ESTUDO CARIOTÍPICO EM DUAS
ESPÉCIES DE CUCULIFORMES AMERICANOS (Aves) (*)
Aldo W a l d r i g u e s ( " ) Iris Ferrari (***)
Antonio Faustino Neto (****)
Resumo
Foram estudadas citogeneticamente duas espécies de aves da Família Cucu-lidae: Piaya cayana (Phaenicophaeinae) e Crotophaga major (Crotophoginae)
captu-radas na região de Manaus (AM). Os cromossomos foram preparados a partir de células da medula óssea, em cultura de curta duração [4 horas). A análise ca-riotípica foi realizada por métodos con-vencionais, obtendo-se dados numéricos, morfológicos e métricos dos cromosso-mos destas duas espécies. A classifi-cação dos cromossomos obedeceu à pro-posição de Levan et al. (1964). Os cro-mossomos foram divididos em três gru-pos: dois de macrocromossomos (grupos A e B) e um de microcromossomos (gru-po C). Foram considerados como macro-cromossomos, todos aqueles que apre-sentaram Proporção Relativa maior ou igual a 4%. Piaya cayana apresentou 76
cromossomos como número diplóide, dos quais 22 foram considerados como ma-crocromossomos. O cromossomo Z ó
submetacêntrico e o segundo do carióti-po, enquanto o W é subtelocêntrico e o décimo primeiro em tamanho. Crotopha-ga major apresentou 64 cromossomos,
dos quais, 22 foram considerados como macrocromossomos. O cromossomo Z é metacêntrico e o quarto do cariótipo, en-quanto o cromossomo W é submetacên-trico e o sexto em tamanho.
INTRODUÇÃO
São poucos os dados citogenéti -cos d i s p o n í v e i s para a classe A v e s . A escassez destes dados
não se v e r i f i c a s o m e n t e nas espé-cies brasileiras, mas s i m , em to-das as espécies do m u n d o . Para se v e r i f i c a r a extensão da inexis-t ê n c i a de dados cariológicos so-bre aves, basta observar os forne-cidos por Takagi & Sasaki (1974): menos de 2 % de aves f o r a m ca-riotipadas, e mais de 5 0 % de suas ordens não apresentavam quais-quer estudos c i t o g e n é t i c o s .
O Brasil, pela sua vasta exten-são t e r r i t o r i a l , pela grande cober-t u r a f l o r e s cober-t a l e pelos climas cober- tropi-cal e s u b - t r o p i c a l , é o país que apresenta maior diversidade em espécies de a v e s . A p e s a r disso m u i t o pouco se conhece citogene-t i c a m e n citogene-t e sobre as m e s m a s , prin-c i p a l m e n t e pelas t é prin-c n i prin-c a s atuais de f o r m a ç ã o de bandas cromossô-m i c a s .
(*) Este estudo foi parcialmente financiado pela Universidade Estadual de Londri-na (PICD-CAPES) e pelo CNPq (Proc. 40.2697/79; PIG III) e Instituto NacioLondri-nal de Pesquisas da Amazônia (INPA).
(**) Fundação Universidade Estadual de Londrina — UEL (***) Faculdade de Medicina de Ribeirão Preto — USP
O estudo c o m p a r a t i v o do carió-tipo de espécies relacionadas po-de c o n t r i b u i r para um m e l h o r co-nhecimento dos m e c a n i s m o s de evolução c r o m o s s ô m i c a , b e m co-mo, das relações f i l o g e n é t i c a s en-tre os grupos a t u a i s . Em aves, os
poucos e s t u d o s c i t o t a x o n ô m i c o s até agora realizados, t ê m demons-trado a presença de certos aspec-tos e m c o m u m na morfologia dos
c r o m o s s o m o s . A maior ou m e n o r semelhança c a r i o t í p i c a das espé-cies relacionadas, parece ser de-pendente da o r d e m a que estas aves p e r t e n c e m , sugerindo a
ocor-rência c o m maior freqüência de rearranjos e s t r u t u r a i s nos cromos-somos de d e t e r m i n a d a s ordens.
Os dados a n a t ô m i c o s , bioquími-cos e c o m p o r t a m e n t a i s disponí-veis sobre os C u c u l i f o r m e s , são insuficientes para e l u c i d a r e m seu relacionamento f i l o g e n é t i c o c o m aves que a p r e s e n t a m aspectos se-m e l h a n t e s .
Estudos c i t o g e n é t i c o s , que m u i -to poderiam c o n t r i b u i r para uma melhor visão evolutiva e taxonó-mica da O r d e m C u c u l i f o r m e s , só f o r a m realizados e m quatro de suas e s p é c i e s : C u c u l u s
cano-rus (Yamashina, 1946), E u d y n a m y s s c o l o p a c e a s c o l o p a c e a
(Ray-Chau-dhuri, 1967), G ü i r a g u i r a (Lucca,
1974; W a l d r i g u e s & Ferrari, 1980) e C r o t o p h a g a a n i (Waldrigues &
Ferrari, 1982). Este trabalho apresenta o estudo cariotípico do A n u -guaçu ( C r o t o p h a g a m a j o r ,
Cro-tophaginae) e do Alma-de-gato
(Pyaya c a y a n a , Phaenicophaeinae)
ambos p e r t e n c e n t e s à Família C u -c u l i d a e .
MATERIAL E MÉTODOS
Foram estudados c i t o g e n e t i c a -m e n t e ani-mais adultos das espé-cies C r o t o p h a g a m a j o r ( G m e l i n ,
1978) (5 machos e 4 fêmeas), e
Piaya c a y a n a (Linnaeus, 1766) (4
machos e 3 f ê m e a s ) (Cuculifor-m e s , A v e s ) , capturados na região de Manaus ( A M ) .
A m e d u l a óssea dos o s s o s lon-gos dos m e m b r o s i n f e r i o r e s f o i re-tirada i m e d i a t a m e n t e após as aves t e r e m sido a b a t i d a s . A m e d u l a destes o s s o s f o i então colhida e ressuspensa e m 10 ml de m e i o de cultura preparado a p a r t i r do
" P o w d e r e d Tissue Culture M é d i u m RPMI 1640" (GIBCO, USA) prepa-rado para c u l t u r a de l i n f ó c i t o s de sangue p e r i f é r i c o .
A p ó s quatro horas de incubação a 41°C, f o r a m colocadas duas go-tas ( a p r o x i m a d a m e n t e 0,10 m l ) de solução de c o l c h i c i n a (0,016 m g / m l ) e homogeneizou-se, dei-xando-se o m a t e r i a l e m incubação por mais duas h o r a s . C o m p l e t a d o o t e m p o de incubação, após agita-ção dos f r a s c o s de c u l t u r a , trans-feriu-se o c o n t e ú d o para t u b o s de c e n t r í f u g a . Centrifugou-se a 800 r . p . m . d u r a n t e 10 m i n u t o s e, eli-minou-se o s o b r e n a d a n t e .
A s c é l u l a s f o r a m ressuspensas em c e r c a de 10 ml de solução hi-potônica (KCI a 0.075M) aquecida a 41°C. O material p e r m a n e c e u 5 m i n u t o s nesta solução, na e s t u f a a 41°C, e, e m seguida, f o i c e n t r i f u -gado por 5 m i n u t o s , a 800 r . p . m .
Eliminou-se a solução hipotôni-ca e procedeu-se a f i x a ç ã o , por três vezes c o n s e c u t i v a s , juntando-se 5 ml de solução f i x a d o r a prepa-rada r e c e n t e m e n t e (metanol — ácido acético na proporção de 3 : 1 ) .
Lâminas p e r f e i t a m e n t e limpas f o r a m deixadas e m u m r e c i p i e n t e c o m água gelada. U m a gota do m a t e r i a l fixado era colocada sobre a p e l í c u l a de água que recobria a l â m i n a . Retirou-se o excesso de água c o m papel de f i l t r o e secou-se u m a parte das lâminas na cha-ma de lâmpada a álcool e a o u t r a parte f o i secada à t e m p e r a t u r a am-b i e n t e . Duas lâminas (das seca-das na c h a m a de álcool) f o r a m co-radas e m solução de orceína,
pre-v i a m e n t e f i l t r a d a (1 g de orceína e m 100 ml de ácido acético glacial 6 0 % ) , durante 3 m i n u t o s . Em se-guida, desidratou-se o material através de banhos sucessivos e m álcool 1 0 0 % , álcool-xilol e x i l o l , e procedeu-se a m o n t a g e m e m Bál-s a m o da C a n a d á .
De cada espécie f o r a m analisa-das m i c r o s c o p i c a m e n t e , no mini- • mo, 50 m e t á f a s e s de diferentes a n i m a i s . C o m a objetiva de imer-são, as m e t á f a s e s f o r a m desenha-das e s q u e m a t i c a m e n t e e m papel s u l f i t e . Esta análise forneceu o mais provável n ú m e r o diplóide de c r o m o s s o m o s de cada espécie a n a l i s a d a .
Uma vez encerrada a análise nu-m é r i c a , as nu-m e l h o r e s nu-metáfases fo-ram selecionadas e fotografadas em F o t o m i c r o s c ó p i o Zeiss c o m ocular 12.5X, o b j e t i v a 100X e opto-var 1.25X. Foi utilizado f i l m e Pa-natomic-X da Kodak. Estas foto-grafias, após copiadas e m papel (Kodabromide F-3), f o r a m recorta-das e os pares de c r o m o s s o m o s utilizados para a m o n t a g e m dos r e s p e c t i v o s c a r i ó t i p o s .
Os c r o m o s s o m o s f o r a m subdivi-didos e m dois g r u p o s : macrocro-m o s s o macrocro-m o s , considerados aqueles c o m PR (proporção relativa) maior ou igual a 4 % , e m i c r o c r o m o s s o -m o s , aqueles c o -m PR inferior a 4 % . Os m a c r o c r o m o s s o m o s fo-ram d i v i d i d o s , por sua vez tam-b é m e m dois g r u p o s : A e B, de
braços ( B ) . Todos os m i c r o c r o -mossomos f o r a m agrupados n u m m e s m o g r u p o : o grupo C.
Foram m e d i d o s os m a c r o c r o mossomos s o m á t i c o s d e , no m í n i -mo, 25 f i g u r a s m e t a f á s i c a s prove-nientes de d i f e r e n t e s e s p é c i m e n s de cada e s p é c i e . A t é c n i c a e m -pregada para estas m e d i ç õ e s foi a seguinte:
A . as m e t á f a s e s selecionadas para o p r o c e s s a m e n t o m é t r i c o deveriam a p r e s e n t a r os c r o m o s s o -mos p e r f e i t a m e n t e c o r a d o s , c o m seus contornos n í t i d o s e c o m as cromátides b e m s e p a r a d a s .
B. vinte e cinco m e t á f a s e s de diversos e x e m p l a r e s de cada espécie f o r a m e s c o l h i d a s e f o t o m i -crografadas e m m i c r o s c ó p i o Z e i s s , pelo processo já d e s c r i t o anterior-m e n t e .
C . o f i l m e f o t o g r á f i c o , após re-velação, f o i m o n t a d o c o m o se fos-se u m d i a p o s i t i v o .
D. os " d i a p o s i t i v o s " f o r a m pro-jetados sobre f o l h a s de papel "Flor-post", por m e i o de u m pro-jetor CABIN 2000-R. O a u m e n t o fornecido por este aparelho foi avaliado através da utilização de uma escala e m u m dos " d i a p o s i t i -v o s " (padrão). Este era projetado antes de cada m e t á f a s e s e r dese-nhada, mantendo c o m i s t o o aumento c o n s t a n t e e n t r e as m e t á f a -s e -s .
E. a projeção f o i f e i t a sobre uma parede f i x a e lisa, onde era
afixada, por m e i o de f i t a a d e s i v a , a f o l h a de papel " F l o r - p o s t " .
F. os c o n t o r n o s dos m a c r o c r o -m o s s o -m o s f o r a -m d e s e n h a d o s so-bre a f o l h a de papel c o m lápis de ponta f i n a .
G . s o b r e o d e s e n h o , f o r a m marcados o s eixos dos m a c r o c r o m o s -s o m o -s (linha que d i v i d i a longitudi-nalmente cada c r o m á t i d e e m par-t e s iguais) c o m lápis e s o b r e e s par-t e passou-se u m a c a r r e t i l h a (Skandia t o o l s ) cujo passo c o r r e s p o n d e a u m a unidade m é t r i c a convencional . Para e f e t u a r e s t a fase do t r a -balho colocou-se, s o b a f o l h a de papel " F l o r - p o s t " , u m a f o l h a de papel carbono c o m f a c e ativa v o l t a -da para as c o s t a s do papel que continha o c o n t o r n o dos macrocro-m o s s o macrocro-m o s , j á c o macrocro-m seus eixos es-t a b e l e c i d o s . Em s e g u i d a , a
carre-tilha era passada c u i d a d o s a m e n t e sobre os e i x o s .
I. os m i c r o c r o m o s s o m o s f o r a m eliminados desta análise, por não se poder d e l i m i t a r n i t i d a m e n t e seus c o n t o r n o s e f o r m a s .
J . f o r a m calculados os tama-nhos médios dos lotes diplóides e haplóides através da média arit-m é t i c a dos valores obtidos dos m a c r o c r o m o s s o m o s das metáfa-ses a n a l i s a d a s .
K. f o r a m calculadas as propor-ções relativas (PR), proporcionais ao tamanho relativo (TR), de cada m a c r o c r o m o s s o m o de acordo c o m a seguinte f ó r m u l a :
C
( P i + * » )PR =
j = n
C (Pi + <•)
i = I
onde, PR = proporção relativa;
V. (PT + q,) = Somatória dos
pon-tos do iésimo m a c r o c r o m o s s o m o na e n é s i m a m e t á f a s e .
j = n
7~! ((Pi + qi) = Somatória dos
i == I pontos de to-dos os macro-c r o m o s s o m o s
na e n é s i m a m e t á f a s e .
Obs.: Preferimos chamar cie PR e não TR devido à não inclusão dos mi-crocromossomos nesta análise. L. para a d e t e r m i n a ç ã o das po-sições dos c e n t r ô m e r o s dos ma-c r o ma-c r o m o s s o m o s de ma-cada uma das espécies analisadas, bem c o m o , para as suas c l a s s i f i c a ç õ e s , f o i utilizada a f ó r m u l a indicada por Levan e t a l . 1964).
M . a razão de braços dos cro-m o s s o cro-m o s foi obtida através da seguinte f ó r m u l a :
C
P IB =
C P ,
onde, B = Razão ou Relação de braços dos macrocro-m o s s o macrocro-m o s .
T""? t\* = Somatória dos pontos
-do braço longo -do ié-s i m o macrocromoié-sié-so- macrocromosso-mo na enésima metá-f a s e .
YZ Pi — Somatória dos pontos
do braço curto do ié-s i m o macrocromoié-sié-so- macrocromosso-mo na enésima metá-f a s e .
RESULTADOS
ALMA-DE-GATO (Piaya cayana)
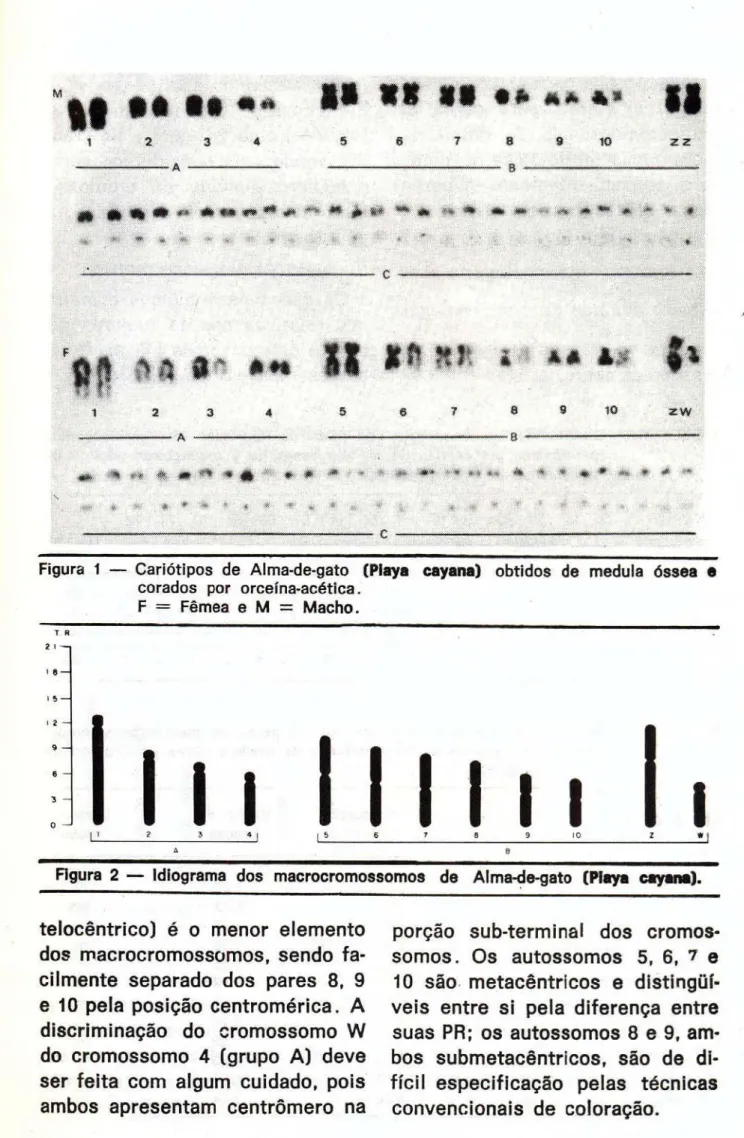
N ú m e r o de c r o m o s s o m o s
Em 77 m e t á f a s e s , analisadas mi-c r o s mi-c o p i mi-c a m e n t e , enmi-controu-se 27
(35,06%) c o m número diplóide de c r o m o s s o m o s igual a 76 (Tab. I), sendo este o mais provável núme-ro diplóide de c r o m o s s o m o s des-ta espécie ( F i g . 1 ) .
Análise m o r f o l ó g i c a e m é t r i c a
Os dados m o r f o l ó g i c o s e métri-cos relativos aos 11 pares de ma-c r o ma-c r o m o s s o m o s s o m á t i ma-c o s obti-dos pela análise de t r i n t a e uma metáfases de m e d u l a óssea de
Piaya cayana são mostrados na
Os c r o m o s s o m o s desta e s p é c i e foram divididos em t r ê s g r u p o s : A, B e C. Os dois p r i m e i r o s grupos correspondem aos m a c r o c r o -mossomos, enquanto no grupo C estão todos os m i c r o c r o m o s s o -m o s . O grupo A é f o r m a d o por
quatro pares de c r o m o s s o m o s , to-dos com c e n t r ô m e r o na região terminal ( t ) . O c r o m o s s o m o 1 é o maior de todo o c o m p l e m e n t o , en-quanto o 2, 3 e 4 c o r r e s p o n d e m ao 5.°, 7." e 10.° e m tamanho,
respec-t i v a m e n respec-t e . A d i s c r i m i n a ç ã o d e s respec-t e s e l e m e n t o s é realizável pela dife-rença entre suas PR.
O grupo B é c o m p o s t o por sete
pares de c r o m o s s o m o s (seis au-t o s s ô m i c o s e o par Z W ) . O cro-m o s s o cro-m o Z ( s u b cro-m e t a c ê n t r i c o ) é o maior do grupo e o segundo maior elemento do cariótipo, sendo dis-tingüível dos demais t a n t o pela sua PR quanto pela posição cen-t r o m é r i c a . O c r o m o s s o m o W
(sub-TABELA I — Distribuição da freqüência dos números diplóides de cromossomos me-tafásicos em células de medula óssea de 7 exemplares de Piaya cayana.
NÚMERO 48 61 63 65 68 69 70 72 73 74 75 76 77 78 79 80 82 83 84
TOTAL
SEXO
MACHO 1 1 1 1 1 1 — 2 4 15 — 6 4 3 2 2 1 45 FÊMEA _ _ 1 1 2 1 1 1 5 12 1 2 2 — — 2 1 3 2
TOTAL 1 1 1 1 1 1 3 2 1 3 9 27 1 8 6 3 2 4 2 77
TABELA II — Dados morfológicos e métricos dos 11 pares de macromossomos
ob-- tidos pela análise de 31 figuras i metafásicas de medula óssea de
Piaya cayana.
Cromossomo Grupo Grupo Proporção Razão de
Classifi-Relativa braços cação
1 12,80 11,49 t
2 A 8,90 8,23 t
3 7,56 7,80 t
4 5,98 7,63 t
Z 12,11 1,75 sm
5 10,84 1,27 m
6 9,43 1,20 m
7 B 8,51 1,19 m
8 6,73 2,65 sm
9 6,30 1,72 sm
10 5,68 1,34 m
M
8 9
B
10 z z
10 z w
1 *
Figura 1 — Cariótipos de Alma-de-gato (Playa cayana) obtidos de medula óssea e
corados por orceína-acética. F = Fêmea e M = Macho.
T R ? I — 1
I 8 —
i 5 —
i í
-» —
e
3
-o -
M i
M i n
i
Figura 2 — Idiograma dos macrocromossomos de Alma-de-gato (Playa cayana).
t e l o c ê n t r i c o ) é o m e n o r e l e m e n t o dos m a c r o c r o m o s s o m o s , s e n d o f a -c i l m e n t e s e p a r a d o dos p a r e s 8, 9 e 10 pela p o s i ç ã o c e n t r o m é r i c a . A d i s c r i m i n a ç ã o d o c r o m o s s o m o W do c r o m o s s o m o 4 ( g r u p o A ) d e v e ser f e i t a c o m a l g u m c u i d a d o , p o i s a m b o s a p r e s e n t a m c e n t r ô m e r o na
p o r ç ã o s u b t e r m i n a l d o s c r o m o s -s o m o -s . O -s a u t o -s -s o m o -s 5, 6, ' e
10 são m e t a c ê n t r i c o s e d i s t i n g ü í -v e i s e n t r e s i p e l a d i f e r e n ç a e n t r e
suas PR; o s a u t o s s o m o s 8 e 9, amb o s s u amb m e t a c ê n t r i c o s , são de d i -f í c i l e s p e c i -f i c a ç ã o p e l a s t é c n i c a s
O grupo C é c o m p o s t o
provavel-mente por vinte e sete pares de m i c r o c r o m o s s o m o s de d i f í c i l ca-racterização m o r f o l ó g i c a e métri-ca e, c o n s e q ü e n t e m e n t e , o parea-mento destes c r o m o s s o m o s é im-praticável (Fig. 1 ) .
ANU-GUAÇU ( C r o t o p h a g a m a j o r )
Número diplóide de c r o m o s s o m o s
Em 88 metáfases analisadas m i -croscopicamente, provenientes de
TABELA III — Distribuição da freqüência dos números diplóides de cromossomos metafásicos em células de medula óssea de 9 exemplares adultos de
Crotophaga major (Anu-guaçu).
NÚMERO 56 57 59 60 61 62 64 65 66 67 68 69 70 71 72
TOTAL SEXO
MACHO 1 1 — 2 3 4 20 3 4 3 2 1 1 1 1 47 FÊMEA 2 1 1 2 2 3 17 4 3 2 2 1 — 1 — 41
TOTAL 3 2 1 4 5 7 37 7 7 5 4 2 1 2 1 88
TABELA IV — Dados morfológicos e métricos dos 11 pares de macrocromossomos, obtidos pela análise de 27 metáfases de medula óssea de Crotophaga
major (Anu-guaçu).
Cromossomo Grupo Grupo Proporção Razão de
Classifi-Relativa braços cação
1 19,95 1,05 m
2 A 15.65 1.73 sm
3 11,94 ¡.07 T l
7 9.55 1.08 m
4 6.92 1.15 m
5 6,37 1.12 m
W 6.31 2,04 sm
6 5.52 1.11 m
7 B 4.80 1.09 m
8 4,79 1.23 m
9 4,17 1.74 sm
10 4,00 1,24 m
nove indivíduos adultos (machos e f ê m e a s ) , encontraram-se 37 (42.04%) c o m 2n igual a 64 (Tab. III), sendo este considerado c o m o o número diplóide de c r o m o s s o -mos mais provável para esta es-p é c i e .
cromosso-tl II
I »
1 2 3
— • A
• 1 99 • • M • • • •
4 5 6 7 8 9 10
II
8
if
1 1 1 1 M U 4 < 1 1 12 3 4 5 6 7 8 9 10 z w
1 I ê m * * * • - « »
Figura 3 Cariótipos de Anu-guaçu (Crotophaga major) obtidos de medula óssea e corados por orceína-acétlca.
F = Fêmea e M = Macho.
I R
I I I
I t I I
I I
Figura 4 — Idlograma dos macrocromossomos do Anu-guaçu (Crotophaga major).
mos sexuais) são m o s t r a d o s na Tab. IV. Os m a c r o c r o m o s s o m o s desta espécie f o r a m s u b d i v i d i d o s e m dois grupos (A e B), enquanto
todos os m i c r o c r o m o s s o m o s fo-ram colocados n u m único g r u p o : o grupo C (Fig. 3).
O grupo A é c o m p o s t o por três
pares de m a c r o c r o m o s s o m o s : dois m e t a c ê n t r i c o s ( e l e m e n t o s 1 e 3)
e u m s u b m e t a c ê n t r i c o (cromosso-mo 2 ) . A d i s c r i m i n a ç ã o dos ma-c r o ma-c r o m o s s o m o s 1 e 3 do grupo
reali-zada pelas diferenças nas PR; en-quanto que pelas posições centro-méricas se pode diferenciar o cro-mossomo 2 dos c r o m o s s o m o s 1 e 3.
O grupo B é c o m p o s t o por oito
pares de m a c r o c r o m o s s o m o s : se-t e pares de a u se-t o s s o m o s e u m par de s e x u a i s . A separação do Z, que é metacêntrico e o maior grupo B, do c r o m o s s o m o 3 (grupo A ) , tam-bém m e t a c ê n t r i c o . pode s e r reali-zada pela diferença nas suas PR. Por este m e s m o tipo de compara-ção se pode diferenciar o Z dos demais elementos do grupo B.
A d i s c r i m i n a ç ã o do cromosso-mo W dos a u t o s s o m o s 6, 7, 8 e 10 pode ser realizada pela posição c e n t r o m é r i c a , já que o W é sub-metacêntrico e e s t e s a u t o s s o m o s são m e t a c ê n t r i c o s . enquanto pela diferença entre suas PR pode dis-tinguir-se o W do a u t o s s o m o 9 ( s u b m e t a c ê n t r i c o ) .
A especificação dos autosso-mos 4, 5 e 6 (todos m e t a c ê n t r i c o s ) deve ser realizada c o m algum cui-dado, pois a d i f e r e n ç a entre suas PR é pequena; o m e s m o acontece em relação aos pares 7, 8 e 10. O idiograma dos m a c r o c r o m o s s o m o s desta espécie está representado
na F i g . 4 .
O grupo C é provavelmente
for-mado por vinte e um pares de mi-c r o mi-c r o m o s s o m o s , sendo os dois primeiros pares claramente telo-cêntricos (Fig. 3 ) . Os demais, em
conseqüência de suas d i m i n u t a s
d i m e n s õ e s , são de d i f í c i l
caracte-rização m é t r i c a e m o r f o l ó g i c a .
DISCUSSÃO
Os dados c a r i o t í p i c o s o b t i d o s
de P. cayana e de C . m a j o r
most r a m grandes diferenças n u m é r i -cas, m é t r i c a s e m o r f o l ó g i c a s en-t r e s i .
Em cerca de 8 0 % das e s p é c i e s de aves já cariotipadas, o n ú m e r o diplóide de c r o m o s s o m o s se s i -t u o u en-tre 76 e 84 c r o m o s s o m o s (Bloom, 1969; Ray-Chaudhuri et a l .f 1969; Ray-Chaudhuri, 1973;
Srb, 1974). Segundo Srb (1974) de
38 espécies da Família A n a t i d a e ( A n s e r i f o r m e s ) já cariotipaJas, 27 ( 7 1 % ) apresentaram 2n igual a 80. enquanto que os números diplói-des mais p r ó x i m o s (78 e 82)-foram observados e m menos de 1 0 % das demais espécies desta famí-l i a . Esta m e s m a homogeneidade numérica foi encontrada em Pha-sianidae ( G a l l i f o r m e s ) por H e r m a n & Holynski (1970).
P. cayana apresenta número
di-plóide de c r o m o s s o m o s provavel-m e n t e igual a 76, p o i s , e provavel-m 77 provavel- me-táfases analisadas, f o r a m encon-tradas 27 (35,06%) c o m este nú-m e r o . Se f o r e nú-m levadas e nú-m consi-deração contagens e m torno des-t e valor (2 unidades para mais o u
mais provável foi 64, pois e m 88 metáfases analisadas, 37 (42.04%) delas m o s t r a r a m tal c o n t a g e m ; que sobe para 6 5 % ao i n c l u i r e m -se os valores de 2n adjacentes a este v a l o r . Portanto, apesar de es-tas duas espécies p e r t e n c e r e m à
mesma f a m í l i a (Cuculidae), p o r é m a d i f e r e n t e s s u b f a m í l i a s (Phaeni-cophaeinae e Crotophaginae), elas m o s t r a m números diplóides de c r o m o s s o m o s bastante d i f e r e n t e s , 76 e 64, r e s p e c t i v a m e n t e .
Os demais C u c u l i f o r m e s já ca-riotipados m o s t r a m n ú m e r o di-plóide de c r o m o s s o m o s inferior a 78: Cuculus canorus (Yamashina,
1946) t e m 2n = 72; Guira g u i r a , mostrou 72 c r o m o s s o m o s na aná-lise de Lucca (1974) e 66 na análi-se de W a l d r i g u e s & Ferrari (1980);
Crotophagaani (Waldrigues & Fer-rari, 1982) t e m 70 c r o m o s s o m o s . O maior n ú m e r o diplóide de cro-m o s s o cro-m o s f o i encontrado e cro-m Eu-dynamys scolopacea scolopacea (Ray-Chaudhuri, 1967) c o m 2n igual a 78.
A exata determinação do núme-ro diplóide de c r o m o s s o m o s e m aves é dificultada pela presença de grande n ú m e r o de microcro-m o s s o microcro-m o s . Estes c r o microcro-m o s s o microcro-m o s são de d i f í c i l caracterização mor-fológica e m é t r i c a , daí advêm as dificuldades de d i s c r i m i n a ç ã o indi-v i d u a l . É c o m u m o aparecimento de m i c r o c r o m o s s o m o s
metafási-cos divididos pelo c e n t r ô m e r o e a s s i m podem ser computados
co-mo dois e l e m e n t o s , ou, em outros casos devido à proximidade de dois m i c r o c r o m o s s o m o s , estes po-dem ser considerados como um único e l e m e n t o .
A observação das posições cen-t r o m é r i c a s nos macrocromosso-mos a u t o s s ò m i c o s destas duas es-pécies (Tab. II e IV) mostrou que entre os dez pares há somente três que são semelhantes c o m re-lação às posições c e n t r o m é r i c a s : c r o m o s s o m o s 5, 6 e 7.
Os autores enfatizam que o cro-m o s s o cro-m o Z é u cro-m dos elecro-mentos que apresenta maior conservado-r i s m o ao longo da evolução caconservado-rio- cario-t í p i c a na classe A v e s , sendo o quarto ou o quinto e m tamanho e exibindo notável consistência em m o r f o l o g i a e conteúdo de DNA (Ohno et ai., 1964; Lucca, 1978). No entanto, e m P. cayana o
cro-m o s s o cro-m o Z é s u b cro-m e t a c ê n t r i c o e o segundo elemento em o r d e m de grandeza (Tab. II), enquanto em C. major ele é m e t a c ê n t r i c o e o quar-to e m tamanho (Tab. IV).
Os dados c i t o g e n é t i c o s , dispo-níveis até o m o m e n t o , mostram que o c r o m o s s o m o W, nas dife-rentes espécies de aves já cario-tipadas, varia bastante, tanto sob o ponto de v i s t a m o r f o l ó g i c o quan-to m é t r i c o . Em algumas espécies chega a ser tão grande quanto o c r o m o s s o m o Z (Renzoni & Vegni-Talluri, 1966; Bulatova & Panov,
1973; Boer, 1975, 1976; A u e t a l . ,
Wal-drigues & Ferrari, 1979), p o r é m , nunca se m o s t r o u maior do que este elemento (Lucca, 1978).
O c r o m o s s o m o W de P. cayana
é subtelocêntrico e o d é c i m o se-gundo em tamanho, enquanto o de
C. m a j o r é s u b m e t a c ê n t r i c o e o
sé-t i m o elemensé-to do cariósé-tipo e m es-cala de grandeza. Portanto, são restritas as igualdades m o r f o l ó g i cas e métricas dos m a c r o c r o m o s -somos destas duas e s p é c i e s de C u c u l i f o r m e s .
Ao se compararem os dados morfológicos e m é t r i c o s dos ma-c r o ma-c r o m o s s o m o s de P. cayana e C. m a j o r , c o m os das demais
es-pécies de C u c u l i f o r m e s já carioti-padas, observa-se que há maior semelhança c o m os de C. m a j o r
do que c o m os de P. c a y a n a .
As-s i m , c o m exceção de P. c a y a n a ,
todas as demais espécies c a r i o t i -padas apresentam o maior macro-cromossomo como m e t a c ê n t r i c o . Os três Crotophaginae ( C . m a j o r , C. ani (Waldrigues & Ferrari, 1982)
e G. guira (Waldrigues & F e r r a r i ,
1980; Lucca, 1974), a p r e s e n t a m concordâncias m o r f o l ó g i c a s e m relação aos três maiores autosso-m o s . Soautosso-mente P. cayana
apresta c r o m o s s o m o s t e l o c ê n t r i c o s entre os maiores m a c r o c r o m o s s o -mos; o que faz c o m que e s t a es-pécie, pertencente à S u b f a m í l i a (Phaenicophaeinae) tenha c a r i ó t i po c o m características m o r f o l ó g i -cas diferentes do das e s p é c i e s analisadas das outras duas
Subfa-mílias (Crotophaginae e C u c u l i
-n a e ) . Porta-nto, seria i n t e r e s s a n t e
a análise c a r i o t í p i c a de e s p é c i e s
das subfamilias C e n t r o p o d i n a e e
N e o m o r p h i n a e , que ainda não
a p r e s e n t a m quaisquer dados cario-lógicos, bem c o m o , de maior nú-mero de espécies das Subfami-lias Cuculinae e Phaenicophaei-nae, quando então poderiam obter-se evidências do relacionamento c a r i o t í p i c o entre as espécies da O r d e m C u c u l i f o r m e s . A única es-pécie da S u b f a m í l i a Crotophaginae a ser cariotipada é C r o t o p h a g a s u l -c i r o s t r i s .
SUMMARY
Two species of birds of the Cuculidae Family, Piaya cayana (Phaenicophaeinae)
and Crotophaga major (Crotophaginae),
captured in Manaus (AM, Brazil) region have been cytogenetically studied. The chromosomes were preparared for 4 hours in culture using bone narrow cells. The karyotypic analyses were done by convencional methods from which nume-rical, morphological and metrical data of the chromosomes were drawn. The mean size of the macrochromosomes, the Re-lative Lengths (PR) and Arm Ratios (B) were calculated on the basis of the mea-surements obtained. A system on de-creasing order of size was used to arran-ge the chromosomes, designating each chromosome individually. The macrochro-mosomes were morphologically classified according to Levan et al. (1964). All
chromosomes that had PR equal or bigger than 4% were considered as macrochromosomes. P. cayana has 2n
complement of 76 chromosomes, from which 22 are considered as macrochro-mosomes. The Z chromosome is subme-tacentric and the second of the karyo-type, however the W is subtelocentric
chromosomes, from which 22 are macro-chromosomes. The Z chromosome is me-tacentric and the fourth of the karyotype, but the W chromosome is submetacentric and the sixth in size.
REFERENCIAS BIBLIOGRÁFICAS
AU, W.; FECHHEIMER, N.S.; SOUKUP; S. 1975 — Identification of the sex
chro-mosomes in the bald eagle.
Cañad. J. Genet. Cytol., 17 :
187-191.
BLOOM, S E .
1969 — A current list of chromosome numbers and variations for species of the avian subclass Carinatae. J. Hered., 60:
217-220.
BOER, L.E.M. de
1975 — Karyological heterogeneity in the Falconiformes (Aves). Ex-perientia, 31:1138-1139.
1976 — Chromosome complements of 16 species of Falconiformes (Aves) and the Karyological relationships of the order. Ge-nética, 46:77-113.
BULATOVA, N.S. & PANOV, E.N. 1973 — Comparative analysis of
ka-ryotypes of 18 species Family Turdidae (Aves). Cariologia,
26:229-244.
HERMAN, E., & HOLYNSKI. R.
1970 — Obecny stan badan nad cyto-gentyka ptakow. II. Ewolucya Chromosomowa. Przeglad Zool.
14:159-173.
LEVAN. A . ; FREDGA, K.; SAMDBERG.
A.R.
1964 — Nomenclature for centromerlc position on chromosomes. He-reditas, 52:201-220.
LUCCA, E.J. de
1974 — Cariótipo de 14 espécies de aves das Ordens Cuculiformes, Galliformes, Passeriformes e Tinamiformes. Rev. Bras. Pesq. Méd. Biol., 7:253-263.
LUCCA, E.J. de & AGUIAR, M.L.R. de 1976 — Chromosomal evolution in
Co-lumbiformes (Aves). Caryolo-gy, 29:59-68.
LUCCA, E.J. de
1978 — Determinação cromossômlca do sexo em aves. Ciênc. Cult.,
30:791-798.
OHNO, S.; STENIUS, C ; CHRISTIAN, L C. ; BEÇAK, W.; BEÇAK, M.L.
1964 — Chromosomal uniformity in the subclass carinatae. Chro-mosoma, 15:280-282.
RAY-CHAUDHURI, R.
1967 — Mitotic and meiotic chromoso-mes of the Koel Eudynamys scolopacea scolopacea. Nu-cleus, 10:179-189.
1973 — Cytotaxonomy and Chromoso-me Evolution in Birds. In: Cy-totaxonomy and Vertebrate Evolution. (Chiarelli, A.B. &
Capanna, E., eds.) (London) LTD., London, Academic Press Inc., pp. 425-483.
RAY-CHAUDHURI. R.; SHARMA, T.; RAY-CHAUDHURI, S.P.
1969 — A comparative study of the chromosomes of birds. Chro-mosoma, 26:148-168.
RENZONI, A. & VEGNI-TALLURI, M. 1966 — The Karyograms of some
Fal-coniformes and Strigiformes.
Chromosoma, 20:133-150.
SRB, V.
TAKAGI, N. & SASAKI, M.
1974 — A phylogenetic study of bird karyotypes. Chromosoma, 46:
91-120. YAMASHINA, M.Y.
1946 — The chromosomes of the cuckoo, the budgerigar, the little ringed plover and Chine-se bamboo pheasant. La
Kro-mosomo, 1:18-23.
WALDRIGUES, A. & FERRARI. I.
1979 — Estudo cromossômico em Cro-tophaga ani (Aves). Ciênc. Cult., 31:03-G.1.6.
1980 — Estudo cromossômico em Gui-ra guisa (Aves). Ciênc. Cult.,
32:267-G.1 .6.
1982 — Karyotypic study of Cuculiform birds. I. Karyotype of the Smocth — billed ani (Croto-phaga ani). Rev. Brasil. Genet.,
5(1): 121-129.