Genotypic diversity of the Killer Cell Immunoglobulin-like Receptors (KIR) and their HLA class I Ligands in a Saudi population
Texto
Imagem
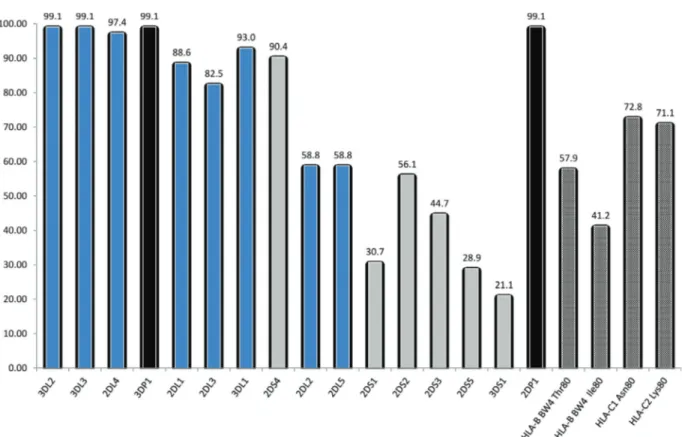
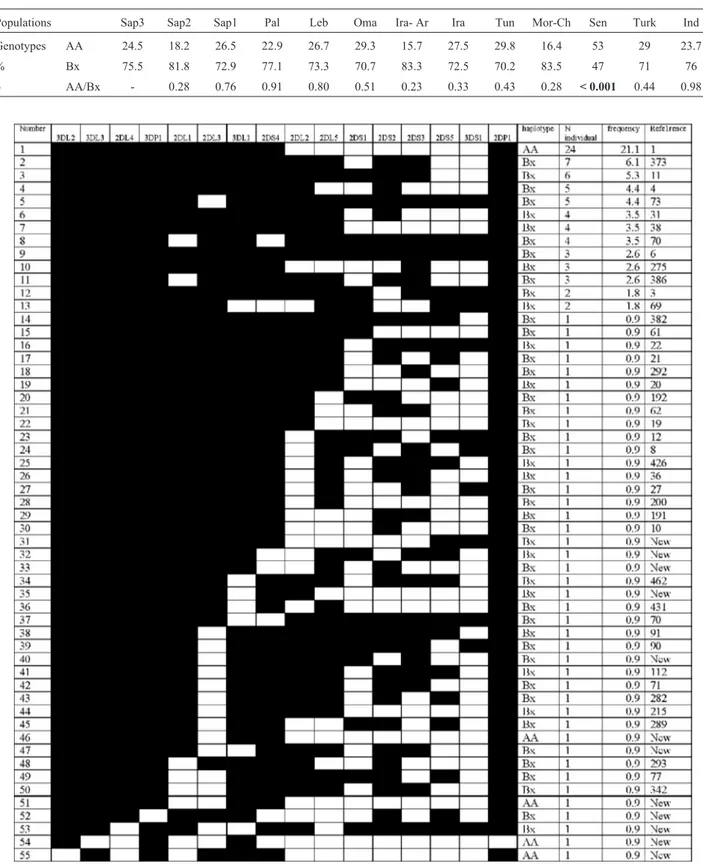
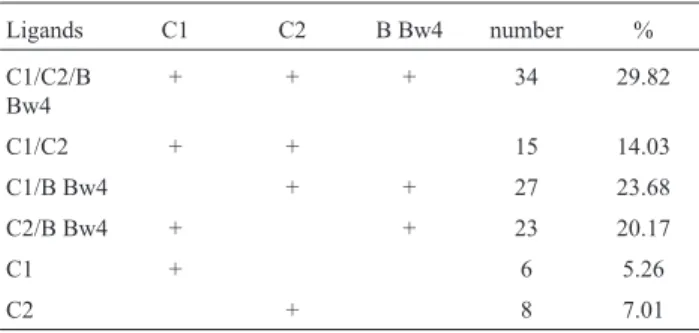
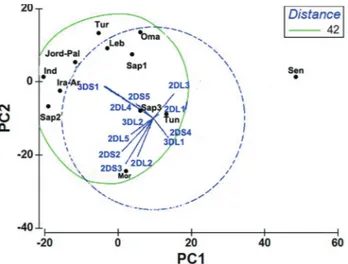
Documentos relacionados
Several studies have shown that certain combinations of killer immunoglobulin-like receptors and human leukocyte antigens (in both donors and recipients) can affect the chances
Through HLA class I (HLA-A* and -B*) and class II (HLA- DRB1*) allele typing from the registry of volunteers (REDOME) of Bauru, São Paulo, we aimed to estimate the frequencies of
A diversidade de frequência dos haplótipos KIR em certas populações sugere que alguns indivíduos apresentam diferentes níveis de proteção contra algumas doenças e o balanço
The objective of this study was to establish a correlation between the frequency of major histocompatibility complex class I/II alleles and killer immunoglobulin-like receptor
in Killer Cell immunoglobulin-like receptors (KIR) and their HLA-C ligands have consistently been associated with viral clearance and chronic infection (Khakoo et al.. Even though
Em suma, conclui-se com este estudo - o qual abordou os projetos e as primeiras experiências da “Luiz de Queiroz”, em Piracicaba, e relacionou-os com outras tentativas
O desenvolvimento pessoal e profissional, o modo como gerem as dificuldades pessoais e relacionais com o doente e família no contexto de fim de vida, são
Distribution of human leukocyte antigen (HLA) class I antigens in the H.. Evaluation of human leukocyte antigen class I and II antigens in Helicobacter pylori -positive
