ww w . r e u m a t o l o g i a . c o m . b r
REVISTA
BRASILEIRA
DE
REUMATOLOGIA
Review
article
Role
of
HIF-1
␣
signaling
pathway
in
osteoarthritis:
a
systematic
review
Javier
Fernández-Torres
a,b,∗,
Gabriela
Angélica
Martínez-Nava
a,
María
Concepción
Gutiérrez-Ruíz
b,
Luis
Enrique
Gómez-Quiroz
b,
Marwin
Gutiérrez
a,baInstitutoNacionaldeRehabilitación“LuisGuillermoIbarraIbarra”,LaboratoriodeLíquidoSinovial,MexicoCity,Mexico
bUniversidadAutónomaMetropolitanaIztapalapa,ProgramadeDoctoradodeCienciasBiológicasydelaSalud,MexicoCity,Mexico
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received11February2016 Accepted28April2016 Availableonline5August2016
Keywords:
Hypoxiainduciblefactor-1␣
HIF-1␣signalingpathway Geneticpolymorphisms Osteoarthritis
a
b
s
t
r
a
c
t
Osteoarthritis(OA)isthemostcommonformofarthritisandisfrequentlydiagnosedand managedinprimarycare;itischaracterizedbylossofarticularhyalinecartilage,whichisa uniqueconnectivetissuethatphysiologicallylacksbloodvessels.Articularcartilagesurvives inamicroenvironmentdevoidofoxygen,whichisregulatedbyhypoxiainduciblefactor (HIF-1␣).HIF-1␣isconsideredthemaintranscriptionalregulatorofcellularanddevelopmental responsetohypoxia.Todate,therelevanceofHIF-1␣intheassessmentofcartilagehas increasedsinceitsparticipationisessentialinthehomeostasisofthistissue.Takinginto accountthenewemerginginsightsofHIF-1␣inthescientificliteratureinthelastyears, wefocusedthepresentreviewonthepotentialroleofHIF-1␣signalingpathway inOA development,especiallyinhowsomegeneticfactorsmayinfluencethemaintenanceor breakdownofarticularcartilage.
©2016ElsevierEditoraLtda.ThisisanopenaccessarticleundertheCCBY-NC-ND license(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Papel
da
via
de
sinalizac¸ão
do
HIF-1
␣
na
osteoartrite:
revisão
sistemática
Palavras-chave:
Fatorinduzívelporhipóxia1-␣
Viadesinalizac¸ãodoHIF-1␣
Polimorfismosgenéticos Osteoartrite
r
e
s
u
m
o
Aosteoartrite(OA)éaformamaiscomumdeartriteefrequentementeédiagnosticadae gerenciadanaatenc¸ãoprimária;écaracterizadaporperdadacartilagemarticularhialina, umtecidoconjuntivoúnicoquefisiologicamentecarecedevasossanguíneos.Acartilagem articularsobreviveemum microambientedesprovidodeoxigênio,queé reguladopelo fatorinduzívelporhipóxia-1␣(HIF-1␣).OHIF-1␣éconsideradooprincipalregulador tran-scricionaldarespostacelularededesenvolvimentoàhipóxia.Naatualidade,arelevância
∗ Correspondingauthor.
E-mail:[email protected](J.Fernández-Torres). http://dx.doi.org/10.1016/j.rbre.2016.07.008
doHIF-1␣naavaliac¸ãodacartilagemtemaumentado,jáqueasuaparticipac¸ãoéessencial nahomeostasedessetecido.ConsiderandoasnovasperspectivasemergentesdoHIF-1␣na literaturacientíficanosúltimosanos,foca-seapresenterevisãonopotencialpapeldavia desinalizac¸ãodoHIF-1␣nodesenvolvimentodaOA,especialmentenomodocomoalguns fatoresgenéticospodeminfluenciarnamanutenc¸ãoourupturadacartilagemarticular.
©2016ElsevierEditoraLtda.Este ´eumartigoOpenAccesssobumalicenc¸aCC BY-NC-ND(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Introduction
Osteoarthritis(OA)isacommonchronicconditionaffecting millionsofpeopleworldwideandisaconsiderablecauseof disability.1Itisthemostcommonrheumaticdisease,everyage canbeaffectedbutprevalenceincreasesdramaticallywithage withagreaterincidenceinsubjectsbetween40and50years old.2
The joints involved are characterized by a breakdown and loss of articular cartilage that leads to a decrease in the joint space and friction between the bones causing swelling,chronicpain,functionalimpairment,deformityand disability.3–6
Todate,thehypoxiainduciblefactor-1alpha(HIF-1␣)has increaseditsrelevanceintheassessmentofcartilagesinceits participationisessentialinthehomeostasisofthistissue.7 ArticularcartilageisahypoxictissueinwhichHIF-1␣isof pivotalimportanceforsurvivalandgrowthofchondrocytes duringcartilagedevelopmentaswellasenergygenerationand matrixsynthesisofchondrocytesinbothhealthyand patho-logicalconditions.8,9Byusingmicroarrays,itwasalsoshown thatHIF-1␣isexpressedinhumanfetalchondrocytes,which meansthatthistranscriptionfactorisessentialfor develop-mentandmaintenanceofcartilage.10
Theviability ofchondrocytesiscompromisedbyseveral phenomenasuchasoxidativestress,inflammatorymediators, biochemicalinjuryandhypoxicconditions.Theavascularity ofcartilage tissue hasallowed establishingwell conserved mechanismswherethechondrocytescansurviveundersuch conditions.Itisnote,thatunderhealthyconditions,oxygen concentration in articular cartilage varies from 0.5 to 10% (∼4–70mmHg,respectively).11,12Whenoxygenconcentration
decreasesandenvironmentturnsincreasinglyhypoxic, HIF-1␣ plays a critical role to maintain homeostasis, through inductiontheexpressionofavarietyofgenesencoding pro-teinstoincreasetheavailabilityofoxygenand nutrientsto homeostaticlevels.8,13
Takingintoaccountofthesepiecesofinformationandthe recentgrowinginterestinHIF-1␣inrheumaticdiseases,14,15 wefocusedthepresentreviewonthepotentialroleofHIF-1␣
inOA,especiallyinassomegeneticfactorsmayinfluencein themaintenanceorbreakdownofarticularcartilage.
Methods
Literaturereviewcriteriaandsearchstrategy
AllrelevantliteratureinthefieldofHIF-1␣andOA,published inthelast15yearswasreviewed.Thesearchincludedoriginal
articlesconcerninghumansand/oranimalmodelspublished betweenJanuary2000andDecember2015.Toidentifyall avail-ablestudies,adetailedsearchpertainingtoHIF-1␣andOAwas conducted according toPRISMA (PreferredReporting Items for Systematic reviewsand Meta-Analyses) guidelines.16 A systematicsearchwasperformedintheelectronicdatabase (PubMed)usingthefollowingMeshsearchtermsinall pos-siblecombinations:“HIF-1␣”or“osteoarthritis”or“articular cartilage” or “HIF-1␣ polymorphisms” or “HIF-1␣ signaling pathway”or“hypoxia”or“rheumaticdiseases”,andthe com-bined phrasesinorder toobtain allgeneticstudies on the relationship of genetic polymorphisms of HIF-1␣ signaling pathwayassociatedwithOA.Inaddition,thereferencelists ofall retrieved articleswere manually reviewed.Two inde-pendentauthors(JFTandGAMN)analyzedeacharticleand performedthedataextractionindependently.Discrepancies wereresolvedbyconsensus.
Inclusionandexclusioncriteria
Weexcludedfromthisreviewthefollowingtypesof publica-tions:articlesnotpublishedinEnglish,casereports,clinical trials, and letters to the editor that were purely commen-tary.Searchresultswerescreenedtoavoidduplicates.Titles, abstracts,andfullreportsofarticlesidentifiedwere systemat-icallyscreenedwithregardtoinclusionandexclusioncriteria.
Results
Todate,ithascompiledimportantinformationabouttherole ofHIF-1␣inrheumaticdiseases.Approximately5599 publi-cationswereidentifiedinPubMeddatabasebetweenJanuary 2000andDecember2015.Theresultsofthesearchstrategy areillustratedinFig.1.
GeneralconceptsofOAgenesisandgenetics
Search terms entered in PubMed database: “HIF-1α”or “osteoarthritis” or “articular cartilage”
or “HIF-1α polymorphisms” or “HIF-1α signaling pathway” or “hypoxia” or “rheumatic diseases”,
and the combined phrases in order to obtain all genetic studies on the relationship of
genetic polymorphisms of signaling pathway of HIF-1α associated
with osteoarthritis.
5599 studies identified in database search
383 potential eligible studies identified
76 full-tex articles assessed for elegibility
5214 studies excluded: not published in english, case reports, clinical trials, letters to the editor that
were purely commentary, published before 2000.
309 studies excluded not related to given topic.
Fig.1–Representationofthesearchstrategy.
AlthoughthepathophysiologyofOA isnotfully charac-terized, several candidate genes have been reported to be associatedwith OA susceptibility. Fernández-Moreno et al. determined thatdespite themultifactorial nature ofOA,it notfollowstheMendelianinheritancepatterns,mostlikely bythealterationsofgeneinteractions.20Theyanalyzed differ-entgeneslocatedondifferentchromosomes,andtheresults revealedthecomplexityofthisfield.Table1showssomegenes andtheir relationshipwiththe differentphenotypesofOA describedinthisstudy.
Meulenbeltpublishedastudyaimedtodeterminewhich signalingpathwaysweremostimportanttothedevelopment ofOA.6Themostcommonpathwaysorgeneswerethe7q22
locuscontainingmultiplepotentialgenes,thegrowth
differ-entiationfactor5(GDF5)gene,frizzledrelatedprotein(FRZB) gene,thedeiodinaseiodothyronine,typeII(DIO2)geneand
theSMAD3gene.
The genetic bases in OA can further refine the under-standingofthegenotype–phenotyperelationship,throughthe presenceofsinglenucleotidepolymorphisms(SNP).Agenetic polymorphismcanbeapivotbetweenamechanismof resis-tanceorsusceptibilityinadisease.Geneticpolymorphisms thataffectingacodingorregulatorysequenceand produce majorchanges inproteinstructureor mechanism of regu-lationofexpression,canresultindifferentphenotypes.23,24 InTable2weshowsomeSNPswithphenotypeswell estab-lished.
Functionandstructureofarticularcartilage
Articularcartilageisahighlyspecializedtissueofjoints;its principalfunctionistoprovideasmooth,lubricatedsurface forarticulationandtofacilitatethetransmissionofloadswith alowfrictionalcoefficient.Injurytoarticularcartilageis rec-ognizedasacauseofsignificantmusculoskeletalmorbidity. Theuniqueandcomplexstructureofarticularcartilagemakes treatment, andrepairorrestorationofitsdefects challeng-ing forthepatient,thesurgeon,andthephysicaltherapist. Thepreservationofarticularcartilageishighlydependenton maintainingitsorganizedarchitecture.40
Table1–GenesassociatedwiththedevelopmentofOA.
Gene Chromosome Phenotypic
manifestation
COL2A1, COL11A1, COL11A2
1,6,12 EarlyonsetOA
COL9A1 6 Earlyonsetofknee
OA
MATN3 2 Earlyonsetofhand
andkneeOA
COMP 19 EarlyonsetofhipOA
hip
COL1A1 17 ReductionofhipOA
inwomen
BMP2 20 Reductionofknee
OAinwomen
TGFB1 19 OA
FRZB,IL4R 2,16 HipOAinwomen
IL1,ASPN,TIMP3 2,9,22 HipandkneeOA
IL6 7 HipOA
AGC1 15 HandOA
VDR 12 Arthritisinseveral
joints
ERA 6 OAinwomen
ADAM12,LRCH1, TNA
3,10,13 KneeOA
CILP 15 KneeOAinmen
CALM1 14 HipOAinJapanese
population
IGF-1 12 IncreasedriskofOA
IL17 6 Susceptibilityto
developOA
TakenandmodifiedfromFernández-Morenoetal.20
onlyrelatedtothelossofECMbutalsochondrocytesdeath. Chondrocytedeathbyapoptosis,necrosis,chondroptosis,or combinationoftheseprocesseshasbeenimplicatedinthe pathogenesisofOA.47
The
HIF-1
␣
system
HIF-1␣isatranscriptionalfactorencodedbytheHIF1Agene locatedwithin chromosome14q21-24 and isformed by15
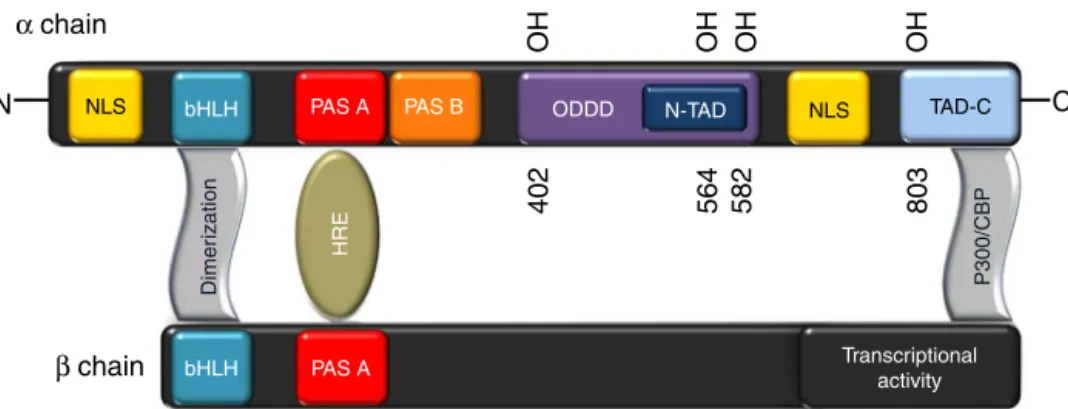
exons;HIF-1␣consistsof826aminoacidsandithasa molec-ular weight of 120kDa.48 HIF-1␣ is a heterodimer of two chains, alpha chain (regulated by oxygen) and beta chain, botharrangedinadoublehelix(basichelix–loop–helix,bHLH). Therearetwonuclearlocalizationsignal(NLS),butonlythat foundintheC-terminalpositionitisresponsibleforthe accu-mulationofHIF-1␣inthenucleus.IntheN-terminalregion, islocatedthebHLHandPER-ARNT-SIMA(PASA)domains, necessaryfordimerizationandDNAbindingthroughhypoxia responseelements(HRE).Finally,theactivesiteofthis pro-teinisanoxygendependentdegradationdomain(ODDD)that functionsasanoxygensensor(Fig.2).49–51
Under normoxia and in the presence of Fe2+ and
2-oxoglutarate, the specific proline residues 402and 564are hydroxylatedonODDDdomainbyprolyl-hydroxylases(PHDs) oxygen dependents, to form a complex with the factor of vonHippel-Lindau(VHL);inturn,thiscomplexbindsto ubiq-uitin (Ub)andit subsequently degraded inthe proteasome (Fig.3).
An external stimulus cellular, as a growth factor that binds to its receptor tyrosine kinase, triggers a cas-cade of signaling pathways within the cell. For example, vascular endothelial growth factor (VEGF) activates the phosphatidylinositol-3-kinase (PI3K) and mitogen-activated proteinkinase(MAPK)pathways(ERK1andERK2).52PI3K acti-vatestheserine/threoninekinase(AKT),andAKTactivatesthe FKBP12 rapamycin-associated protein,mTOR, RAFT (FRAP), whichinducestheexpressionofHIF-1␣.
Underconditionsofreducedoxygen(hypoxia),PHDs activ-itydecreases,whichstabilizesHIF-1␣andaccumulatesinthe cytoplasm tobephosphorylated by MAPK.53–55 Once phos-phorylated, HIF-1␣ ittranslocatestothenucleusand binds toHIF-1subunit(also knownas arylhydrocarbon nuclear translocator,ARNT)toformthe[HIF-1␣/HIF-1]complex.This complexthroughtheHRE,bindstospecificDNAsequences 5′TAGCGTGH3′presentinpromoterregionsofgenesfor
sub-sequentexpression.47,55,56
Someofthesetargetgenesincludenitricoxidesynthase 2 (NOS2), vascular endothelialgrowth factor (VEGF), eryth-ropoietin (EPO), someglucose transporters (GLUT1, GLUT3), Insulin-likegrowthfactortype2(IGF2),whichpotentiallyacts inorder tomaintain thechondroprotective functions chal-lenged by the detrimental conditions occurring in the OA
Table2–Singlenucleotidepolymorphisms(SNP)associatedwithOAprocesses.
Gene/dbSNPrsId Phenotype Firstauthor[Reference]
WISP1rs2929970 SusceptibilitytospinalOAinwoman Uranoetal.25
RAGErs2070600 SusceptibilitytokneeOA Hanetal.26
DVWArs7639618 SusceptibilitytokneeOA Zhangetal.27
ACErs4343,rs4362 SusceptibilitytokneeOA Qingetal.28
MATN3rs8176070 SusceptibilitytoOA Guetal.29
DIO2rs225014 SusceptibilitytoOA Meulenbeltetal.30
ADAMTS14rs4747096 kneeOAinwoman(Thaipopulation) Poonpetetal.31
ADAM12rs1871054 IncreasedriskofOA Wangetal.32;Kernaetal.33
HIF1Ars11549465 Protectiveroleinthelossofarticularcartilage Fernándezetal.34 IL6rs1800796 ProtectivefactorforhipandkneeOAintheelderly Fernandesetal.35 IL16rs11556218,rs4072111,rs4778889 DecreasedkneeOArisk Luoetal.36;Liuetal.37
α
chain
β
chain
N
NLS bHLH NLS TAD-CC
bHLH
PAS A
PAS A Transcriptional
activity
HRE
Dimer
ization
P300/CBP
PAS B ODDD N-TAD
OH
OH
OH
OH
402
564
582
803
Fig.2–Structureofhypoxiainduciblefactor1-␣(HIF-1␣).TheNH2-terminalofHIF-1␣andHIF-1consistsofbHLH(basic
helix–loop–helix)andPAS(Per-ARNT-Simhomology)domainsthatarerequiredforheterodimerizationandDNAbinding.
TheCOOH-terminalofHIF-1␣(residues531–826)containstwotransactivationdomains(TADs).Theshorthalf-lifeofHIF-1␣
undernonhypoxicandposthypoxicconditionsisduetorapidubiquitinationandproteasomaldegradation.HIF-1␣residues
400–600,thisregionwasdesignatedtheoxygen-dependentdegradationdomain(ODDD).
jointenvironment(Fig.4).8,54–59Thisrelationshipamong dif-ferentgenesrendersthecloserelationofHIF-1␣withseveral pathologies.60–64
Genetic
polymorphisms
in
the
HIF-1
␣
system
and
their
importance
in
OA
Totrytoexplainsimplytheinteractionofgenetic polymor-phisms associatedwithHIF-1␣ with importanceinOA, we dividedthissystemintothreestages:(1)genesthatactivate
HIF-1␣;(2)proteinsthatdirectlyinteractwithHIF-1␣;and(3) genesdrivenbyHIF-1␣.
GenesthatactivateHIF-1˛system
HIF-1␣activationcanstartbythebindingofdifferentproteins totheirreceptorsonthecellmembrane;theseproteinsmaybe enzymes,growthfactors,interleukinsorothertypemolecules; thattheycanbeaffectedbythepresenceofgenetic polymor-phismswhichmaybeassociatedwiththedevelopmentofOA. Yang etal. analyzedtheeffect ofreceptor advanced glyca-tionendproducts(RAGE)polymorphismsonsusceptibilityto
Normoxia
Ubiquitination
Hidroxilation
O
2
PHD
OH
OH
HIF-1α VHL HIF-1α HIF-1α
VHL
OH Ub
Ub
OH
Proteasome
Degradation
Fig.3–HIF-1␣activityundernormoxicconditions.Undernormoxiaconditions,thespecificprolineresidues402and564on
ODDDdomainarehydroxylatedbyoxygendependentprolyl-hydroxylases(PHDs)thatleadstotheformationofacomplex
withthevonHippel-Lindau(VHL)factor;whichinturn,bindstoubiquitin(Ub)andissubsequentlydegradedbythe
VEGF
PI3K
AKT
FRAP PHD
P
P MAPK
HIF-1α HIF-1α
Transcription
HIF-1α
HIF-1α
HRE
HIF-1β
HIF-1α
VEGF
NOS II/III
NQO1
MMPs
EPO
ET-1
GLUT-1 tranporterGlucose
Mitogen Erythropoiesis
Matrix remodeling Antioxidant Vasodilation extravasation Angiogenesis
O
2Hypoxia
Fig.4–HIF-1␣activityunderhypoxicconditions.Underhypoxicconditions,HIF-1␣isstabilizedandphosphorylatedby
MAPK;oncephosphorylated,HIF-1␣translocatestothenucleusandbindstoHIF-1subunittoformthe[HIF-1␣/HIF-1]
complex.Thiscomplex,throughtheHRE,bindstospecificDNAsequence5′TAGCGTGH3′presentinthepromoterregionsof
severalgenesfortheirsubsequentactivation.
andseverityofOA inaHanChinesepopulation. RAGE par-ticipatesinregulatinginflammation,evenintheproduction ofmatrixmetalloproteinases(MMPs).MMP-1degrades carti-lage,whichmayresultinOAdevelopment.Theyfoundthat twopolymorphisms(rs1800625andrs2070600)inRAGEgene showed a significant association between OA patients and healthycontrols(OR=0.42,p=0.016,andOR=2.78,p=0.047, respectively).65InthestudyperformedbyHanetal.,26 they evaluatedthat the presenceofrs2070600 polymorphism in
RAGEgeneininteractionwithobesity,maydeterminethe sus-ceptibilityofkneeOA.
Swellametal.reportedapotentialinfluenceof intereukin-1receptorantagonist(IL-1RA) genepolymorphism onknee OA risk. IL-1 gene issupposed tobe involvedin the carti-lagedestructionprocess.Inthisregard,interleukin-1receptor antagonist (IL-1RA) competing with IL-1 for binding to its receptormayactasaninhibitorofcartilagebreakdown.They conductedacase–controlstudywithkneeOApatients,and concludedthatIL-1RN*2allele representasignificantfactor influencingtheseverityandcourseofkneeOA(p=0.002).66
Fernandes et al., analyzed the influence of pro-inflammatory cytokine IL-6 with severity and functional status of OA in elderly individuals, and determined that
the rs1800796 polymorphism is a protective factor for the
presenceandseverityofhipandkneeOAintheelderly.The individualsharboringtheCallelehavelowerprevalenceand severityof OA when comparedto individuals without this polymorphism.35
Meanwhile,interleukin-16 (IL-16),apleiotropiccytokine, playsafundamentalroleininflammatorydiseases.Liuetal. determinedthat, comparedwiththeC/C genotype,the C/T
genotypeincreasedtheriskofprimarykneeOAinrs4072111
of IL-16 gene (OR=1.83); however, compared with the T/T
genotype, the T/G genotype decreased the risk of primary
kneeOAinrs11556218polymorphism(OR=0.37).37Likewise,
Luo et al. evaluated the same polymorphisms and deter-minedthesamebehavior.36TheseresultssuggestthatIL-16 gene polymorphisms are associated with the risk of knee OA.
Finally,thereareothergenesassociatedwithactivationof HIF-1␣,suchasPIK3R1,AKT2,GSK3B,IL6,anditwillbe neces-sarytoexploretheirgeneticvariantsandtodeterminetheir participationinthedevelopmentofOA.
ProteinsthatinteractdirectlywithHIF-1˛
ThestabilizationofHIF-1␣inthecytoplasmbasicallydepends on the hydroxylation atspecific sites within ofthe ODDD domain. However, the presence of genetic polymorphisms couldalterthestructuralpropertiesofthetranscripts,andthis mayinfluencethesusceptibilityorresistancetodiseases,such aswasanalyzedbyUchanzkaetal.inautoimmunediseases associatedwithHLA-B*27allele.67
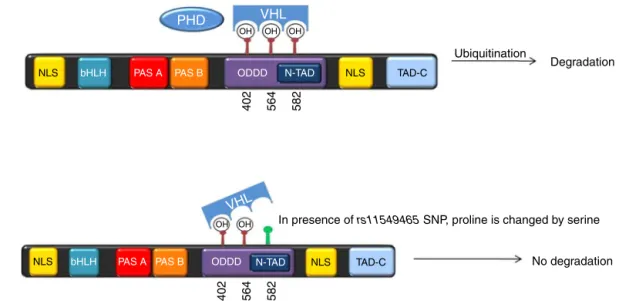
In2003Tanimotoetal.demonstratedthatthesubstitution ofprolinebyserineinthe582(P582S)position,duetothe pres-enceofsinglenucleotidepolymorphism(rs11549465)within
HIF1Agene,enhancesitstranscriptionalactivity,56duetoan
PHD
bHLH
bHLH
PAS A
PAS A
PAS B
PAS B
ODDD
ODDD
N-TAD
N-TAD
VHL
VHL
NLS
NLS
NLS
NLS
In presence of rs11549465 SNP, proline is changed by serine
No degradation
402 564 582
402 564 582 OH
OH OH
OH OH
TAD-C
TAD-C
Ubiquitination
Degradation
Fig.5–Thepresenceofrs11549465polymorphism,generatesachangeofprolinebyserine,whichconfersgreaterstability
totheHIF-1␣proteinduetopoorinteractionbetweenVHLandhydroxylationsiteswithintheODDD.
or T allele OR=0.2, p=0.004).34 This phenomenon may be explainedbythefactthatthepresenceofthispolymorphism confersgreaterstabilitytotheHIF-1␣proteinduetopoor inter-actionbetweenVHLandhydroxylationsiteswithintheODDD (Fig.5).
Otherpolymorphismmedicallyimportant,withincreased transcriptionalactivitywithinHIF1Agenethatalsowastested byTanimoto etal., isAla588Thr (rs11549467).Similarly, we evaluatethispolymorphisminpatientswithOA,butdidnot findhimassociated.However,thisopensthepossibilitytobe evaluatedinotherpopulationsinordertobetterunderstand theirinfluenceinOA.
Additionally, it have been described polymorphisms for
EGLN1(alsoknownasPHD2,prolyl-hydroxylase2),VHLand
HIF1AN(HIF1Ainhibitorfactor)genesinteractingdirectlywith
HIF-1␣, but not have been evaluated in OA and that may be of clinical importance in this disease. Some studies in rheumatoidarthritis(RA)demonstratedthe expressionand regulationofprolylhydroxylasedomain(PHD)enzymesand factor-inhibitingHIF-1␣(FIH-1),whichregulatecellular HIF-1␣ levels. It isknown that RAis characterizedby hypoxia andtheexpressionofhypoxia-inducibletranscriptionfactors (HIFs),whichcoordinatecellularresponsestohypoxia.Muz etal.conductedthisstudyinRAfibroblast-likesynoviocytes, and concluded that PHD-2 is the majorhydroxylase regu-latingHIFlevelsandthe expressionofangiogenicgenesin arthriticcells.PHD-2appearstoregulateresponsesrelevantto arthritisviaHIF-␣,highlightingthemajorimportanceofthis enzymeinhypoxia-andangiogenesis-dependent inflamma-torydiseases.69Thismakesussupposethatthepresenceof geneticpolymorphismsofthesegenesmayaffectthestability ofHIF-1␣contributingimportantlyintheOAdisease.
GenesdrivingbyHIF-1˛
OA chondrocytes are metabolically active, displaying increasedsynthesisoftypeIIcollagen.Incomparisonwith
healthycartilage,OAarticularchondrocytesexhibitincreased
in vivosynthesis ofcollagenprolyl-4-hydroxylase typeII, a pivotal enzyme in collagen triple helix formation.70 Once stabilizedHIF-1␣inthecytoplasm,severaldownstreamgenes areexpressedinordertorestoremultiplecomponentsofthe extracellularmatrix.Thepresenceofgeneticpolymorphisms inthesegenesmayalterthefunctionofspecificproteinsthat restorejointtissues,promotingthedevelopmentofOA.
Raine et al.performed anallelic expression analysisof theOA susceptibilitygeneCOL11A1inhumanjointtissues. By using RNA from OA cartilage of individuals undergo-ing elective joint replacement for OA of the hip (total hip replacement, THR) or of the knee (total knee replace-ment, TKR), they observed a significant allelic expression imbalance(AEI)atrs1676486(p<0.0001)withtheT-allele cor-relatingwithreducedCOL11A1expression.AEIatrs1676486
is a risk factor for lumbar disk herniation, but not for OA.71
Rodríguez-Fontenla et al. conducted a meta-analysis of nine GWASto assess candidate genesforassociation with OA, and only 2 of the 199 candidate genes (COL11A1 and
VEGF) were associated with OA in the meta-analysis. Two polymorphisms inCOL11A1 gene(rs4907986andrs1241164) showed association with hip OA in the combined analysis (OR=1.12,p=1.29×10−5,andOR=0.82,p=1.47×10−5,
respec-tively);and thers4908291withthesexstratified analysisin womenonly(OR=0.87,p=1.29×10−5).Other polymorphism
inVEGFgene(rs833058)showedassociationwithhipOAin
menonly(OR=0.85,p=1.35×10−5).72
Theoxygenandnutrientssupplytoarticularcartilageis by diffusion from the synovial fluid. The role of vascular endotheliumgrowth factor (VEGF)iscritical for angiogene-sisinsubchondralbone.8Thereareonlyfewstudiesrelated withgenevariantsofVEGFgenethatmaycontributetothe developmentandprogressofOA.
Sánchezetal.evaluatedtwopolymorphismsofVEGFgene,
withhealthy controls, but didnot findassociation.73 Yuan etal.conductedameta-analysistoorderunderstandthe rela-tionshipbetweenthepathogenesisofOAandtheexpression levelsofVEGFinmultiplediseasetissuesinthesepatients. Atotalof11case–controlstudies,containing302OApatients and195healthycontrols,demonstratethatVEGFexpression levelsinOApatientsaresignificantlyhigherthanhealthy con-trols(standardizedmeandifference=1.18,95%CI:4.91–9.11,
p<0.001),andtheselevelsstronglycorrelatewiththe patho-genesisofosteoarthritis.74
OneofthemechanismsofcartilagedegradationinOAis enzymaticproteolysisoftheextracellularmatrixby metallo-proteinases.MMP-1,producedbychondrocytesandsynovial cells,isamajorproteaseoftheMMPsfamily.
Barlaset al.evaluated threepolymorphisms inthe pro-moter of matrix metalloproteinase-1 (MMP-1), MMP-2 and
MMP-9genesinpatientswithkneeOAand comparedwith
ethnicallymatchedcontrol.Theyfoundsignificantdifferences betweenthe groups regardingthe genotype distribution of
MMP-1polymorphism(p=0.001).Thefrequenciesof1G/1Gand
1G/2GgenotypesweresignificantlyhigherinthekneeOAthan inthecontrols(p=0.002,andp=0.006,respectively).In addi-tion,1G allele frequencyofMMP-1 gene washigher inthe patientsthaninthecontrolgroup(p=0.0001).Thegenotype distributionsandallelefrequenciesofMMP-2andMMP-9gene polymorphismsdidnotdifferbetweentheOAandthecontrol groups(p>0.05).Thesefindingssuggestthatthe−16071G/2G
polymorphism(rs1799750)intheMMP-1genemaycontribute tosusceptibilitytokneeOA.75
Similarly, Lepestos et al. evaluated the rs1799750 poly-morphisminMMP-1 gene,but theydidnotfindsignificant associationincrudeanalysis;however,aftermultiple logis-ticregression analysis, 1G/2G was associatedwithreduced oddsofknee OA by75% in males,comparedtogenotypes 1G/1G+2G/2G,adjustingforageandBMI(adjustedOR=0.25,
p=0.035).76
Finally, Honsaweketal. analyzedthe MMP-3 (rs3025058,
−1612)polymorphism withknee OApatients.The5Aallele
frequencywasindicatedas15.5%,and6Aallelewasas84.5% inOApatients,whereasitwas10–90%inthecontrolgroup. Accordingly,thepresentstudyhasindicatedthatthe−1612
5A/6ApolymorphismgenotypesofMMP-3genepromoterdo notplayaroleinthedevelopmentofOA.77
TheseresultssuggestthattheMMPsfamilyactivityis influ-encedbypresenceofgeneticvariants,whichwouldbreakthe balancebetweensynthesisanddegradationofextracellular matrix,andthisconditionmaycontributetosusceptibilityof OA.
Nitricoxide(NO)isessentialinthemaintenanceofvascular tonusandthepresenceofendothelialimpairment(reduced vascularrelaxation)maysuggestaproblemregardingtheNO pathway.NOisproducedbyendothelialNOsynthase(eNOS), andits productioncanbe influencedbypolymorphisms of theeNOSgene.78Todate,nostudieshavebeendonerelated withOAandgeneticpolymorphismsofNOS;however,several geneticpolymorphismsintheeNOSgeneareassociatedwith thepathogenesisofRA.
ThelevelofNOisincreasedinRApatients,andastudy suggestedthat NOcan regulatethe balance ofTh1/Th2 in
autoimmune diseases, and it was a key mediator of apo-ptosis withinrheumatoid arthritis joints.Anet al. studied twopolymorphismsoftheeNOSgene(rs2070244,T-786C;and
rs1799983,G894T)inpatientswithRA,andobservedthat
indi-vidualswiththe−786CCgenotypehaveanincreasedriskof
RA.79
Brenol et al. evaluated the T-786Cpolymorphism inRA patientscomparingwithextraarticularmanifestations.They found that the Callele was significantly associated(p cor-rected=0.032), suggesting the participation of the T-786C polymorphismoftheeNOSgeneandRA.80
TheseresultswemakesupposethateNOSgene polymor-phismscanhaveanimportantimpactondevelopmentofOA; itwillbenecessarytoexplorethesegeneticvariantsto cor-roborateit.
The erythropoietin-mediated bone marrow response to anemia is under the control of hypoxia-inducible factors (HIFs),themasterregulatorsofoxygenandironhomeostasis. Thehypoxiccharacteristicsofjointcartilagemakethat HIF-1␣participatesactivelyallowingtranscriptionoftargetgenes. Erythropoietin(EPO)geneisexpressedafterofanincreaseof HIF-1␣oncytoplasm.81,82 Buttodate,noscientificevidence thatsupportsthepossibleassociationbetween erythropoie-tinandcartilagelossinOA;andevenmore,thepresenceof polymorphisms inEPOgenecouldrepresentsanimportant factorassociatedwithriskofOA.
Clinical
relevance
of
PHDs
inhibitors
as
potential
therapeutic
targets
in
OA
Todate,HIF-1inhibitorsareclassifiedbytheirHIFinhibitory mechanism,includingaffectingonHIF-1␣proteinlevel, HIF-1 dimerization,HIF-1 DNA binding,or HIF-1␣ transcription of target genes.83 Due to the results described above, the mainobjectiveforachievingatherapeuticeffectinthe treat-ment ofOA atcartilage level, could stabilizeHIF-1␣in the cytoplasm sothat it can induce the expressionof restora-tiongenes.Naturally,therearegeneticpolymorphismsthat increasethetranscriptionalactivityofHIF-1␣incomparison withcommonisoform.Attheexperimentallevel,has evalu-atedtheDimethyloxallylGlycine(DMOG),apotentinhibitor ofprolyl-hydroxylases.84 Theendogenous HIF-1␣ levelscan be increasedby the suppression of PHD activity, eitherby reducing the cellular oxygen level or bycombining the Fe (II) competitively. DMOG is a cell permeable, competitive inhibitorofthePHDs.DMOGisananalogof2-oxoglutarate, and inthis wayitinhibits notonlythe HIFprolylbut also asparaginylhydroxylases.Besidethatitispredictedtoinhibit other members of 2-oxoglutarate-dependent dioxygenases. TherearethreeHIF-prolylhydroxylasesknowninmammals, and they are encodedbyseparate genes: PHD1, PHD2, and
PHD3.Likeall2-oxoglutarate-dependentdioxygenases,PHDs require oxygen for hydroxylation, as well as tricarboxylic acidcycleintermediate,2-oxoglutarate(␣-ketoglutarate),iron (Fe2+), and ascorbate as cofactors.When oxygen levelsare
OtherinhibitorsofPHDswithpotentialbeneficialeffects aredeferoxamine(DFO)andcobaltchloride(CoCl2),aniron
chelatorandacompetitiveinhibitorofiron,respectively;they areroutinelyusedbothinvitroandinvivotoinhibitPHDs activ-itybycompetingforendogenousiron(II).Otherironchelators, such as ciclopirox olamine, and competitive inhibitors of iron,such asCu2+, Zn2+,and Mn2+,are alsoused asPHDs
inhibitors.53
Likewise,therearestudiesshowingthatHIF-1␣andits tar-getgeneVEGF,arecriticalregulatorsofangiogenic–osteogenic coupling.7,87 In addition to chondrocytes, osteoblasts also express HIF-1␣ and promote skeletal vascularization dur-ing endochondralboneformation; manipulationofthe HIF system via pharmacological or genetic approaches, is an attractive strategy for treating hypoxic diseases, including skeletaldiseasessuchassubchondralboneinflammation dur-ingOA.53
Conclusion
Is worth mentioning that several associations’ studies betweenSNPsandOA diseaseremainunconfirmedor con-troversialduetobiasinpatientenrollingcriteriadifferences inOAaffectedjointsites,inclassificationandstagingmode.
Cartilage destruction in OA mediated by catabolic enzymesandchondrocytedeath,includingapoptosisand/or autophagy,alsocontributetothepathogenesis.Thestudies showed that the expression of HIF-1 is increased in OA cartilagetomediatetheresponseofchondrocytestohypoxia; HIF-1 acts as a survival factor by enhancing extracellular matrixsynthesisand inhibitingapoptosis88;HIF-1servesto regulatebothautophagyandapoptosisandHIF-1isofpivotal importanceincartilage homeostasis.89 Also,it willalso be necessarytoexploreotherisoforms ofHIF,suchasHIF-2␣, which it seems to havethe opposite effectto HIF-1␣. The HIF-2 proteinactsas abrakeonthe autophagy-accelerator functionofHIF-1,andpromoteschondrocytehypertrophy,a terminaldifferentiationstatecharacterizedbyauniquegene expressionprogram,includingtypeXcollagenandthetypeII collagen-degradingproteaseMMP-13.9
Althoughfurtherstudiesshouldelucidatetheexact mech-anismofHIF-1inOAthecurrentevidenceinducetoconsiderit asapromisingapproachtothetreatmentofOA.However,the resultshavebeenreportedtoindicatethatgeneticmarkers couldcontributetotheunderstandingofthenaturalhistory ofthisdisease.
Conflict
of
interest
Theauthorsdeclarenoconflictsofinterest.
Acknowledgments
Theauthors wish tothankCarlosAguilar-González forhis valuablesupportinthedesignofFigs.3and4.
r
e
f
e
r
e
n
c
e
s
1.VosT,FlaxmanAD,NaghaviM,LozanoR,MichaudC,Ezzati M,etal.Yearslivedwithdisability(YLDs)for1160sequelaeof 289diseasesandinjuries1990–2010:asystematicanalysisfor theGlobalBurdenofDiseaseStudy2010.Lancet.
2012;380:2163–96.
2.FicheraC,PappalardoA,TrioloG,GalloM,ValentiniG, BagnatoG.Epidemiologyandriskfactorsinosteoarthritis: literaturereviewdatafromOASISstudy.Reumatismo. 2004;56:169–84.
3.KomatsuM,KamimuraM,NakamuraY,MukaiyamaK, IkegamiS,UchiyamaS,etal.Rapidbonedestructionina patientwithkneeosteoarthritis.Acasereportandreviewof theliterature.ClinCasesMinerBoneMetab.2014;11: 232–5.
4.SchiphofD,vanMiddelkoopM,deKlerkBM,OeiEH,Hofman A,KoesBW,etal.Crepitusisafirstindicationof
patellofemoralosteoarthritis(andnotoftibiofemoral osteoarthritis).OsteoarthritisCartilage.2014;201: 631–8.
5.MusumeciG,AielloFC,SzychlinskaMA,DiRosaM, CastrogiovanniP,MobasheriA.OsteoarthritisintheXXIst century:riskfactorsandbehavioursthatinfluencedisease onsetandprogression.IntJMolSci.2015;16(3):
6093–112.
6.MeulenbeltI.Osteoarthritisyear2011inreview:genetics. OsteoarthritisCartilage.2012;20:218–22.
7.SchipaniE,RyanHE,DidricksonS,KobayashiT,KnightM, JohnsonRS.Hypoxiaincartilage:HIF-1alphaisessentialfor chondrocytegrowtharrestandsurvival.GenesDev. 2001;15:2865–76.
8.PfanderD,CramerT,HypoxiaSwobodaB.HIF-1alphain osteoarthritis.IntOrthop.2005;29(1):6–9.
9.CoimbraIB,JimenezSA,HawkinsDF,Piera-VelazquezS, StokesDG.Hypoxiainduciblefactor-1alphaexpressionin humannormalandosteoarthriticchondrocytes.
OsteoarthritisCartilage.2004;12(4): 336–45.
10.StokesDG,LiuG,CoimbraIB,Piera-VelazquezS,CrowlRM, JiménezSA.Assessmentofthegeneexpressionprofileof differentiatedanddedifferentiatedhumanfetalchondrocytes bymicroarrayanalysis.ArthritisRheum.2002;46(2):
404–19.
11.PfanderD,SwobodaB,CramerT.TheroleofHIF-1alphain maintainingcartilagehomeostasisandduringthe
pathogenesisofosteoarthritis.ArthritisResTher.2006;8:104. Epub2006Jan18.
12.StröbelS,LoparicM,WendtD,SchenkA,CandrianC, LindbergR,etal.Anabolicandcatabolicresponsesofhuman articularchondrocytestovaryingoxygenpercentages. ArthritisResTher.2010;12:R34.
13.GrimmerC,PfanderD,SwobodaB,AingnerT,MuellerL, HenningF,etal.Hypoxia-InducibleFactor1␣isinvolvedinthe prostaglandinmetabolismofosteoarthriticcartilagethrough up-regulationofmicrosomalprostaglandinEsynthase1in articularchondrocytes.ArthritisRheum.2007;56:
4084–94.
14.DistlerK.Hypoxiaandangiogenesisinrheumaticdiseases.Z Rheumatol.2003;62Suppl2:II43–5.
15.WuL,HuangX,LiL,HuangH,XuR,LuytenW.Insightson biologyandpathologyofHIF-1␣/-2␣,TGF/BMP,
Wnt/-catenin,andNF-(Bpathwaysinosteoarthritis.Curr PharmDes.2012;18(22):3293–312.
meta-analyses:thePRISMAstatement.PLOSMed. 2009;6:e1000097.
17.GuilakF.Biomechanicalfactorsinosteoarthritis.BestPract ResClinRheumatol.2011;25:815–23.
18.LeeKM,ChungCY,SungKH,LeeSY,WonSH,KimTG,etal. Riskfactorsforosteoarthritisandcontributingfactorsto currentarthriticpaininSouthKoreanolderadults.Yonsei MedJ.2015;56:124–31.
19.DeFilippisL,GulliS,CaliriA,RomanoC,MunaóF,Trimarchi G,etal.Epidemiologyandriskfactorsinosteoarthritis: literaturereviewdatafromOASISstudy.Reumatism. 2004;56:169–84.
20.FernándezM,RegoF,BlancoF.Geneticsinosteoarthritis. ReumatolClin.2007;3Supl3:S13–8.
21.ChapmanK,ValdesAM.GeneticfactorsinOApathogenesis. Bone.2012;51:258–64.
22.ShiD,ZhengQ,ChenD,ZhuL,QinA,FanJ,etal.Association ofsingle-nucleotidepolymorphisminHLAclassII/IIIregion withkneeosteoarthritis.OsteoarthritisCartilage.
2010;18:1454–7.
23.SealA,GuptaA,MahalaxmiM,AykkalR,SinghTR,
ArunachalamV.Tools,resourcesanddatabasesforSNPsand indelsinsequences:areview.IntJBioinformResAppl. 2014;10:264–96.
24.KnezK,SpasicD,JanssenKP,LammertynJ.Emerging technologiesforhybridizationbasedsinglenucleotide polymorphismdetection.Analyst.2014;139:
353–70.
25.UranoT,NarusawaK,HosoiT,OuchiY,NakamuraT,InoueS. Associationofasinglenucleotidepolymorphisminthe WISP1genewithspinalosteoarthritisinpostmenopausal Japanesewomen.JBoneMinerMetab.2007;25:
253–8.
26.HanZ,LiuQ,SunC,LiY.Theinteractionbetweenobesityand RAGEpolymorphismsontheriskofkneeosteoarthritisin Chinesepopulation.CellPhysiolBiochem.2012;30: 898–904.
27.ZhangR,YaoJ,XuP,JiB,VoegeliG,HouW,etal.Association betweengeneticvariantsofDVWAandosteoarthritisofthe kneeandhip:acomprehensivemeta-analysis.IntJClinExp Med.2015;8(6):9430–7.
28.QingZ,YeJ.AssociationbetweenACEpolymorphismsand osteoarthritissusceptibility.IntJClinExpPathol.
2015;8(6):7391–6.
29.GuJ,RongJ,GuanF,JiangL,TaoS,GuanG,etal.MATN3gene polymorphismisassociatedwithosteoarthritisinChinese hanpopulation:acommunity-basedcase–controlstudy. ScientificWorldJ.2012;2012:656084.
30.MeulenbeltI,MinJ,BosS,RiyaziN,HouwingJ,SlagboomE, etal.IdentificationofDIO2asanewsusceptibilitylocusfor symptomaticosteoarthritis.HumMolGenet.2008;17: 1867–75.
31.PoonpetT,HonsawekS,TammachoteN,KanitnateS, TammachoteR.ADAMTS14genepolymorphismassociated withkneeosteoarthritisinThaiwomen.GenetMolRes. 2013;12:5301–9.
32.WangL,GuoL,TianF,HaoR,YangT.Analysisofsingle nucleotidepolymorphismswithinADAM12andriskofknee osteoarthritisinaChineseHanpopulation.BiomedResInt. 2015;2015:518643.
33.KernaI,KisandK,TammAE,KummJ,TammAO.Two single-nucleotidepolymorphismsinADAM12geneare associatedwithearlyandlateradiographicknee osteoarthritisinEstonianpopulation.Arthritis. 2013;2013:878126.
34.Fernández-TorresJ,Hernández-DíazC,Espinosa-MoralesR, Camacho-GalindoJ,Galindo-SevillaNdelC,López-MacayA,
etal.Polymorphicvariationofhypoxiainduciblefactor-1A (HIF1A)genemightcontributetothedevelopmentofknee osteoarthritis:apilotstudy.BMCMusculoskeletDisord. 2015;16:218.
35.FernandesMT,FernandesKB,MarquezAS,CólusIM,Souza MF,SantosJP,etal.Associationofinterleukin-6gene polymorphism(rs1800796)withseverityandfunctional statusofosteoarthritisinelderlyindividuals.Cytokine. 2015;75:316–20.
36.LuoSX,LiS,ZhangXH,ZhangJJ,LongGH,DongGF,etal. Geneticpolymorphismsofinterleukin-16andriskofknee osteoarthritis.PLoSOne.2015;10:e0123442.
37.LiuZ,MaL,QiuS,JiaT.Geneticpolymorphismsof interleukin-16areassociatedwithsusceptibilitytoprimary kneeosteoarthritis.IntJClinExpMed.2015;8:1401–5, eCollection2015.
38.PanF,TianJ,WinzenbergT,DingC,Jones.Association betweenGDF5rs143383polymorphismandknee osteoarthritis:anupdatedmeta-analysisbasedon23,995 subjects.BMCMusculoskeletDisord.2014;15:
404.
39.TawonsawatrukT,ChangthongT,PingsuthiwongS,Trachoo O,SauraT,WajanavisitW.Ageneticassociationstudy betweengrowthdifferentiationfactor(GDF5)polymorphism andkneeosteoarthritisinThaipopulation.JOrthopaedSurg. 2011;Res6:2–5.
40.SophiaFoxAJ,BediA,RodeoSA.Thebasicscienceofarticular cartilage:structure,composition,andfunction.SportsHealth. 2009;1:461–8.
41.MarianiE,PulsatelliL,FacchiniA.Signalingpathwaysin cartilagerepair.IntJMolSci.2014;15:8667–98.
42.ChenJL,DuanL,ZhuW,XiongJ,WangD.Extracellularmatrix productioninvitroincartilagetissueengineering.JTransl Med.2014;12:88.
43.FindlayDM,AtkinsGJ.Osteoblast–chondrocyteinteractionsin osteoarthritis.CurrOsteoporosRep.2014;12:127–34.
44.IwamotoM,OhtaY,LarmourC,Enomoto-IwamotoM.Toward regenerationofarticularcartilage.BirthDefectsResCEmbryo Today.2013;99:192–202.
45.MaldonadoM,NamJ.Theroleofchangesinextracellular matrixofcartilageinthepresenceofinflammationonthe pathologyofosteoarthritis.BiomedResInt.2013;2013: 284873.
46.vanderKraanPM.Osteoarthritisyear2012inreview:biology. OsteoarthritisCartilage.2012;20:1447–50.
47.ZhangFJ,LuoW,LeiGH.RoleofHIF-1␣andHIF-2␣in osteoarthritis.JointBoneSpine.2015;82:144–7.
48.LobodaA,JozkowiczA,DulakJ.HIF-1andHIF-2transcription factors–similarbutnotidentical.MolCells.2010;29:
435–42.
49.GörlachA.RegulationofHIF-1alphaatthetranscriptional level.CurrPharmDes.2009;15:3844–52.
50.SemenzaGL.HIF-1:mediatorofphysiologicaland pathophysiologicalresponsestohypoxia.JApplPhysiol. 2000;88:1474–80.
51.MaxwellPH.Hypoxia-induciblefactorasaphysiological regulator.ExpPhysiol.2005;90:791–7.
52.FukudaR,HirotaK,FanF,JungYD,EllisLM,SemenzaGL. Insulin-likegrowthfactor1induceshypoxia-induciblefactor 1-mediatedvascularendothelialgrowthfactorexpression, whichisdependentonMAPkinaseandphosphatidylinositol 3-kinasesignalingincoloncancercells.JBiolChem. 2002;277:38205–11.
54.CatrinaSB,OkamotoK,PereiraT,BrismarK,PoellingerL. Hyperglycemiaregulateshypoxia-induciblefactor-1alpha proteinstabilityandfunction.Diabetes.2004;53:
3226–32.
55.ZhouJ,HaraK,InoueM,HamadaS,YasudaH,MoriyamaH, etal.Regulationofhypoxia-induciblefactor1byglucose availabilityunderhypoxicconditions.KobeJMedSci. 2008;53:283–96.
56.TanimotoK,YoshigaK,EguchiH,KaneyasuM,UkonK, KumazakiT,etal.Hypoxia-induciblefactor-1a
polymorphismsassociatedwithenhancedtransactivation capacity,implyingclinicalsignificance.Carcinogenesis. 2003;24:1779–83.
57.YamadaN,HorikawaY,OdaN,IizukaK,ShiharaN,KishiS, etal.GeneticvariationintheHypoxia-InducibleFactor-1␣ geneisassociatedwithtype2diabetesinJapanese.JClin EndocrinolMetab.2005;90:5841–7.
58.TzouvelekisA,NtoliosP,KaramerisA,KoutsopoulosA, BoglouP,KoulelidisA,etal.Expressionofhypoxia-inducible factor(HIF)-1a-vascularendothelialgrowthfactor
(VEGF)-inhibitorygrowthfactor(ING)-4-axisinsarcoidosis patients.BMCResNotes.2012;5:654.
59.Sartori-CintraAR,MaraCS,ArgoloDL,CoimbraIB.Regulation ofhypoxia-induciblefactor-1␣(HIF-1␣)expressionby interleukin-1(IL-1),insulin-likegrowthfactorsI(IGF-I)and II(IGF-II)inhumanosteoarthriticchondrocytes.Clinics(São Paulo).2012;67:35–40.
60.KonacE,DoganI,OnenHI,YurdakulAS,OzturkC,VarolA, etal.Geneticvariationsinthehypoxia-inducible
factor-1alphageneandlungcancer.ExpBiolMed. 2009;234:1109–16.
61.LiP,CaoQ,ShaoPF,CaiHZ,ZhouH,ChenJW,etal.Genetic polymorphismsinHIF1Aareassociatedwithprostatecancer riskinaChinesepopulation.AsianJAndrol.2012;14: 864–9.
62.AlidoostiM,GhaediM,SoleimaniA,BakhtiyariS,Rezvanfard M,GolkhuS,etal.Studyontheroleofenvironmental parametersandHIF-1Agenepolymorphismincoronary collateralformationamongpatientswithischemicheart disease.ClinBiochem.2010;44:1421–4.
63.BahadoriB,UitzE,MayerA,HarauerJ,DamK, Truschnig-WildersM,etal.Polymorphismsofthe
hypoxia-induciblefactor1geneandperipheralarterydisease. VascMed.2010;15:371–4.
64.NagyG,KovacsR,KereszturiE,SomogyiA,SzekelyA,Nemeth N,etal.Associationofhipoxiainduciblefactor-1alphagene polymorphismwithbothtype1andtype2diabetesina Caucasian(Hungarian)sample.BMCMedGenet.2009;10: 79.
65.YangHY,ChuangSY,FangWH,HuangGS,WangCC,Huang YY,etal.EffectofRAGEpolymorphismsonsusceptibilityto andseverityofosteoarthritisinaHanChinesepopulation:a case–controlstudy.GenetMolRes.2015;14:
11362–70.
66.SwellamM,MahmoudMS,SamyN,GamalAA.Potential influenceofinterleukin-1receptorantagonistgene polymorphismonkneeosteoarthritisrisk.DisMarkers. 2010;28:299–305.
67.Uchanska-ZieglerB,LollB,FabianH,HeeCS,SaengerW, ZieglerA.HLAclassI-associateddiseaseswithasuspected autoimmuneetiology:HLA-B27subtypesasamodelsystem. EurJCellBiol.2012;91:274–86.
68.HongJM,KimTH,ChaeSC,KooKH,LeeYJ,ParkEK,etal. Associationstudyofhypoxiainduciblefactor1alpha(HIF-1 alpha)withosteonecrosisoffemoralheadinaKorean population.OsteoarthritisCartilage.2007;15:
688–94.
69.MuzB,LarsenH,MaddenL,KiriakidisS,PaleologEM.Prolyl hydroxylasedomainenzyme2isthemajorplayerin regulatinghypoxicresponsesinrheumatoidarthritis. ArthritisRheum.2012;64:2856–67.
70.GrimmerC,BalbusN,LangU,AignerT,CramerT,MüllerL, etal.RegulationoftypeIIcollagensynthesisduring osteoarthritisbyprolyl-4-hydroxylases:possibleinfluenceof lowoxygenlevels.AmJPathol.2006;169:491–502.
71.RaineEV,DoddAW,ReynardLN,LoughlinJ.Allelicexpression analysisoftheosteoarthritissusceptibilitygeneCOL11A1in humanjointtissues.BMCMusculoskeletDisord.2013;14: 85.
72.Rodriguez-FontenlaC,CalazaM,EvangelouE,ValdesAM, ArdenN,BlancoFJ,etal.Assessmentofosteoarthritis candidategenesinameta-analysisofninegenome-wide associationstudies.ArthritisRheumatol.2014;66: 940–9.
73.Sánchez-EnríquezS,Torres-CarrilloNM,Vázquez-Del MercadoM,Salgado-GoytiaL,Rangel-VillalobosH, Mu ˜noz-ValleJF.Increaselevelsofapo-A1andapoBare associatedinkneeosteoarthritis:lackofassociationwith VEGF-460T/Cand+405C/Gpolymorphisms.RheumatolInt. 2008;29:63–8.
74.YuanQ,SunL,LiJJ,AnCH.ElevatedVEGFlevelscontributeto thepathogenesisofosteoarthritis.BMCMusculoskelet Disord.2014;15:437.
75.BarlasIO,SezginM,ErdalME,SahinG,AnkaraliHC,Altintas ZM,etal.Associationof(−1607)1G/2Gpolymorphismof
matrixmetalloproteinase-1genewithkneeosteoarthritisin theTurkishpopulation(kneeosteoarthritisandMMPsgene polymorphisms).RheumatolInt.2009;29:
383–8.
76.LepetsosP,PampanosA,KanavakisE,TzetisM,KorresD, PapavassiliouAG,etal.AssociationofMMP-1−16071G/2G
(rs1799750)polymorphismwithprimarykneeosteoarthritis intheGreekpopulation.JOrthopRes.2014;32:
1155–60.
77.HonsawekS,MalilaS,YuktanandanaP,TanavaleeA, DeepaisarnsakulB,ParviziJ.AssociationofMMP-3(-1612 5A/6A)polymorphismwithkneeosteoarthritisinThai population.RheumatolInt.2013;33:435–9.
78.CookeGE,DoshiA,BinkleyPF.Endothelialnitricoxide synthasegene:prospectsfortreatmentofheartdisease. Pharmacogenomics.2007;8:1723–34.
79.AnJD,LiXY,YuJB,ZhaoY,JinZS.Associationbetweenthe eNOSgenepolymorphismsandrheumatoidarthritisriskina northernChinesepopulation.ChinMedJ(Engl).
2012;125:1496–9.
80.BrenolCV,ChiesJA,BrenolJC,MonticieloOA,FranciscattoP, BirrielF,etal.EndothelialnitricoxidesynthaseT-786C polymorphisminrheumatoidarthritis:associationwith extraarticularmanifestations.ClinRheumatol.2009;28: 201–5.
81.HaaseVH.Regulationoferythropoiesisbyhypoxia-inducible factors.BloodRev.2013;27:41–53.
82.TortiL,TeofiliL,CapodimontiS,NuzzoloER,IachininotoMG, MassiniG,etal.Hypoxia-induciblefactor-1␣(Pro-582-Ser) polymorphismpreventsirondeprivationinhealthyblood donors.BloodTransfus.2013;11:553–7.
83.XiaY,ChoiHK,LeeK.Recentadvancesinhypoxia-inducible factor(HIF)-1inhibitors.EurJMedChem.2012;49:
24–40.
85.BrocatoJ,ChervonaY,CostaM.Molecularresponsesto hypoxia-induciblefactor1␣andbeyond.MolPharmacol. 2014;85:651–7.
86.SmithTG,TalbotNP.Prolylhydroxylasesandtherapeutics. AntioxidRedoxSignal.2010;12:431–3.
87.DanisA.MechanismofbonelengtheningbytheIlizarov technique.BullMemAcadRMedBelg.2001;156(1–2):107–12.
88.KohMY,PowisG.Passingthebaton:theHIFswitch.Trends BiochemSci.2012;37:364–72.


![Fig. 4 – HIF-1␣ activity under hypoxic conditions. Under hypoxic conditions, HIF-1␣ is stabilized and phosphorylated by MAPK; once phosphorylated, HIF-1␣ translocates to the nucleus and binds to HIF-1 subunit to form the [HIF-1␣/HIF-1]](https://thumb-eu.123doks.com/thumbv2/123dok_br/19082418.494175/6.918.178.752.102.450/activity-hypoxic-conditions-conditions-stabilized-phosphorylated-phosphorylated-translocates.webp)