Bolm Inst. oceanogr . , S Paulo, 28(2) :1-15, 1979
COPEPODS OF THE GENUS
OITHONAFROM CANANÉIA REGION (LAT.
25°07'S)
LO NG .
47°56'W)
VE RA LÚCIA FONSECA & MARIA SCINTILA DE ALMEIDA PRADO Ins tituto Oceanográfico da Universidade de são Paulo
Synopsis
Some eC'oZ.ogi ca l aspects of two congeneric species of Cy do poid copepods~ OmoYla
ov~ and O. oligohalirta~ have been studied
for
the first t i me in the estuarine region of Cananéia. The spatial distribution and seasona l variation ofbothspecies of Odhorta are studied in relation to salinity~ tempera ture and disso l ved oxygen. The results obtained are compared to previous UJork done in the a:rea and to s i milar-studies undertaken elsewhere.In troduction
The genus Omorta is widely dis tributed and occurs in alI kinds of marine en-vironments (estuarine, coas~al and
oce-anic waters). It is not usually recorded i n zooplankton samples due to its small si ze in relation to the mesh aperture of th e gause commonly used in zooplankton net s . The species of CdhoYla are very simi lar and often misidentified (Grice, 1960) •
The purpose of this paper is the s tudy o f the spatial and temporal distribution of OdhoYla oligohaliYla and O. o v~ in the estuarine region of Ca-nanéia, South littoral of the State of s ão Paulo, Brazil (Lat. 25°07'S - Long. 47 ° 56 ' W) .
Copepods are the dominant enimals of the plankton in the Cananéia region and among them the genus OdhoYla is one of the mos t abund ant (Teixeira, Tundisi & Kut ner , 1965).
The t wo species of Odhona found in Canané ia water s are morphologically almost alike (Fonseca & Bjornberg, 1976; Fonseca , 1976) but have different
patterns of distribution in relation to salini t y .
Tempe r a ture , salinity and dissolved oxygen wer e considered the main environ-ment al fac t ors in the study of the distribu tion of the species along the estuary .
The sal inity of estuarine waters suffers wide flu ctuations (Kinne, 1967). The seasonal v ariation of salinity is remarkable i n es tuaries especially in Summer. Resides this, it has a semi-diurnal sali nity variation due to tidal
Publ. YlQ 444 do
IrlJ.:Jt.
oc.e.aYlogtt. da U.óp.movements (Fraga, 1972).
The area delimited for this study is very narrow but suffers a very strong influence of tidal movements.
Many papers have been published on plankton from the Cananéia region (Tei-xeira, Tundisi
&
Kutner, 1965; Teixeira, Tundisi & Santoro Ycaza, 1969; Tundisi, 1970; Prado, 1972; Kutner, 1972; Tundi-si, 1972) but this is the first on conge-neric species of copepods living there. It is also the first time that zoo-plankton samples were collected ~n ariver of this complex estuarine region. Material and methods
Plankton samples were taken monthly (June, 1973 - May, 1974) at three fixed stations as follows: the first station was located in the upper waters of Ta-quari river, the second at its mouth and the third in Trapandé Bay (Fig. 1). The depths of the stations I, 11 and 111 were respectively 1.70, 3.80 and 4.00 m. The influence of coastal water is great-er at St. 111 and smallgreat-er at St. I. Surface plankton samples were taken by means of a zooplankton net type WP3
(Fraser, 1968) with its size reduced to half and mesh aperture 75 ~m. AlI
hauls were taken with the ship running at slow speed, against the tide, during 3 minutes.
The first and second stages of O.
ou-gohaliYla and O. O l ' ~ were obtaine d from ovigerous females in the laboratory (Fonseca & Prado, in prepar ation) .
Salinity determinations were made by the Harvey Method and dissolved oxygen by the Winkler Method. The temperature was recorded with a
2 Bolm Inst. ocea nogr. , S Paulo , 28(2), 1979
~---An aliquot of 5 ml of each haul was obtained by means of a Stempel pipette and alI the nauplii stages and the adults of both sexes were counted up. Copepo.di tes were disregarded due to the diff icult i es of quick identif i cat ion .
The number of nauplii and aduits was plotted against salinity and temp erature of each haul (T. S.
P.).
\ ~.' 'C:$.'~ o~
2v-r
~~~
Km
Fig. 1. Sampl ing location. Cananéia region.
Resu1ts
1. Hydrographical data
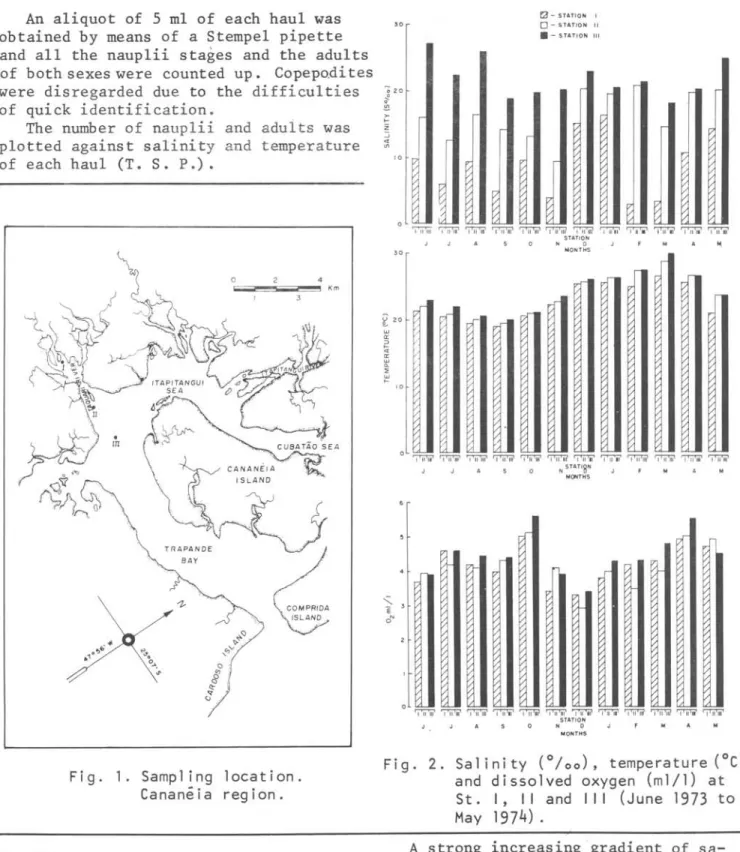
Figure 2 shows the data of temperature, salinity and dissolved oxygen of St. I, 11 and 111, from June 1973 to May 1974. A small increasing gradient of temper-ature from St. I to St . 111 was observed. These differences can be explained by the sampling time and the position of the stations. Samples were always taken earlier in the morning at the St. I and later on at the St. 11 and 111. The Station
r
is aIs o more sheltered by coastal vegetation than St.rr
andrrr.
,o
10
r:a -S TA TIO ,", I
0-5T ATIO '" 11
• - 5 TAT IO N I II
0'iti""iil~~~7iI'iii'~7ii)jjl~~~'I7ii'''i''7r'ii'
5TATION
o ~
'o MONT HS
o ~~7ii"ijj1~~~~7iiíi'~ '~~'I"ii"iW
N SUTl8'" A 1111
MQNTHS
O~7ií'ii'~7ii"ii''IW~7ii''ii'~7ti'iii'7iT'ii?r'j'T'jj'TjiIifTiiTiiiT
J .
Fi g. 2.
STATlO N
• o •
M ONTHS
Sa 1 i n i ty (0/00). temperature (OC) and dissolved oxygen (ml/l) at St. I. I I and I I I (June 1973 to May 1974).
A strong increasing -gradient of sa-linity was registered from St. I to 11 and 111, due to tidal movements and to the influence of the river discharge.
The dissolved oxygen in the same day does not show significative differences at the three stations.
The samples were always taken in the morning at any tidal leveI. Figure 3
shows the tidal leveI in each haul.
2. Distribution and seasonal variation
of
OUhona ova1..Á..,6and
O. oUgohaLina.2.1. OUhona. ova1..Á..,6
a-FONSECA
&
PRADO: O~tho~a: Cananéia r-I """"
• '00
~ 180
TIME
18 / 4 /1 .74
" ,
'00
""
'50
ê ,O<)
""
'50 '4.
""
• • 10 • 12
'1/ 2 /1'74 1&/ 10 /1 91"
-l.
TIME
1'/ "'1174
I
, , ITATI_bundant. A total of 1,921,000 adu1ts and 796,800 naup1ii was found in a11
the samp1es (Tab. I).
Horizontal distribution:
this specieswas more abundant at the St. 11 and 111 than at the St. I, excepting in January when it was more numerous at the St. I. At this time a higher sa1inity was
registered at a11 stations. The presence of
O.
ov~ at the St. I coincided a1ways with higher sa1inity.O.
ov~ occurred in a range of sa1inities from 12.67 to 27.16%0. It was most fre-quent in waters of sa1inity a~out20.00%0 (Fig. 4). The naup1ii of
O.
ov~ occurred in a narrower range of sa1inity than the adu1ts, from 14.30 to 27.16%0 (Fig. 5).Seasonal variation:
the seasona1distribution on nauplii and adu1ts of
O.
ov~ is shown in Figures 6 and 7.St. I
-
the greater number of adu1tspecimens was registered in January (80,600 specimens per samp1e) when the sa1inity was 16.48%0 and the naup1ii were more abundant in December (68,400
specimens per samp1e) in water of sa-1inity 15.23%0. The occurrence of
O.
ovati~ at the St. I, on1y duringFig.
3.
Tidal level at CananéLa (data furnished by the Physics Section - Instituto Oceano-gráfico - Universidade de são Paulo.the months of Decerober, January and May, is associated with the high tide (Fig. 3). A1though the station time was not registered in January, the samp1es were taken during high tide based on the sa1inity data.
St. II
-
the maximum number ofadu1ts and naup1ii per samp1e was 675,000 and 296,800 respective1y in February when the sa1inity
regis-tered was 20.82%0.
St . III
-
nauplii and adu1 ts weremost abundant in January. A number of 45,600 adu1ts and 39,000 naup1ii per samp1e was recorded in waters of sa1inity 20.52% 0 .
This species was more numerous during the Summer months which is characteristic of the ho1op1ankton of the Cananeia region. The higher number of
O.
ov~, at the three stations, occurred in February, one of the warmest months (27.30 0C). Excepting August, adu1ts and juve-ni1es occurred a11 the year round.Ovigerous fema1es were recorded during the who1e period of study, but the higher numbers occurred during the summertime (Fig. 12). This finding suggests that there is a greater intensity of breeding in the warmer season. The greater oc-currence of naup1ii, in February, just fo11ows the maximum of oviger-ous fema1es. It was not possib1e to estab1ish the number of gener-ations, nor the span of 1ife of the species in this region, as a11
stages of deve10pment occurred con-tinuous1y throughout the period of study.
3
4
Bolm Inst. oceanogr., S Paulo, 28(2), 1979o 1- 1.000
O 1.001 _ 10.000
LJ
10.001 _ 100.00031
O
+ 100.00030
29
2e 27
2e
o
~
25
UJ a:: :::J
.... 24
~
a::
UJ 23
!lo
~
UJ 22
....
2 1
20
111
4 5 e 7 8 9 10 12 13 14 15 UI 17 le 19 20 21 22 2S 24 26 28 17 28 SALlNITY (% o)
Fi g.
4.
T.
S. P. diagram-
adult specimens ofOilhona.
ova1..ÁÃ . NUMBER OF SPECIMENSO 1 - 1 .000
32
O 1.001 10 .000
3 1
O
10 . 001 - 100 .00030
O
+ 1000000
29
0
28
27
88
~
~ 26
.
.
8
08
8
w 2 5
a:: :::J
.... 2 4
~
c::
8
G
GJ
w
GJ
Cl- 23 ::e
w
c::J
.... 22
8
2 1
:!J
.
G
8
20
8
19
3 4 5 e 8 9 10 11 12 13 14 15 16 17 18 111 20 21 22 23 24 25 26 27 28 SALlNITY (%0)
Fi g.
5. T.
S. P. d iagram-
naupl i i ofOilhona.
ova1..ÁÃ.o f male s and females of
O.
ova1..ÁÃ. Females 2.2. O~o»aot{gohali»a
FONSECA
&
PRADO:
Olthona:
Cananéia
5
o.
ov~ (Tab. I). A total of 633,600 adu1ts and 6691°00 naup1ii were recordedin all samp1es.
HorizontaZ distribution:
the specieswas recorded in a range of sa1inity from 3.15 to 22.32% 0 • The higher number
(more than 1,000 specimens/samp1e)was found in water with sa1inity from 3.15
Table 1- Number of specimens of
O-i..:thona oUgohaLLna.
and
O. ovai.Á.Á
collected in
ea~hstation (June 1973 to May 1974)
Specles
O.<..tho .. ia o.t.<.goha.t.<.na o.<..tho na ova.tü Specles o.<..thol'la o.t.<.goha.t'<'l'Ia o.<..thol'la ova.t.<.I> Data Male Female
Naupllus I
Naupllus 11 Naupllus 111 Naupllus IV Naupllus V Naupll us VI
Mal~
Female
Naupll us I
Na u p I I u 5 I I Naupllus 111 Naupllus. IV Naupll us V ·N li u P I I u s V I
3.400
20.000
200 20 O
4.60 O
7.20 O
8.600
Copepodltes* 18.400
Date
Male Female
Naupllus I Naupllus 11 Naupllus 111 Naupllu 's IV Naupllus V Naupll us VI Male Fema I e
Naupll us I Naupllus 11 Naupllus 111 Naupllus IV Naupll us V Naupllus VI
Copepodltes*
Ju ne
11 111
2.800 400
26.400 15.000
101.400 87 . 400
(1.800) (400)
2.400 24.600 41 .20 O
32.800
800
I .800 4.800 4.000
88.800 89.000
Septemtrer
11
200 4.600
400
I .000 2.400 ' 2.600 4.800
I .000
17.200
111
200
2.400 21 .800 (600) 600
600
I . 400 2.800 5.800 6 . 000
36.600 200 400 200 July 11
41 .200
141 .200 (4 .6 O O)
600 I • O O O 5.400
44.200
111
800
600 103.000
2.200 2.600 200 600 400 800 3.400 115.600
4.20 O
9.200
55.600
( I • O O O) 200 400 400
I .80 O
2.20 O
7.600
315.600 49.600
October
11 111
600 6.200
19.400 (600)
200 3.000
200
200
I . 80 O 2.600 4.000
2.200 2 . 000 12.400
600 I .600 1 .600
(800)
20 O
200
35.400 28.000 2 . 200
August
11
November
11
37.400 114.00 O
(I I .400)
200
I .20 O
6.600 9.600 13.600 6 . 200
111
111
14 . O O O 70 . 600 (4.400 )
800
3 . O O O
3 . 400
6
B o 1 m I n 5 t. O c ea nc ..;.:-...;.' ~' -..,;;.5...;..;P a~u~1;..:o:;.,i, !...:2:.:8...:.(.::2.:..) .l.' ....:...:1 9~7"-::9:::... Species o,U:ho na Data ~ation Develop-~menta I stage ~
Male Female
Nauplius I Nauplius 11
ot~go hal~na Naupl ius I I I
Naupl ius IV Naupliu sV Nauplius VI
O~.thona ovatü Species o~ .thona Male Female
Nauplius I Naupl ius 11 Nauplius 111 Naupl ius IV Naupl i us V Naupl ius VI
Copepodites*
Data
Male Female
Nauplius I Naupl ius 11
ot~go hat~na Nauplius I I I
Naupl ius IV NaupliusV Naupl ius VI
o.i..tho na ovatü,
Male Female
Nauplius Naupllus 11 Nauplius I11 NdUPlius IV Nauplius V Nauplius VI
Copepodites*
( ) - Ovigerous females
4.000
I .000
I .800 3.000
December
11 I I I
200
400 4.800 ( I . O O O)
400 I .000
January
11 111
(400)
800 2.4 O O 3.600
I .4 O O
800 10.600 600 20.800 9 . 800 13.bOO
15 .00067.80 0 6.200 58 . 400 33 . 400 152.600
(5.40 0) (400) (1.400) (2 . 400) (8.400)
1.6002 . 000
10.800 10 . 600
26.000 19.000
30.000 26.600
200 400 800
3 . O O O
6 . 600 13.20 O 9 . 400
126.000 204.000 10.400 130 . 400
March
11
200 19 . BOo
1.200 134.800 (6 . O O O)
2. O O O 5.60 O
7 . O O O 59 . 000
5 . 800 90 . 000
4,600 94 . 600
4 . 200 111
600 5 . 600
800 4 . 400 16.800
I I .800
19.400 705 . 20024 . 400
2,20 O
36 . 800
(.1 .4 O O)
6. O O O 1i .600 12 . O O O 3 . 600
42.600
800 I .800 I . 20 O
53.800
Ap r i I
11
17 . 400 82.400 (7 . 600)
200 1 . 400
2 . O O O
600
I .20 O 7 • O O O
15.20 O 15 .6 00
205.400
111
I .6 O O
9.000 400
81.80018,000
( I . 40 O)
4 .0 00 3 .200
7 . 800 9.000
II.ÕOO 11.600
9.000 4 .600
41.600 215 .0 007 5.000
cont.
February
11 111
I . O O O
205.400 467.000 (2.600 )
3 . O O O 55.000 127.800 I I I .000
2.107.200 May 11 35,400 60.400 (200) 9.000 19.400 17.200 72,600
10 . O O O 24.800
(I .400)
200 400
I .4 O O
800
57.800
111
8,60 O 35 , 400
1 • O O O 4.20 O
I • O O O
53,400
*
- Copepodites were not counted separately by stage of development because of difficulties in rapid identificationto 16.48%0 (Fig. 8). The naup1ii of
O.
oligo~na occurred 1n sa1inities from 3.15 to 16.48%0 (Fig. 9). The greater abundance occurred 1n sa1inities around 11.00° /00.SeasonaZ variation: Figures 10 and 11
show respective1y the seasona1 variation of adu1ts and naup1ii of O.
oligohalina.
St. I - the higher number of O.oligo-halina
was registered in Apri1 (40,400FONSECA
&
PRADO:OLtho»a:
Cananéia1. 000 . 000
'00 .000 .00. 000
•
i 100. 000 a 10 . 000e
00 . ' "I
40.000i
10.000i
I:,,:
' . 000
L ...
1. 000. - "ATlCMI I
['1 - .,... I
• - ,,.TIC* •
~
I
7'ii"i'~ 7IIi'í'Ii''TI'i''''''''''''I'I'ii'~í'i'i''I7i''T7i' í7i'
Fig. 6.
SI
50
251
28
27
:3 2tI
o
t. 2~ (j
L&J
o: 24 :l
I-'"
o: 25 L&J
elo ~ 22
L&J
I-21
20
UI
3
'TA'IOIII
Relative abundance and seasonal of the adult specimens of
Oi-thon.a ovaLú.:,
at the St. I, IIand III (June 1973 to May 1974).
NUM ER OF S
II 1-1.000
O tOOI - 10.000 O 10.001 - 100.000
O
•
100.000[::J
G ~
~
8G
G
4 ~ 11 7
•
•
10 12 15[!J
14
!
!
e
i
.
-i
,i
~
!
'00. 000
.... 006
'0.000
. - "ATIOlf I
0 - nATlO1f "
.- ""TIOIII •
7
'I7íii'~ ~ '177.' TI"'i'~~~~~'"i'i"""ii'f'I"ITi"1
t'A Tl ON
Fig.
7.
Relative abundance and seasonal variation of the naupl i i -Oi-tho»a ova.LL6 .
8
(j
0 0 ~
G I!
G li
I!
I~ 111 17 I' Je 20 21 22 211 24 ZI! zs 27 211
SALlNIT'I' ("lo o )
F i g. 8. T. S. P. di agram -
Oi:tho»a oügohaL[»a.
S t. II - the species was more abundant in Ju1y (187,000 adu1ts and 267,800
naup1ii/samp1e) in water of sa1inity 12.67%0.
St. III - the species was hard1y repre-sented at this s tation and more than 1,000
8
Bolm Inst. oceanogr., S Paulo, 28(2), 197932
31
30
29
28
27
~ 2 6
... 25
a::
::::>
~ 2 4
a::
...
Cl. 23~
w
t- 22
21
20
19
NUMBER OF SPECIMENS
o l - 1.000
O 1.001 10000
O
10.001 100.000O
+ 100 .000C!J
~;J
~
~
::!J
G G
.
.
.
3 4 5 6 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
SA LlN IT Y (%.)
Fig. 9. T. S. P . diagram - nauplii of
OJ..:thoVl.a. oUgohaLtn.a. .
• - IT.TlO,. ,
.00 000 o - ITATlO". •
• - IroloTIOIIII ••
'00 000
10. 000
_] ll_
1-Fig. 10. Relative abundance and seasonal var i at i on of
OLthoVl.a. oUgo haLtn.a.
at the St. I, I I and I II (June
1973
to May1974).
this time.
The characteristic pattern of seasonal fluctuation for O. oUgoh~Vl.a. was not established. The higher abundances are observed to be in July, November and March. The greatest number occurred in July, with
100 .000
.
t
; 100.000 80.000
• - 'TATIQN ,
0- n AT IOM I
• - S'rAT,,* •
IIO,.T",
Fig. 11. Relative abundance and seasonal variation of the naupli i of
OJ..:thoVl.a. oUgohatiVl.a.(June 1973
to May
1974).
a LotaI of 142,000 specimens for alI three stations when the water temperature was 20.80oC.
FONSECA
&
PRADO: O~thona: CananéiaAs with
O.
ov~, it was not possible to establish the number of generations,• - STATlON I
[J _. 'TAnON ,I
• - nAT!ON.
9
nor the span of life of O.
oUgohaUna
"
.000n
'I: I
I'because al1 the deve10pmental stages, i .~
n
I! I!
ai s o adul ts, o ccurred throughou t the year. ~ I;
i
i
! ;
. ,
i I
Sex ratio (Table lI): females specimens ~ .000
were always more abundant than males.
, ,
I,
, ' , I ,
__ .L.L.
! I
___ -L.L
11 _
I1I
Fig. 12 . Relative abundance and seasonal variation of the ovigerous females of O~thona ov~ and
of
O.
oügohaLüta
at the S t. I,II and III (June 1973 to May 1974) .
I ~)OÇ .
.000
~ • 000
~I."- . " ·-t!:iTi I I' U.1 '1'119. --:~:~ II-' ",7 .. ' '":"'Ii' ~;jl '711'7 ~;~ ' I~';" '·'r;-r.,
,U"IOH
o ,
''fAT IO N
Tab 1 e I I - Sex-rat i o of O~thoYla ov~ and of
O.
oügohaLtna
P e r c e n t a 9 e Month
Station I Station I I Station 111
o. ovali~ o . oligohalina o . avalia o . oligohalina o. ovali~ o . oligchalina
Ju ne 14 . 52
July
August
September o
October 45 . 82 O
November 13.63
Oecember 5.06 12.64
J;,nua ry 25.80 6 . 45 21 . 46
February 3 O . 42
March 14 .28
Apr i 1 5 . 44 16.20
May 2.17 36.87
Discussion and Conclusion
Many papers have been published on systematics and ecology of O~hona
ov~ col1ected in Brazi1ian coas tal and brackish waters. Herbst (1955) de-scribed the fema le of this species from the Cananeia region. Bjornberg (1963) mentions the presence of O. ov~~ as
occasiona1 visitors in coas tal waters off South Brazi1. The deve1opmenta1 stages of O. ova~ were described by Bjornberg (1972) from specimens co1-1ected in Santos, Ubatuba and Cananeia (State of são Paulo). Tundisi & Tun-disi (1968) studied the to1erance to sa1inity of O. ov~ in 1aboratory
22. O O
4.16 23.66 22.97
O
12.32
14 . 59 13.98
9.67 15 . 06
8 . 33 7.78 27.62 9.67 9.76 19.54
O
conditions. Turtdisi (1972) studied the distribution and annua1 variation of this species in the estuarine region of Cananeia. The male of O. ov~ was described by Santos (1970) from Itapagipe Bay (Bahia State). The author found a second species of
o~thona in Cananeia waters and based on Gonzã1ez & Bowman (1965) identified
i-10
Bolm Inst. oceanogr ., S Paulo, 28(2), 1979
dentified by Santos (1970) was new to science and described it as O~hona
oügohaiJ..na.
For the first time the presence of
O.
ovaiM
and O.oügohaiJ..na.
in ariver of this region is registered.
Horizontal distribution
ofO~honaovaiM and
O.
oügohaiJ..na
Sa1inity is an irnportant factor controlling the distribution of organ-isms in estuari es (Kilby, 1955; Gunter, 1956; Kinne, 1971; Vernberg
&
Vernberg, 1972). Many papers on 1aboratorystudies of the sa1inity to1erance by marine copepods are avai1ab1e in the 1iterature (Marsha11
et
alt.,
1935; Barnes, 1953; Ranade, 1957; Hooper,1960; Lance, 1962, 1963, 1964; Battag1ia &
Bryan, 1964; Tundisi
&
Tundisi, 1968; Moreira&
Yamashita, 1973). The distri-bution ofEunytemona
an6~ni6 in reJation to sa1inity was studied by Vaupe1-K1ein &Weber (1975) based on fie1d and
1abora-tory observations. .
Figures 4 and 5 show that O. ov~
prefers higher sa1inities than O.
oügo-haiJ..na
and that both species can coexistin a sma11 range of sa1inity (12.67 to 16.48% 0). The coexistence of both species beyond these 1imits was on1y oc-casiona11y registered. Intermediate forms between the two species were not recorded. Bay1y (1965) found that two c10se1y re1ated species of P~eudo
diaptomUó
are ab1e to coexist in .Brisbane River but one of the spec~es
is a1ways dominant. O~hona bnev~
eonni6
and O.nana
occur together inA11igator Harbor but the first is dominant (Grice, 1956). The dominance of one of the two species was a1so verified in the range of sa1inity in which they occur toget"er in Cananeia.
Tundisi (1972) estab1ished the 1imits of salinity for adults and copepodits of
O.
ovaiM
as 1.00 - 34.06° /00 but higherabundances (+ 10,000 specimens) were more frequent from 9.05 to 30.68%0. Tundi-si
&
Tundisi (1968) found that under 1aboratory conditions the superior and inferior 1etha1 sa1inities arere-spective1y 6.30° /00 and 26.00 to 30.00° /00. The first va1ue correspo~ds to the 10w
sa1inity in which
O.
oügohaiJ..na
can be found and 34.06%0 to the higher sa-1inity, thus Tundisi&
Tundisi (1968~and Tundisi (1972) probab1y worked w~th
a mixed popu1ation of the two species instead of experimenting on
O.
ov~on1y, and their data therefore can not
be compared to ours.
Santos (1970) ana1ized various p1ankton samp1es c011ected in coasta1 water from the Brazi1ian coast and found
O.
ovaiM
from Ponta do Seixas (PA) inthe North to paranaguâ (PR) in the South in sa1inities varying from 11.91 to 35.89%0'
The limits of salinities registered for
O.
ovaiM
by Santos(op.
c~t.) in Cananeiaagree with the present data. On the other hand, the data recorded for O. hebM (=
0-ügohaiJ..na)
in Todos os Santos Bay bySantos
(op.
~.) ar~ opposite to those found in Cananeia. Possib1y the O~onac011ected at Todos os Santos Bay is not
O.
oügohaiJ..na.
The presence of O.
ovaiM
at Station I~s associatec with the incomming tide.
O.
oügohaiJ..na
was on1y occasiona11yfound at St. 111, and in sma11 number, suggesting that St. 111 is an expatri-ation area for the species (Yashnov, 1966) .
The inferior limi t of salini ty in which naup1ii of O.
ovaiM
occurred is higher than that registered for adu1ts of the same species. The 1imits of sa1inity for naup1ii of O.ovaiM
registered by Tundisi (1972) are much wider than the present records and must be based on experiments with mixed specimens of O.úvaiM
and O.oügohaiJ..na.
O.
oügohaiJ..na
naup1ii were mostabundant in water of sa1inity varying from 3.15 to 16.48%0' The sa1inity 1imit for naup1ii and adu1ts are co-incident.
The resu1ts obtained show that the samp1ing method was not good for the naup1ii because the first stages
suffered more se1ection than the 01del stages by escapement through the net meshes. Stage I, 11 and 111 were scarce1y represented in the samp1es. Obviously a natural population presents higher densities of younger than 01der stages due to morta1ity.
The number of adu1ts of O. ov~ was twice the number of naup1ii and the a-dults of O.
oügohaiJ..na
were slight1y 1ess abundant than their nauplii. These records suggest that there is a migration of adult specimens from coasta1 waters into the estuary since the number of samp1es in the three Stations were the same. O.oügohaiJ..na
is an endemic species of the estuary.FONSECA
&
PRADO:
O~thona:Cananêia
the species composition changed along the De1aware River characterizing the inner reaches, a medium portion and the en-tr ance of the es~uary.
It is impo r tant t o rema rk t hat the species considered i n this paper ar e congeneric and very closely r e l ated. The occurrence of congene r ic and
com-petitor species is very often found in estuaries. Lance (1963) showed experi-mentally thatclosely related planktonic species can have different abilities to survive in low salinity waters. Ae~
t On6a
has a higher to lerance to dilutedwater than
A.
b~6llo~a and this species is more tolerant thanA.
~ca u dat a .Jeffries (1967) records con generic associations of some species of Copepoda and Cladocera in a region between New England and Virginia. He registers the coexistence of
o;;thona
b~ev~co~~ and O. ~~~ and classified them as estu-arine-marine and euryhaline-marine species respectively. According to Jeffries' classification,O.
oligohalina
is a true estuarine species and
O.
ov~ is an estuarine-marine species.Jeffries
(op.
~.) made some refer-ences on the speciation of the genus E~ytemo~ which appears to have pene-trated the osmo-regulatory barrier inde-pendently, forming a chain of species with overlapping distribution along thesalt gradient. Each member of the chain probably had a better ability to toler-ate lower salinity. A continuous chain of species extending from salty to fresh water can be the result of an
intra-generic competition.
The .presence of two very closely related species of
Oithona,
with differ-ent limits of salinity tol~rance, in anarrow section of the estuary, can be explained by the mechanisms described by Jeffries
(op.
~.) and indicates that the genus O~hona has suffered speci-ation into the estuary.The intraspecific conflict plays an important role in the distribution of certain species. Hodgkin & Rippingale (1971) examined this problem in Swan River (Australia). The great abundance of copepods that prey upon nauplii can restrict the distribution of other
euryha~ine species of the same group,
to an 1nner part of the estuary where the environmental conditions are adverse to the predator species. The relation between O. OVctÜ4 and
kaJdÁ...a
~jebo~g~ in Cananeia seems to be analogous to that described by Hodgkin
&
Rippingale11
(op.
~.). Grea t numbers of nauplii,copepodi tes and adul ts of
A.
~j ebo~g~ were registere d in Sts. 11 and 111 in Septemb e r , while O. OVctÜ4 was less numerous .A.
lilije bo~g~ was still abundant and the quantity of O. OVctÜ4 strongly decreased in October. This inverse , relation ofA.
~jebo~g~ andO. OVctÜ4 can probably be ascribed to
predation effe cts since the environmental conditions were good to the occurrence of O. OVctÜ4.
Oxygen:
the valulO's of dissolved oxygenfound in the Taquari River are higher than those recorded by Prado (1972) in depth, by Tundisi (1972) at the surface and by Kutner (1972) at the surface in a tidal creek of the Cananeia region. According to Kato (1966) the remarkably low quantity.of oxygen in the tidal
creek water is caused by a characteristic state of reduction in the upper reaches of the "river" and indicates a correlation with the quantity of reduced matter in
the water. Taquari River has a differ-ent condition from that found in the tidal creek because low values of
dissolved oxygen were never found. Thus dissolved oxygen is not a controlling factor of the distribution of O~hona in this section of the Cananeia estuarine region.
Temperature: a slight increasing gradient of temperature was recorded from S t. I to S t. III throughout the period of sampling. However this small difference ot temperature was not sufficient to af-fect the horizontal distribution of the genus O~hona in this area. The gradient of temperature neve r goes beyond the limit of tolerance of both species.
SeasonaZ variat ion
Estuaries have a marked seasonal vari-ation (Riley, 1967). The Summer and Winter are well characterized in
Cana-neia by the quantitative composition of p1ankton (Tundisi, 1972).
Jeffries (1962a,b) showed that one species can seasonally take the place of another closely related to it in an estuary of the east coast of America. He supposed that these congeneric species were in competition and that environmental
conditions (salinity and temperature
combinations) favor one of the two species as in the case of the two
Oithona
from Cananeia, but these do not show the pattern of seasonal substitution.O. OVctÜ4 and O.
otigohatina
were12
Bolm Inst. oceanogr., S Paulo, 28(2), 1979
excepting in August when only animal skeletons and diatoms valves were present. The higher occurrence of (1.
oUgoha.üvr.a
was recorded in July when temperature was only a little higher than August and salinity was within the limit of tolerance of bothspecies. This lack of plankton must be due to some other factor than temperature and salinity.
Three peaks in the abundance of
O.
oU-gohalina
were registered throughout theyear (July, November and March) alI at St. 11. Ovigerous females and nauplii were most abundant in these same samples. Possibly the salinities found at St. 11 during these months are elose to the opti-mum for O.
oUgohalina.
A more regular annual distribution was observed in O. ov~. A higher number of ovigerous females and nauplii occurred during the surnrnertime. It seems that the reproductive activity is more intense in this period. A significative number of ovigerous females (more than 1,000 speci-mens/sample) begins in November and reaches its maximum in January.
No data exists on seasonal variation of
o.
ov~ and O.oUgohalina
in otherto Rayrnont
&
Miller (1962), Deevey (1948) found a main period of reproduction of O.bnevieonni6
in Tisbury Great Pond besidessmaller broods. Faber (1966) registered a great number of O.
b/l.e.vieonni6
inNarraganset Bay in the Surnrner. According to Sazhina (1971) O.
nana
has seven annu-aI maxima and possibly an eighth de-pending on the temperature. In lower temperatures the development takes a longer time. It is difficult to es-tablish the number of generations ofOithona
in Cananéia because therepro-duction is continuous throughout the year.
Cananéia region shows the small annu-aI variation of temperature typical of a tropical region. This variation is not wide enough to control the distri-bution of planktonic animaIs or to de-termine a marked life cycle as it does in colder regions. Here, the salinity and predation seem to play a more important role on the occurrence and distribution of planktonic animaIs.
Sex-ratio
The present records confirm previous areas. O. ov~ showed an analogous pattern ones from this area (Teixeira e;t
ai.,
of seasonal variation to that of the O. 1965; Tundisi, 1972). Adult femalesbnevieonni6
found by Grice (1956) in of bothoithona
are always morenumer-I ' H b O
nana
1 has one ous than adult males (Tab. 11). ItA ~gator ar o r . . a so ' h " n +
1 ' (G' 1956' E terly must be po~nted out t at Te~xe~ra
I::A-annua max~murn r~ce, , s , _o ( ~:+) d T d' , (on
eit)
1928' Woodmansee 1958' Hirota, 1962) al-!.U.·
op.
l.A.A..:
an un ~~~ I V ' , •" ' , ' , ' r e f e r to a m~xed populat~on of O~hovr.a though ~t occurred ~n d~fferent per~ods _o:.
O
o:h-O:
, h 1 ov~ plus . o~go ~vr.a.
of the year accord~ng to t e p ace con- , ,
, d O i :, •• :0:. h aI a ~ The l~terature conta~ns many data on
s ~dere. • /.,).{.1rKAA.oO as one annu m x .... - ,
' h ' f ' d (L' 1967) sex-rat~o of copepods (Carnpbell, 1934; mum ~n t e Norweg~an Jor s ~e, . ,
O
U hali
h 1 d' rete Medn~kov, 1962). The males of many" O h
go
hna
t thas severao~s~
n;l/I; _ species have a shorter span of life thanmax~ma t roug ou e year as . ~/V~'~ ,
+~:, (G d 1953) d O i : ... :0:, the females. The sex-rat~o of one
nO-6vlA./.J
un ersen a n . / . , ) . { . 1 r K A A . o O , ,e;t
ai
'1973) A k' spec~es can vary depend~ng on the place(Frolande~ .~, . pea ~n where they live (Bogorov, 1939 after
reproduct~vehact~v~ty was not observed Mednikov, 1962). Mednikov (1962) studied throughout t e seasons. the sex-ratio of three pelagic groups:
Tundisi (1972) estab1ished two estuarine, superficial oceanic and bathy-patterns of seasonal cycles for copepods pelagic. He carne to the conclusion that from Cananéia region. The first includes the percentage of males increases from
O.
ovaiM,
Ae~ ~jebongi andPana-
bathypelagic to the estuarine. Heob-eaianM
efW...6-6~o-6bú.6 which have a period ta.ined 50% of males in epiplanktonicof more intense reproduction. This is species such as
Aeaktia tumida
Willey typical of tropical estuarine species of and Centnopag~ memUhft~ehi Willey. The planktonic copepods. 'l.he secor:d inclu~es p<;pulations ofOithona
stu~ied in Cana-P-6eudo~aptomMaeutu-6,
Eutenp~vr.aaeuti-
neia never showed such a h~gh percentagenno n-6
andLabidoeena
M-uv~ which of males.have several discrete maxima. The present
findings confirm the posi tion of O. ov~
Cone 1 us
ions
and addO. oUgohativr.a
to the secondgroup.
There are very few records on seasonal variation of
oithovr.a
nauplii. AccordingFONSECA
&
PRADO:
O~hona:Cananéia
2
3
-According to Jeffries' classification (1967), O.ov~ is estuarine-marine and O. o~gohalina is true-estuarine. The presence of two closely related congeneric species, the distribution of which partially overlaps in a
narrow range of salinity suggests that 3
-13
sofreu especiação dentro do estuário. A salinidade tem um papel importante na ocorrência e dis tribuição des tas especies desde que a temperatura nun-ca alnun-cançou valores limitantes para estas espécies.
the genus O~hona has suffered speci- Acknowledgements
4
-ation within the estuary.
O.
ov~ was more abundant thanO.
o~gohalina. Both species occurred throughout the year.
5 - O. ov~ has an intense reproductive period in the Summer and others less intense throughout the year. O.o~go
halina
showed only three discretemaxima alI year round.
6 - The co-existence, throughout the year of a population composed by nauplii and adults indicates
continuous breeding of these species in the area with periods of greater reproductive activity. 7 - It was not possible to determine the
number of generations.
8 - The salini ty plays an important role in the occurrence and dis tributions of both species since the temperature never reached limiting values for both species.
9 - The percentage of adult females of both species was always higher than adult males.
Resumo
Alguns aspectos ecológicos de duas espé-cies congenéricas de copépodos ciclopói-des, O~hona ov~ e O. o~goh~na são estudados pela primeira vez na região estuarina de Cananéia. A distribuição espacial e variação sazonal destas espé-cies são estudadas em relação
ã
salini-dade, temperatura e oxigênio dissolvido. Os resultados obtidos são comparados com trabalhos preliminares feitos nesta área e estudos similares em outras regiões. As principais conclusões obtidas são as seguintes:1 - O. ov~ ocorre em uma faixa de sa-linfdade de 12,67 a 27,16%0 e O. o~goh~na em 3,45 a 16,48%0' 2 - A presença de duas espécies
congené-ricas estreitamente relacionadas cu-ja distribuição se recobre parcial-mente em uma estreita faixa de sali-nidade sugere que o gênero o~hona
We wish to express our thanks to Dra. Tagea K. S. Bjornberg for her criticaI reading of
the manuscript and valuable remarks.
Bibl iography
BARNES, H. 1953. The effect of lowered salinity on some barnacle nauplii. J. Anim. Ecol., 22:328-330.
BATTAGLIA, B.
&
BRYAN, G. W. 1964. Some aspects of ionic and osmotic regulation in T~be (Copepoda, Harpacticoida) in relation to polymorphism and geographi-cal distribution. J. mar. biol. Ass. U. K., 44:1-15.BAYLY, l. A. E. 1965. Ecological studies on the planktonic Copepoda of the
Brisbane River estuary with special reference to
Gfadio6enen6
pectinatU6(Brady) (Calanoida). Aust. J. mar. Freshwat. Res., 16(3) :315-350.
BJ~RNBERG, T. K. S. 1963. On the marine free-living copepods off Brazil. Bolm Inst. oceanogr., S Paulo, 13(1) :1-142. 1972. Developmental stages of some tropical and subtropical planktonic marine copepods. Stud. Fauna Curaçao, 40:1-185.
CAMPBELL, M. H. 1934. The life history and postembryonic development of the copepods, CafanU6 tOn6U6 and Euc.haeta
japonic.a.
J. biol. Bd Can., 1:1-65.CRONIN, L. E.; DAIBER, J. C. & HULBERT, E. M.
1962. Quantitative seasonal aspects of zooplankton in the Delaware River estu-ary. Chesapeake Sci., 3:63-93.
ESTERLY, C. O. 1928. The periodic oc-currence of Copepoda in the marine plankton of two successive years at La Jolla, California. BulI. Scripps Instn Oceanogr. tech. Ser., 1(14) :247-345.
FABER, D. J. 1966. Seasonal occurrence and abundance of freeswimming copepod nauplii in Narragansett Bay. J. Fish. Res. Bd Can., 23(3):415-422.
_1_4 __________________________________ ~B~o~l~m~l~n~s~t~.~o~c~e~a~n~c ~' r., S Paulo, 28(2) ~l~
Cyclopoida). Bo1m Zool., Univ. S Pau-lo, 1:205·-238.
FONSECA, V. L. 1976. Copêpodos do gênero O~hona da região de Cananêia (Lat.
25°07'S; Long. 47°56'W). Dissertação de mestrado. Universidade de são Paulo, Instituto Oceanografico, 68p.
- - - & BJÔRNBERG, T. K. S. 1976.
Oithona oligohalina
sp. n. de Cananêia(Est. de são Paulo) e considerações sobre O. ov~ Herbst (Copepoda,
Cyc10poida). Anais Acad. bras. Ciênc., 47: 127-131.
FRAGA, F. 1972. E1 agua marina. In:
Castellví, J.
etal.
-
Ecologia marina. Caracas, Editorià1 Dossat, p.67-99. FRASER, J. H. ed. 1968. Standardizationof zoop1ankton samp1ing methods at sea. Larger mesozoop1ankton. Report of ICES/SCOR/UNESCO, Working Party n9 3. Monogr. oceanogr. Methodo10gy, (2) :160-163.
FROLANDER, H. F.; MILLER, C. B.; FLYNN, M. J.; MYERS, S. C. & ZIMMERMANN, S. T. 1973. Seasona1 cyc1es of abundance in zoo-p1ankton popu1ations of Yaquina Bay, Oregon. Mar. Bio1., 21(4) :277-288. GONZÁLEZ, J. G. & BOWMAN, T. E. 1965.
P1anktonic copepods from Bahia Fos-forescente, Puerto Rico, and adjacent waters. Proc. U. S. natn. Mus.,
117(3513) :241-304.
GRICE,G.D. 1956. A qualitative quantitative seasona1 study of Copepoda of A11igator Harbor. oceanogr. Inst. FIa St. Univ., 37-76.
and the Contr.
(68) : 1960. Copeprids of the genus O~hon~ from the Gu1f of Mexico. BulI. mar. Sci. Gu1f Caribb., 10(4):485-490.
GUNDERSEN, K. R. 1953. Zoop1ankton in-vestigations in some fjords in Western Norway during 1950-1951. FiskDir. Skr., Ser. Havunders., 10(6):1-54.
Gu~TER, G. 1956. Some re1ations of
fauna1 distributions to sa1inity in estuarine waters. Eco10gy, 37:616-619. HERBST, H. V. 1955. Cyc1opoida
Gnathos-torna (Crustacea, Copepoda) der
Brasi1ianischen At1antikküste. Kie1er Meeresforsch., 11(2) :214-229.
HIROTA, R. 1962. Species composition and seasona1 changes of copepod fauna in the vicinity of Mukaishima. J. oceanogr. Soc. Japan, 18(1) :35-40.
HODGKIN, E. P. & RIPPINGALE, R. J. 1971 . Interspecies conf1ict in estuarine copepods. Limno1. Oceanogr., 16(3): 573-576.
HOOPER, A. F. 1960. The resistance of marine zoop1ankton of the Caribbean and South At1antic to changes in sa-1inity. Limno1. Oceanogr., 5:43-47. JEFFRIES, H. P. 1962a. S alini ty-space
distribution of the estuarine cope-pod genus
Eunytemoha.
Int. Revue ges. Hydrobio1. Hydrogr., 47:291-300.1962b. Succession of two ka.Jr;Ü.a species in estuaries. Limno1. Oceanogr., 7:354-364.
1967. Saturation of estuarine zoop1ankton by congeneric associates. In: Lauff, G. H., ed. -Estuaries. Washington, Am. Ass. Adv. Sci., ' p.500-508.
KATO, K. 1966. Geochemica1 3tudies on the mangrove region of Cananêia, Brazi1. I. Tida1 variations of water properties. Bo1m Inst. oceanogr.,
S Paulo, 15(1) :13-20.
KILBY, J. D. 1955. The fishes of two gu1f coasta1 marsh areas of Florida. Tu1ane Stud. Zool., 2(8):175-247. KINNE, O. 1967. Physio10gy of estuarine
organisms with specia1 reference to sa1inity and temperature: general aspects. In: Lauff, G. H., ed. -Estuaries. Washington, Am. Ass. Adv. Sci., p .525-540.
1971. Marine eco10gy: a comprehensive, integrated treatise on 1ife in oceans and coasta1 waters. London, Wi1ey-Interscience, vo1. 1, part 2.
KUTNER, M. B. B. 1972. Variação es tacio-nal e distribuição do fitop1âncton na região de Cananêia. Tese de doutorado. Universidade de são Paulo, Instituto de Biociênciês, 104p.
LANCE, J. 1962. Effects of water of reduced sa1inity on the vertical m~
gration of zoop1ankton. J. mar. bio1. Ass. U. K., 42:131-154.
1963. The sa1inity to1erances of some estuarine p1anktonic copepods. Limno1. Oceanogr., 8:440-449.
FONSECA
&
PRADO:
Oithona:
Cananéia
LIE, U. 1967. The natural history of the Hardangerfjord. 8. Quantity and composition of the zoop1ankton, Sep-tember 1955 - SepSep-tember 1956. Sarsia,
30:49-64.
.
MARSHALL, S. M.; NICHOLLS , A. G. & OR.. ~ , A. P .
1935. On the bio10gy of
CalanUó
6).11-m~ehieUó. VI . Oxygen consumption i n re1ati on to environmenta1 conditions. J. mar. b io l . Ass. U. K., 20(1):1-28.
MEDNIKOV, B. M. 196 2 . On the sex-ratio in deep sea Calano ida. Crustaceana, 3:105-109.
MOREIRA, G. S.
&
YAMASHITA, C. 1973. Experimental studies on phys iologi-cal and behavioura1 response mecha-nisms of the p1anktonic copepodEu-t~pina
aeuti6nonó
(Dana) to various sa1inities. Bo1m Zool. Bio1. mar., n.s., (30):233-244.PRADO, M. S. de A. 1972. Mysidacea (Crustacea) da região 1agunar de Ca-naneia. Tese de doutorado. Universi-dade de são Paulo, Instituto de Bio-ciências, 86p + 68 figs.
Primary producti on, zoop1ankton standing-stock and some environ-mental fac to r s . Int. Revue ges. Hydrob io1. Hydrogr., 54(2):289-301. TUNDISI, J. 1970 . O p1âncton
estua-rino. Contrções Inst. oceanogr. Univ. S Paulo, ser. Oceano bio1.,
(19):1-22 .
& TUNDISI,T.M. 1968.
P1ankton studies in a mangrove en-vironment. V. Sa1inity to1erances of some p1anktonic crustaceans.
15
Bo1m Inst. oceanogr., S Paulo, 17(1): 57-65.
TUNDISI, T. M. 19 72. Aspectos ecológi-cos do zoop1âncton da região 1agunar de Cananeia com especial referência aos Copepoda (Crustacea). Tese de doutorado. Universidade de são Paul o, Instituto de Biociências, 191p.
VAUPEL-KLEIN, J. C. von & WEBER, R. E. 1975. Distribution of
Eunytemona
a66i~ (Copepoda: Ca1anoida) in re1ation to sa1inity: fie1d and 1aboratory observations. Neth. J. Sea Res., 9(3-4):297-310.
RANADE, M. R. 1957. Observations on the
resistance of
TigniopUó
6ulvlL6 (Fisher) VERNBE~G, W. B. & VERN~ERG, F. J. 1~72.to changes in temperature and sa1ini- Env~ronmenta1 phys~ology of m a r~ne
ty. J. mar. bio1. Ass. U. K., 36: animaIs. New York, Springer-Wer1ag,
115-119. 346p.
RAYMONT, J. E. G. & MILLER, R. S. 1962. Production of marine zoop1ankton with ferti1ization in an enc10sed body of sea water. Int. Revue ges. Hydrobio1. Hydrogr., 47(2):169-209.
RILEY, G. A. 1967. The p1ankton of estuaries.
In:
Lauff, G. H., ed. Estuaries. Washington, Am. Ass. Adv. Sci., p.316-326.SANTOS, J. J. 1970. P1âncton da Baía de Todos os Santos, com especial referên-cia aos copepodos. Dissertação de Mestrado. Universidade de são Paulo
,
Instituto de Biociências, 49p.SHAZINA, L. I. 1971. Annua1 cyc1e of deve10pment of mass Copepoda in the B1ack Sea. Gidrobiol. Zh., 7(5): 38-46. (em russo).
TEIXEIRA, C.; TUNDISI, J. & KUTNER, M. B.
1965. Plankton studies in a mangrove environmeLt. 11. The standing-stock and some eco1ogical factors. Bolm Inst. oceanogr., S Paulo, 14:13-41.
& SANTORO YCAZA, J. 1969. P1ankton studies in a mangrove environment. VI.
WOODMANSEE,R.A. 1958. The seasona1 distribution of the zoop1ankton off Chicken Key in Biscayne Bay , F)orida. Eco1ogy, 39:247-262.
YASHNOV, V. A. 1966. Water masses and p 1ank ton. 4.