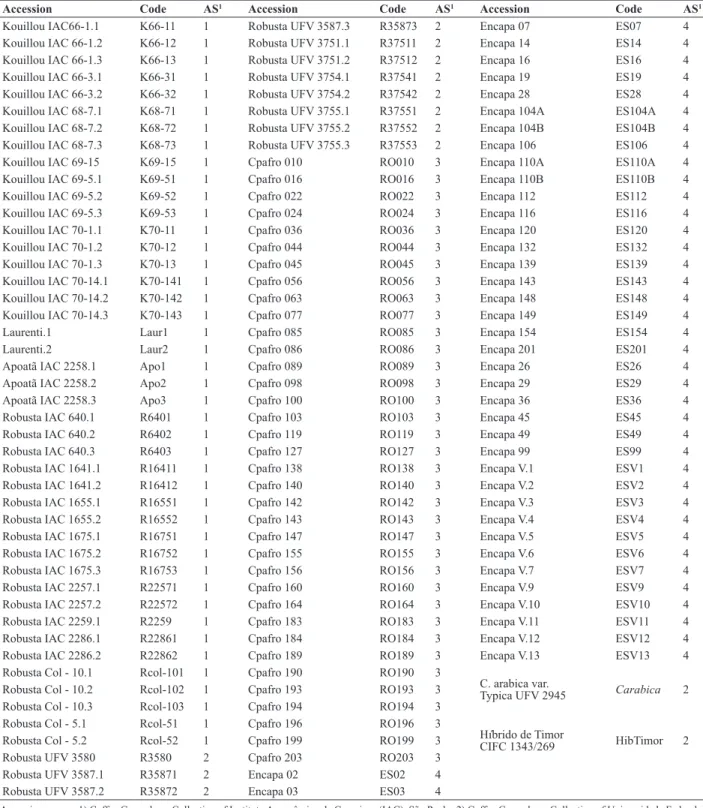
Molecular diversity in Coffea canephora germplasm conserved and cultivated in Brazil
Texto
Imagem

Documentos relacionados
A capacidade para exercer influência depende, em grande parte, da eficácia dos processos de comunicação.. Emissor Receptor Receptor Emissor Código O processo de comunicação
financeiras, como ainda por se não ter chegado a concluo entendimento quanto à demolição a Igreja de S. Bento da Ave-Maria, os trabalhos haviam entrado numa fase
Uma, o aumento do grau de intervenção do MCPS na política de habitação, estimulado pelo decreto-lei n.º 39978 de 20 de dezembro de 1954, que atribuía ao ministro das Corporações
• The possibility of calling an early election imply, by itself, that left-wing man- dates are characterised by a higher expected inflation level, and via (8) also a higher
O experimento foi realizado com uma amostra de 43 alunos dos cursos de licenciatura em Engenharia Informática da Universidade Fernando Pessoa, como já referido,
Este trabalho teve como objetivo a elaboração de sabão a partir do óleo de nabo forrageiro e do óleo residual de fritura, visando a preservação do meio
• Levantar dados estatísticos de incêndios em edificações residenciais e em áreas verdes atendidos pelo Corpo de Bombeiros no município de Belém no período
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
