A with structural organization of surfactant and SP-D with surfactant homeostasis [12].
9
Figure 5 – Trimeric carbohydrate recognition and neck domains of surfactant protein A (on the left) and head and neck domains of surfactant protein D (on the right). Adapted from [3].
The first description of the presence of hydrophobic surfactant proteins came from Phizackerley and co-workers in 1979 [24]. Unlike SP-A and SP-D, SP-B and SP-C do not have similar structural properties. SP-B is a disulphide-linked homodimer composed of two 79-residue polypeptide chains with a total net charge of approximately +6 at physiological pH, which interacts preferentially with superficial parts of the lipid layer and probably lacks transmembrane domains. SP-C is the most hydrophobic protein of the pulmonary surfactant material and the only one which is not produced by another type of body cells, lacking known homologous proteins. It consists of a transmembranar polypeptide chain with 33-35 residues (depending on the species and N-terminal truncation), with a net charge of approximately +3 at physiological pH and 1 or 2 (depending on the species) palmitoyl groups linked via thioester bonds to the cysteine residues [22, 24]. Together, SP-B and SP-C are important for [4]:
i. Lowering surface tension;
ii. The transport, attachment and incorporation of new surfactant material; iii. Rapid adsorption and stabilisation of the surfactant dynamic film.
10
Only the structure of SP-C was already fully determined and is depicted in
Figure 6.
Figure 6 –SP-C’s structure without the palmitoyl groups.
1.2.
S
URFACTANTP
ROTEINC:
B
IOCHEMISTRYince its discovery the pulmonary surfactant protein C (SP-C) has been extensively studied and although there is detailed structural information [26] about it, its functions in pulmonary surfactant are not completely understood yet [4, 12, 16, 17, 27-29].
1.2.1. Biosynthesis and sorting
Human SP-C is encoded by a single gene, located in the short arm of chromosome 8, that is transcribed and translated into a 21kDa proprotein – proSP-C. proSPC contains 197 amino acids, from which only amino acids 24-58 represent the mature SP-C [12]. This proprotein acts as a noncleavable signal sequence and membrane-anchoring domain towards the membrane of the endoplasmatic reticulum
11
(ER). Most of the experimental data is consistent with a type II orientation in the ER membrane, in which the N-terminus of the proprotein is located in the cytoplasm and the C-terminus in the lumen of the ER [17, 29].
During the transport of proSP-C through the secretory pathway, from type II pneumocyte cells to the lamellar bodies (LB), the proSP-C is palmitoylated on cysteine residues and cleaved – Figure 7. Four proteolytic steps occur that remove C- and N-terminal flanking domains from the mature SP-C: the C-flanking domain is removed in two cleavage steps in an acidic compartment distal to the Golgi and then the N-flanking domain is removed in two cleavage steps, the first one occurs in the multivesicular bodies (MVB) and the second one already in the LB [17] – Figure 7.
Figure 7 – Processing and sorting of pro-SP-C. On the left, the surfactant synthesis and on the right the proSP-C and its processing intermediates as well as the aminoacid sequence of the mature human SP-C. ER: endoplasmatic reticulum; MVB: multivesicular body; LB: lamellar body; TM: tubular myelin. Adapted from [17].
12
The release of LB in response to external stimuli [29] and their unpacking induces the formation of tubular myelin which supplies new SP-C to the interface monolayer or to the surfactant reservoir bilayers. When inserted into phospholipid membranes, SP-C disrupts lipid packing, promoting the movement of lipids between different membrane layers. Its helical structure and attached palmitoyl groups are the key factors for SP-C function, enhancing lipid spreading and film stability during the breathing cycle [28].
1.2.2. Structure
Isolated SP-C is composed of a mixture of isoforms with a major form taken as the biologically active molecule [27]. The human main form of SP-C is a dipalmitoylated polypeptide chain containing 35 amino acid residues with four different conserved regions concerning different species [26, 27, 30]:
i. The ‘poly-valine region’, from position 13 to position 28, which contains exclusively aliphatic residues with branched side-chains, mainly valine residues, that contributes largely for SP-C’s hydrophobicity;
ii. The C-terminal heptapeptide segment, after the poly-valine region (29-35); iii. Lysine-Arginine dibasic pair at positions 11 and 12;
iv. Palmitoylated cysteines at positions 5 and 6 (only absent in dog and mink) surrounded by prolines at positions 4 and 7.
The SP-C structure can be divided in three main parts: the intrinsically disordered N-terminal region (1-9), the helix region (9-34) and the C-terminal residue (35) with no assigned structure – Figure 8.
13
Figure 8 – Human SP-C sequence. Adapted from [29].
Palmitoylation
S-palmitoylated proteins can be broadly classified into 4 groups: the transmembrane proteins palmitoylated on cysteine residues located in close proximity to transmembrane domains (group 1), the proteins that are first myristoylated (group 2), the proteins that are first farnesylated (group 3) and the peripheral membrane proteins modified with palmitate only (group 4). The palmitoylation increases the hydrophobicity of proteins, contributing to their membrane association, and may play an important role in the subcellular protein trafficking or in the modulation of protein-protein interactions [31].
SP-C is a true lipoprotein, being part of group 1 S-palmitoylated proteins. Saturated palmitoyl moieties are linked as thioesters to adjacent cysteine residues, affecting the structure of SP-C and influencing its interactions with lung surfactant phospholipids [30, 32]. The palmitoylation occurs in the proSP-C state and takes place in a compartment distal to the ER, probably in the ER-Golgi intermediate compartment
14
Pig vs Human surfactant protein C
The structure of the acylated pig surfactant protein C was determined by two-dimensional H-NMR methods in a chloroform/methanol/0.1 HCl mixture (32:64:5, v:v:v) in 1994 by Jan Johansson and co-workers [26], being the only structure of SP-C available. They identified a regular α-helix motif comprising positions 9-34 with a length of 37 Å, which perfectly matches the thickness of a fluid DPPC bilayer, and a flexibly disordered domain comprising the N-terminal part, including the palmitoylcysteines. SP-C has a very non-polar surface formed by the poly-valine part of the helix (23 Å) whose side-chains contribute approximately 80% for the solvent accessible surface of the protein [27].
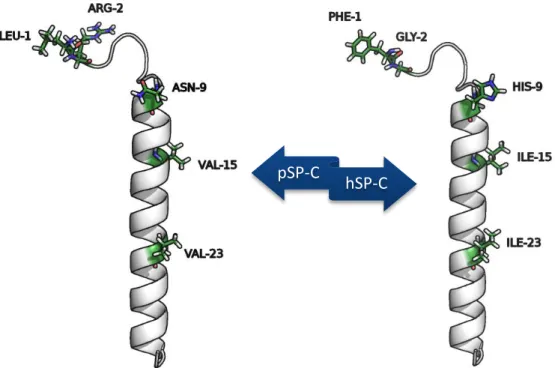
In terms of secondary structure motifs, both pig and human SP-C are expected to be similar despite their different aminoacid sequence. Their differences are illustrated in Figure 9 and consist of 5 mutations:
Figure 9 – Differences between the pig SP-C (pSP-C) and the human SP-C (hSP-C) in terms of aminoacid sequence.
The mutation in the N-terminal residue changes a Leucine in pSP-C for a Phenylalanine in hSP-C. This mutation does not introduce a significant structural modification as it switches between two nonpolar side-chain amino acids.
pSP-C
15
Although the mutation in the second position causes the loss of a positively charged residue (arginine) to a glycine, which can confer a greater flexibility to the N-terminal end, this positive charge is recovered in the mutation of the ninth residue that changes an asparagine for a histidine residue. This change of a positive charge from position 2 to position 9 is the most significant variation from pSP-C to hSP-C.
The remaining two mutations are equivalent, changing a Valine residue for an Isoleucine residue in the positions 20 and 23.
1.2.3. Transmembranar structure
Despite strongly supporting a transmembranar orientation in phospholipid bilayers the size of the SP-C α-helix in organic solvents (37 Å) does not agree with the thickness of phospholipid monolayers [27]. However, it was shown that a helix tilt angle of 24º relative to the normal in a lipid bilayer and of 70º in a lipid monolayer allows the maximum hydrophobic matching between the lipid acyl chains and the protein helical segment revealing that the helical portion of SP-C can act as a “hydrophobic lever” that anchors and spreads a maximum number of lipid molecules
16
Figure 10 – Different conformations of SP-C with the progressive compression of the surface film that leads to the formation of the surfactant bilayer reservoirs. Adapted from [7].
1.2.4. Functions
Beyond the functions that SP-C shares with SP-B (see section 1.1 Lung
Surfactant), SP-C is particularly important for the exclusion of surface material upon
compression and the reincorporation of the excluded material upon expansion, preventing the loss of surfactant material for the water subphase [19, 34, 35]. The surfactant protein C maintains the different layers of excluded material associated with each other and with the interface, promoting the stabilization of these multilayer structures. This seems to be possible due to its highly hydrophobic helix and its palmitoylated groups that maintain the protein attached with the membranar phospholipids within different lipid layers (including the interface monolayer) and due to its hydrophilic residues that have the propensity to interact with lipid headgroups of the same or the adjacent layers [4, 19, 35, 36] – Figure 11.
17
Figure 11 – Schematic representation of the fusion of the reservoir bilayer with the interface stabilized by SP-C during the pulmonary surfactant adsorption. Adapted from [4].
1.3.
PH-
DEPENDENT MISFOLDING OFSP-C
everal human diseases result from the failure of a specific peptide or protein to adopt or maintain its native conformation, undergoing conformational changes that lead to the conversion of its soluble native state into a misfolded form that, ultimately, progresses into highly stable and organized fibrillar aggregates, called amyloid fibrils. The greater stability of these aggregated forms causes the accumulation of fibrils in the organs and/or tissues of patients suffering from amyloid diseases [37].
Independently of the native structure of the peptide/protein, the final fibrillar aggregates have some features in common: a unique fibrillar macroscopic appearance (long non-ramified fibrils), binding affinity to thioflavin T and congo red dyes and a similar cross-β sheet structure where the β-sheet motifs are perpendicular to the fibril axis [37] – Figure 12. This conformation maximizes the interchain hydrogen-bonds that promote the protein aggregation.
18
Figure 12 – Three-dimensional (left) and two-dimensional (right) structural model of a fibrillar aggregate showing a cross-β pattern. Adapted from [38].
The formation of amyloid fibrils does not necessarily require a complete loss of protein structure before the aggregation phase or, in other words, the pathway for fibril formation can be distinct from that of unfolding. In the cases where the native state is thermodynamically more stable than the dominant partially unfolded state or when the native structure itself has a significant propensity to aggregate, the first step in the aggregation process is the formation of native-like or native oligomers, respectively, that retain the native topology and the original enzymatic activity. These intermediates undergo structural modifications to form amyloid protofibrils with extensive β-structure which are then enzymatically inactive [37].
Despite the initial postulate being that the accumulated material was the causative agent of various deposition disorders, more recent findings have raised the possibility that precursors of amyloid fibrils, such as the small oligomers and/or the protofibrils, are the real pathogenic species. The reason why prefibrillar aggregates could be toxic to cells, and hence appear to be the most likely causative agents of pathogenicity in deposition diseases, is now at the front line of research in this field
19
Different factors are known to strongly promote amyloid formation and some of them are related with the amino acid sequence of the polypeptide chain. A high content in hydrophobic residues, especially if they are solvent accessible, and a low content in charged ones, increase the aggregation propensity under normal physiological conditions. In addition, a high propensity to form β-sheet motifs and a low susceptibility to acquire α-helix conformation clearly contribute to the protein misfolding and aggregation propensities [37].
There are also some extrinsic factors related to amyloid susceptibility. The concentration of the protein promotes the protein aggregation and the pH as well as the ionic strength of the protein solution, can promote the misfolding and the aggregation process [37].
1.3.1. Pulmonary Alveolar Proteinosis
Among the amyloid diseases we can find the Pulmonary Alveolar Proteinosis (PAP), which was first recognized in 1958 when the existence and physical characteristics of the normal alveolar lining fluid had been recently established. Although there are three distinct types of PAP, congenital, secondary and acquired/idiopathic, they all result from the accumulation of surfactant material in the alveolar space with the associated disturbance of pulmonary gas exchange and minimal interstitial inflammation [39, 40].
Congenital cases occur in the neonatal period and are likely to be caused by mutations in the genes encoding for surfactant proteins B or C or in the gene that encodes for the receptor for granulocyte colony stimulating factor (GM-CSF), leading to severe respiratory distress [39-42].
Secondary PAP accounts for 5 to 10% of PAP cases and occurs in the adulthood due to lung infections, hematologic malignancies and after exposure to inhaled chemicals and minerals, being associated with the impairment or reduced number of alveolar macrophages [39-42].
20
More than 90% of all cases of PAP occur as a primary acquired disorder with no identifiable familial predisposition or secondary etiology in previously healthy adults with an average age of 39 years-old [40, 42].
Acquired PAP etiology... one step further?
The accumulated material in patients with PAP was first proposed to be functional surfactant in 1965 and three years later it was established that its content was chemically similar to that of lung surfactant material [42]. In vitro and in vivo experiments demonstrated that the accumulation of this material in the alveolar space was the result of a decreased surfactant clearance, rather than its increased synthesis, leading to the hypothesis that PAP resulted from a defect in macrophage function. Dysfunctions in GM-CSF were then proposed as the main cause of the clearance impairment [40, 42].
However, the decrease in the removal of surfactant material can be a cause as well as a consequence of surfactant accumulation and reports of GM-CSF therapy or lung transplantation have not been accepted as a reliable treatment of PAP. The best promising treatment seems to be the Whole Lung Lavage (WLL) in which the proteinaceous material is washed out from the alveoli and the effective oxygenation and ventilation is re-established [41, 43, 44].
The treatment for PAP was an important step to avoid its persistency, but the molecular mechanisms behind its origin would give us a way to prevent it. This is why the efforts to unveil the cause of PAP are still continuing to be done.
In 2001, Magnus Gustafsson and co-workers published an article in which they compared the brochoalveolar lavage (BAL) fluid of a PAP patient with those of six healthy controls. Although amyloid fibrils could be detected in both (PAP and healthy) cases, the amounts of aggregated material were only significant in the PAP case. Further analysis showed that significant amounts of aggregated surfactant protein C were specific of this disorder [45] and thus the clearance of alveolar SP-C fibrils is expected to be significantly impaired compared to that of native SP-C. In addition,
21
dimeric forms of SP-C, as well as their toxicity for alveolar macrophages and subsequent derangement of cellular metabolism, have been reported [46-50].
1.3.2. The stability of SP-C structure
The dimeric forms and the significant amounts of SP-C in the accumulated surfactant material of PAP patients are not, however, the only clues that point to the misfolding and further formation of amyloid fibrils of SP-C as the main cause of PAP.
Although the helical structure of this protein has been shown to be the thermodynamically most stable state in a membrane environment, studies of SP-C in solution have resulted in the observation of conformational modifications to an aggregated β-rich state and concomitant fibril formation [45, 51-58]. The α-helical monomeric state of SP-C seems to be a metastable intermediate which may undergo α→β transitions that ultimately result in amyloid fibril formation, depending upon the kinetics of particular pathways available and the milieu in which the protein resides
[56]. There are multiple factors that justify the misfolding of the protein and they are
as follows.
The α/β discordance
First of all, and without looking for anything else besides the polypeptide chain of surfactant protein C, the α-helix of the mature structure contains a very long continuous stretch of valine residues, which are well known to be underrepresented in helices and overrepresented in β-strands. Long stretches of α-helix/β-strand discordance in helical proteins can destabilize the α conformation, being strongly suggested to be a good indicator of the amyloid formation. The high β-strand propensity of these regions indicates that they are less likely to refold into a helical conformation than regions for which helices are predicted [52, 53].
Despite being stabilized within a membrane (apolar) environment, this strongly hydrophobic segment of SP-C is destabilized when in solution (more polar milieu) and, therefore, the removal from the membrane seems to contribute to SP-C amyloid formation.
22
Depalmitoylation
In spite of the presence of palmitoyl groups being essential for a spectrum of surface active behaviours relevant for lung surfactant function, such as anchoring SP-C in the bilayer or surface film and permitting maximal protein-phospholipid interactions
[32], isoforms of SP-C where one or both of the palmitoyl chains are missing have been
found (dSP-C) [27]. This may result from the labile nature of thioester-linked acyl chains, indicating that the protein undergoes cycles of depalmitoylation and repalmitoylation [31, 59].
However, these cycles can affect both incorporation into and interaction with the surrounding phospholipids. The depalmitoylation of SP-C was observed to affect the helix structure of the protein [60] by causing approximately 20% reduction of its α-helical content in a membrane environment at neutral pH [32, 62]. The removal of the palmitoyl moieties reduces the hydrophobic matching between the lipid acyl chains and the protein helical segment, and causes a possible loss of electrostatic interactions between the SP-C’s N-terminal positively charged residues and the phospholipid headgroups [32, 27, 62]. In addition, the loss of the acyl chains seems to decrease the kinetic stability of SP-C, since non- or mono-palmitoylated forms have a higher rate of α→β transition in organic solvent than the dipalmitoylated form [55].
All evidence points to a relation between the loss of the palmitoyl moieties, the misfolding of SP-C and further amyloid fibril formation. This hypothesis is supported by the identification of dSP-C in the aggregated material of a PAP patient’s BAL fluid [45]. For this reason, depalmitoylation seems to be another factor promoting C aggregates formation and can even be a contributing factor for the removal of SP-C from the bilayer or surface film upon compression (during expiration) [56].
Effect of pH
In 2003, Dluhy and co-workers [56] published an article in which they studied a pH-dependent mechanism for the formation of amyloid fibrils from dSP-C through the analysis of its transmission infrared spectra in organic solvent at different pH values, from 1.8 to 8.7. They observed that dSP-C at low pH adopted an α-helical secondary
23
structure similar to that of the fully acylated form at neutral pH. However, with the increase of pH, bands corresponding to amyloid motifs (twisted or coiled β-sheet strands or β-hairpins) and protein aggregation events were observed together with residual α-helix peaks, being the β-amyloid band the major contributor. A new decrease in pH showed the reversibility of this α→β transition with the re-emerge of the α-helix conformation with total recovery of the helical structure at the pH value of 1.8. This work identified the pH as one more contributive factor for the amyloid fibril formation from dSP-C. But is this factor physiologically relevant? Can the pH and SP-C acylation be somehow related?
To address the first question we should look at the proSP-C processing pathway in alveolar type II cells – Figure 7 in the section 1.2.1. As cited before, the proteolytic events that remove the N- and C-flanking domains of SP-C require an acidic environment [17], therefore an acidification in the secretory pathway is expected. In fact, during the first steps of the processing pathway, before any proteolytic event, proSP-C experiments a neutral pH that is progressively lowered to the value of approximately 5.5 in the lamellar bodies, where the final cleavage steps occur [63]. After that, and in the protein’s way towards the interface, there is a new increase in pH up to around 7, the value that was measured in the alveolar subphase [64].
As for the second question, the acidification of the milieu has already been shown to affect the acylation equilibrium of SP-C [30, 65]. The treatment of surfactant protein C with basic agents is known to promote the protein deacylation [30]. Actually, at a pH value equal or greater than 8 the loss of palmitic groups is more likely to occur and at neutral pH a partial deacylation is observed [65].
Taking all these observations into account, we can identify the pH as another possible contributing factor for the amyloid fibrils formation from SP-C and even expect its correlation with the protein depalmitoylation. Since the loss of the palmitoyl moieties promotes the protein exclusion from the surface membrane, the pH can be indirectly related with the protein exposure to the subphase aqueous environment.
24
1.4. S
TUDYING THEM
ISFOLDING OFSP-C
BYM
OLECULARM
ODELLINGM
ETHODSP-C is one of the most non-polar naturally occurring polypeptides known and its pronounced hydrophobicity makes it difficult to handle experimentally since the protein is unstable under certain conditions. The structure of SP-C is especially sensitive to its environment, depending on protein-solvent interactions and on the rearrangement of the solvent molecules around it [66]. These features make computer simulation methods a perfect candidate to study SP-C.
The conformational behaviour of the deacylated surfactant protein C in different solvent conditions (chloroform, methanol or water) was already studied using computational approaches by several groups [67-70], but a proper organic solvent mixture resembling the one used in the structure determination (and other experimental studies) was never used. Besides, the simulation time in those theoretical studies was always too small (<10 ns) to account for a proper analysis of the conformational behaviour of the protein, and the dependence of the helix stability and formation of β-sheet motifs on pH was never addressed.
Accounting for the destabilizing effects of pH and depalmitoylation on the stability of SP-C, we propose to study the conformational stability of the deacylated protein in a chloroform/methanol/water mixture at different pH values in the range 3 to 8, using state-of-the-art molecular modelling methods. We are particularly interested in studying the atomic-level aspects of the misfolding process of the dSP-C observed experimentally and in the characterization of misfolded beta-rich species that may be intermediate states in the amyloid fibril formation process.
25
2. Molecular Simulation
umerical simulations began to play a leading role in weapons design during the Second World War [71] and since then they have been developed and their application has extended to several fields. In 1952, Metropolis, the Rosenbluths and the Tellers performed the first computer simulations of fluids [72] and since then the development of statistical mechanics, of new algorithms and new ways of modelling in computational biochemistry and molecular biology, have dictated the increasing popularity of these theoretical methods. Of course that none of this would have been possible without the progress in the computational field given that computer constraints were then, and are still now, the principal limiting factor in computer simulation [71]. Computers have become faster, cheaper and more powerful.
There are several levels of modelling that focus on different aspects of molecular motion and are useful to study different biological processes – Table 1.
Table 1 – Examples of levels of modelling [73].
Methods Degrees of freedom Timescale Quantum Dynamics atoms, nuclei, electrons picoseconds
Quantum Mechanics atoms, nuclei, electrons no time scale
Classical Statistical Mechanics
(MD, MC, force fields)
atoms, solvent nanoseconds microseconds
Statistical Methods
(database analysis)
groups of atoms, aminoacid residues, bases
no time scale
Continuum Methods
(hydrodynamics and electrostatics)
electrical continuum, velocity continuum, etc.
supramolecular
Kinetic Equations populations of species macroscopic Currently, computer simulation methods are widely used to reproduce or predict the molecular behaviour of biological systems in order to understand the interactions/events underlying at atomic level. One of the major applications is the
26
study of the dynamic nature of biological molecules and the large structural changes that they can undergo specially if submitted to different environmental conditions. Some examples of the computational approaches in use include: determination of macromolecular structure based on either nuclear magnetic resonance (NMR) or X-ray data, prediction of 3D structures based on primary sequence when the experimentally determined structure is not available and drug development, design or refinement. Clearly, computational and theoretical studies of biological molecules have advanced significantly in recent years and much thanks to the ever-increasing number of available structures and significant methodological improvements. The combination of these advances with the exponential increase in computational resources has greatly expanded, and will continue to expand, the applicability of computational approaches to the study of biomolecules [74].
In this chapter we will talk about the laws of statistical mechanics that enable us to relate the microscopic computational data with macroscopic experimental results, and about some molecular simulation methods like Molecular Mechanics/Molecular Dynamics, Continuum Electrostatics/Monte Carlo and Constant-pH Molecular Dynamics methods.
2.1. S
TATISTICALM
ECHANICS–
F
ROM MICROSCOPIC DATA TO MACROSCOPIC PROPERTIESrom a macroscopic point of view we focus on thermodynamic properties (like temperature, heat capacity, pressure) that are usually measured by typical experiments as average properties, averaged over a large number of particles and, usually, also averaged over the time of the measurement [72, 76]. On the other hand, at a microscopic level we focus on the behaviour of individual atoms, which involves abstract ideas and tends to be based on theory rather than experiment, since no real experiment provides us with such detailed information [75, 74].
27
Computer simulation techniques enable us to get atomic level insights into a particular system using a microscopic approach. However, rather than being interested in the individual properties of the constituent atoms and molecules, we are interested in the average performance of our simulated system [76]. In addition, we will eventually want/need to compare our computational (microscopic) data with experimental (macroscopic) studies. This requires a link between the two approaches.
The connection between the microscopic behaviour and macroscopic properties of molecular systems is governed by the laws of statistical mechanics which allow us to express thermodynamic properties in terms of microscopic quantities [71].
2.1.1. The Ergodic Hypothesis
For any statistical problem it is easy to understand that the greater the number of measurements performed, the closer the calculated sample mean would be to the true mean, and the smaller the sample deviation would become. Transported to the computer simulations, this means that if one generates several copies of the system of interest where some thermodynamic properties are kept constant, the calculated average over this large number of replicates would follow the same principle – Figure
13. Such an average is called an ensemble average and, according to the ergodic
hypothesis, the averaged value over this ensemble is exactly the same as the time average that would be calculated by studying the time evolution of the original system: both kinds of measurement should produce the same result [75-77].
28
Figure 13 – Example of a canonical ensemble scheme, where the number of particles (N), the volume (V) and the temperature (T) are kept constant [76].
Molecular simulation methods allow us to generate a statistically representative ensemble of configurations of the system in order to obtain good ensemble averages. Following the natural evolution of the system makes it possible to predict the static and dynamic properties directly from the underlying interactions between the molecules [74].
There are four major types of ensemble in the theory of statistical thermodynamics, and they are classified according to what thermodynamic properties are kept constant in all the replicas – Table 2.
Table 2 – Types of thermodynamic ensemble where N stands for number of particles, V for volume, T for temperature, E for total energy, p for pressure and μ for chemical potential [76].
Type of ensemble Canonical Microcanonical Isothermal- -Isobaric Grand canonical Constant thermodynamic properties N, V, T N, V, E N, p, T μ, V, T