THE POWER OF THE SMALL: THE EXAMPLE OF Paracoccidioides brasiliensis CONIDIA
Texto
Imagem
Documentos relacionados
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
This log must identify the roles of any sub-investigator and the person(s) who will be delegated other study- related tasks; such as CRF/EDC entry. Any changes to
Além disso, o Facebook também disponibiliza várias ferramentas exclusivas como a criação de eventos, de publici- dade, fornece aos seus utilizadores milhares de jogos que podem
Despercebido: não visto, não notado, não observado, ignorado.. Não me passou despercebido
Assim, surge o projeto de pesquisa Uso de Técnicas de Gamificação como Auxílio ao Ensino e Aprendizagem de Lógica de Programação, a fim de desenvolver um ambiente
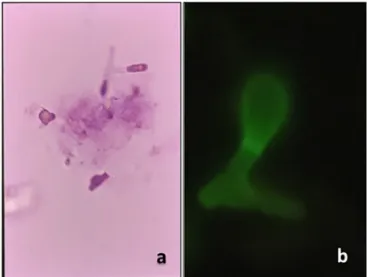
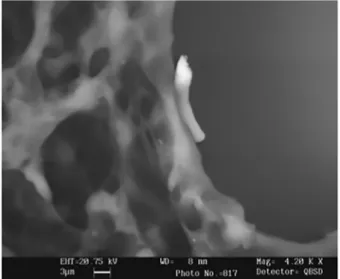
Fig. brasiliensis intracellular transition and CFU of conidia after lysozyme inhibition in normal or TNF-α activated Møs. brasiliensis conidia, and B) yeast cells observed after 96
i) A condutividade da matriz vítrea diminui com o aumento do tempo de tratamento térmico (Fig.. 241 pequena quantidade de cristais existentes na amostra já provoca um efeito
didático e resolva as listas de exercícios (disponíveis no Classroom) referentes às obras de Carlos Drummond de Andrade, João Guimarães Rosa, Machado de Assis,
