Structural and Functional Studies of
H.
seropedicae
RecA Protein
–
Insights into the
Polymerization of RecA Protein as
Nucleoprotein Filament
Wellington C. Leite1
*, Carolina W. Galvão2, Sérgio C. Saab1, Jorge Iulek3, Rafael M. Etto3,
Maria B. R. Steffens4, Sindhu Chitteni-Pattu5, Tyler Stanage5, James L. Keck6, Michael M. Cox5
*
1Department of Physics, Ponta Grossa State University (UEPG), Av. Carlos Cavalcanti, 4748, CEP. 84.030–900, Ponta Grossa, PR, Brazil,2Department of Structural and Molecular Biology and Genetics, Ponta Grossa State University (UEPG), CEP 84030–900, Ponta Grossa, PR, Brazil,3Department of Chemistry, Ponta Grossa State University (UEPG), CEP 84030–900, Ponta Grossa, PR, Brazil, 4Department of Biochemistry and Molecular Biology, Federal University of Parana, CEP 81531–980 Curitiba, Brazil,5Department of Biochemistry, University of Wisconsin–Madison, Madison, WI, 53706–1544, United States of America,6Department of Biomolecular Chemistry, University of Wisconsin School of Medicine and Public Health, Madison, WI, 53706, United States of America
*[email protected](MC);[email protected](WL)
Abstract
The bacterial RecA protein plays a role in the complex system of DNA damage repair. Here, we report the functional and structural characterization of theHerbaspirillum seropedicae
RecA protein (HsRecA). HsRecA protein is more efficient at displacing SSB protein from ssDNA thanEscherichia coliRecA protein. HsRecA also promotes DNA strand exchange more efficiently. The three dimensional structure of HsRecA-ADP/ATP complex has been solved to 1.7Åresolution. HsRecA protein contains a small N-terminal domain, a central core ATPase domain and a large C-terminal domain, that are similar to homologous bacte-rial RecA proteins. Comparative structural analysis showed that the N-terminal polymeriza-tion motif of archaeal and eukaryotic RecA family proteins are also present in bacterial RecAs. Reconstruction of electrostatic potential from the hexameric structure of HsRecA-ADP/ATP revealed a high positive charge along the inner side, where ssDNA is bound inside the filament. The properties of this surface may explain the greater capacity of HsRecA protein to bind ssDNA, forming a contiguous nucleoprotein filament, displace SSB and promote DNA exchange relative to EcRecA. Our functional and structural analyses pro-vide insight into the molecular mechanisms of polymerization of bacterial RecA as a helical nucleoprotein filament.
a11111
OPEN ACCESS
Citation:Leite WC, Galvão CW, Saab SC, Iulek J, Etto RM, Steffens MBR, et al. (2016) Structural and Functional Studies ofH.seropedicaeRecA Protein–
Insights into the Polymerization of RecA Protein as Nucleoprotein Filament. PLoS ONE 11(7): e0159871. doi:10.1371/journal.pone.0159871
Editor:Maria Spies, University of Iowa, UNITED STATES
Received:May 9, 2016
Accepted:July 8, 2016
Published:July 22, 2016
Copyright:© 2016 Leite et al. This is an open access article distributed under the terms of the
Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability Statement:All PDB files are available from thehttp://www.rcsb.org/database (accession number(s) 5JRJ).
Introduction
The bacterial RecA protein plays a role in the complex system of DNA damage repair. RecA protein catalyzes strand exchange reaction between single-strand DNA (ssDNA) and homolo-gous double-strand DNA (dsDNA) molecules, and also induces the expression of DNA repair proteins in response to DNA damage through a regulatory network known as the SOS response [1–8].
TheEscherichia coliRecA protein (EcRecA) monomers bind onto DNA producing a right-handed helical nucleoprotein filament [9,10]. Recently the EcRecA crystal structures of RecA-ssDNA and RecA-dsDNA complexes have been reported [11], confirming that each RecA monomer interacts with three nucleotides of ssDNA or three base pairs of dsDNA. In the ssDNA complex structure the ATP-binding site is located at the subunit-subunit interface. The EcRecA-ssDNA nucleoprotein filament represents the structural intermediate responsible for homology pairing to a donor dsDNA, and the RecA-dsDNA structure is an end product after the strand exchange reaction [11].
The bacterial RecA monomer has three major domains: a small N-Terminal (NTD), a core ATPase domain and a large C-terminal domain (CTD). Members of RecA family, including the RadA protein of Archaea and the Rad51 and Dmc1 proteins of eukaryotes, share a core ATPase domain that contains the nucleotide binding site, the conserved Walker A and B motifs, and the putative DNA binding site(s), designed loops L1and L2[12–18].
The NTD of RecA family also shares an important domain responsible for self-polymeriza-tion of monomers in the presence or absence of DNA. The NTDs of the human Rad51 and the archaeal RadA protein have been both implicated in dsDNA binding [12,19], while RecA pro-tein has an extra CTD for dsDNA binding [20]. The NTD of EcRecA protein structure contains 33 amino acids and assumes a conformation including anα-helix motif and a coil region. The coil region has a shortβ-loop polymerization motif, which interacts with aβ-strand from the
core ATPase domain, and is responsible for polymerization of the helical nucleoprotein fila-ment in the compressed and extended RecA nucleoprotein conformations. Similar to theβ
-loop of RecA protein, RadA/Rad51/Dmc1 proteins have aβ-polymerization motif in the NTD which together with a subunit rotation motif is responsible for structural transitions from an inactive hexameric ring to right and left—handed filaments [12,13,21,22].
The RecA protein can bind both ssDNA and dsDNA, but the nucleation occurs much more rapidly on ssDNA [8]. RecA protein nucleates onto DNA as oligomers of 4–6 subunits, and then the polymerization rapidly extends primarily in the 50to 30DNA direction at rates greater
than 1000 monomers/min at 37°C [23–25]. Two conformations of helical RecA filaments have been structurally characterized. The first is an inactive and compressed form, formed by RecA alone or bound to ADP, either in the absence of DNA or bound to ssDNA or dsDNA. The sec-ond one is an active and extended filament, formed in the presence of DNA and ATP-γ-S or
ATP [24,26–28].
In the present study, we report a functional and structural characterization of a RecA pro-tein fromHerbaspirillum seropedicaeSmR1 (HsRecA), an endophytic bacterium capable of fix-ing nitrogen under ammonium and oxygen limitfix-ing conditions. Analysis of theH.seropedicae
SmR1 genome sequence indicates the absence ofrecF,dinI,dinB,recBCD,sbcA and sbcCD
genes and the presence of a truncatedumuCgene [29,30], suggesting that RecA plays an important and novel role in DNA recombination and repair systems in this bacteria. Using ATPase, strand exchange assays and electron microscopy images, we compared wild type HsRecA and EcRecA protein activities, focusing on HsRecA protein binding onto ssDNA. We also determined the three-dimensional structure of HsRecA protein and explored the molecu-lar mechanisms of polymerization of bacterial RecA as a helical nucleoprotein filament.
Material and Methods
Reagents
Restriction enzymes were purchased from New England Biolabs. Glycerol, Tris buffer were purchased from Fisher. All other reagents were obtained from Sigma unless otherwise described.
Plasmid Construction, Cloning and Overexpression Host
The fragmentNdeI/BamHIcontaining the wild typerecAgene ofH.seropedicaefrom the plas-mid pAETWT-HMK [29] was cloned intoNdeI-BamHIdigested vector pET21a (Novagen). The integrity of the cloned fragment was confirmed by sequencing.E.coliSTL2669, a nucle-ase-deficient strain, was used as host to overexpress theH.seropedicaeRecA protein after 0.42 mM isopropyl-β-D-thiogalactopyranoside (IPTG) addition and 3–4 h incubation.
Protein Purification
The native wild type EcRecA protein was purified using previously described protocols [31,32] and the native wild type HsRecA was purified as follows. All purification steps were carried out at 4°C. Cell paste (approximately 13 g) containing RecA protein was flash-frozen with liquid N2, then thawed overnight on ice in a lysis solution of 250 mM Tris-HCl (80% cation, pH 7.8)
and 25% (w/v) sucrose, adjusting cell to 20% (w/v) ratio. The cells were frozen 2 times in N2,
following addition of lysozyme solution (2.5 mg/mL final concentration lysozyme in 250 mM Tris-HCl (80% cation, pH 7.8), and addition of 0.02 mL of 500 mM EDTA per mL of final lysis solution. The lysate was sonicated for 20 min, using 30s on / 30s off cycle, 60% output, and then centrifuged per 1h30min to remove cells debris. DNA-binding proteins and HsRecA pro-tein was precipitated from the lysate supernatant with addition of 0.111 mL of 5% (w/v) poly-ethyleneimine per mL of lysate and incubated for 1 h. The pellet was washed with R-buffer (20 mM Tris-HCl (80% cation, pH 7.8), 10% glycerol, 0.1 mM EDTA and 1 mM dithiothreitol) and then HsRecA protein was extracted from the pellet by addition of R-buffer + 300 mM ammonium sulfate two times.
and EcRecA concentration were determined using absorbance at 280 nm and the extinction coefficient 1.60×104and 2.23×104M−1cm−1, respectively.
E.colisingle-stranded DNA binding protein (SSB) was purified as described previously [33] and its concentration was determined using an extinction coefficient of 2.83× 104M−1cm−1at
280 nm.
ATPase Assay
ATP hydrolysis was measured by a coupled spectrophotometric assay as previously described [34,35] at 37°C. The assays were carried out in a Varian Cary 300 dual beam spectrophotome-ter equipped with a temperature controller and a 12-position cell changer. Regeneration system of ATP (10 U of pyruvate kinase mL−1and 3 mM phosphoenolpyruvate) from ADP and
phos-phoenolpyruvate is coupled to the conversion of NADH to NAD+(2 mM NADH and 10 U of
lactate dehydrogenase mL−1), which can be monitored by a decrease in absorbance at 380 nm.
Although the maximum absorbance for NADH occurs at 340 nm, absorbances were measured at 380 nm to remain within the linear absorbance range of the spectrophotometer for the extended length of time required in these experiments. The amount of ATP hydrolyzed was calculated using the extinction coefficient of NADH, 1.21 mM-1cm-1. The standard reaction condition was RecA-buffer (25 mM Tris- OAc (80% cation, pH 7.5), 1 mM dithiothreitol, 5% (w/v) glycerol, 3 mM potassium glutamate, and 10 mM magnesium acetate). Concentrations of HsRecA, EcRecA, DNA, SSB, and ATP are indicated in figure legends.E.coliSSB was used to stimulates the ATPase activity of both HsRecA and EcRecA proteins when due.
DNA Three Strand Exchange Reactions
Three strand exchange reactions were carried out as described previously [36,37] in a RecA-buffer at 37°C. An ATP regeneration system (10 U of pyruvate kinase mL−1and 2.5 mM
phos-phoenolpyruvate) was also included. The EcRecA or HsRecA protein were pre-incubated with circular ssDNA M13mp18 (7249 mer) for 20 min, SSB protein and ATP were then added, fol-lowed by another 10 min of incubation. The reaction was initiated by the addition of
M13mp18 linear dsDNA and incubated for 60 min. To stop the reaction, 5μL of a stop and
load reaction buffer (15% Ficoll, 0.25% bromophenol blue, 0.25% xylene cyanol, 25 mM EDTA, and 10% SDS) was added into each 10μL aliquot collected. Samples were subjected to
electrophoresis in 0.8% agarose gels with TAE buffer. Concentrations of HsRecA, EcRecA, DNA, SSB, and ATP are indicated in figure legends.
Electron Microscopy
A modified alcian method was used to visualize RecA filaments. Activated grids were prepared as described previously [20]. Samples for electron microscopy analysis were prepared as follows. All incubations were carried out at 37°C. EcRecA or HsRecA (6.7μM) was preincubated with 20μM
M13mp18 circular ssDNA in a RecA-buffer containing ATP regeneration system (10 U mL-1
cre-atine phosphokinase- PK and 12 mM phosphocrecre-atine- PC) for 20 min. Then, 3 mM ATP with or without 2μM SSB protein were added, and the reaction was incubated for another 10 min. To
stabilize the filaments, 3 mM ATPγS was added and further incubated for 3 min.
The reaction mixture was diluted to a final DNA concentration of 0.4 ng/μL with diluting
water followed by immersion in two 10 mL beakers of double distilled water. After the sample was dried, it was rotary-shadowed with platinum. This protocol is designed for visualization of complete reaction mixtures, and no attempt was made to remove unreacted material. Although this approach should yield results that give a true insight into reaction components, it does lead to samples with a high background of unreacted proteins. Imaging and photography were car-ried out with a TECNAI G2 12 Twin Electron Microscope (FEI Co.) equipped with a GATAN 890 CCD camera. Digital images of the nucleoprotein filaments were taken at X 15000 Magni-fication. Ten filaments each from EcRecA and HsRecA were measured three times using Meta-morph analysis software and the average length was calculated in nm. The 0.5μm scale bar was
used as a standard to calculate the number of pixels perμm.
Crystallization, Data Collection and Structure Refinement
All crystallization experiments were performed using the hanging-drop vapor diffusion method in 24-well plates at 20°C. All conditions were tested with the apo HsRecA form and with the ligands ATPγS and ADP. The initial crystallization screening was carried out using a
reservoir consisting of 1 mL of each composition from the Cryo Suit (QIAGEN), PEG Ion, Natrix, Crystal Screen 1 and 2 and Crystal Screen Lite (Hampton Research) crystallization kits. Each drop contained 1μL reservoir solution and an equal volume of the HsRecA protein
solu-tion (4 mg/mL) plus ligands when used. Clusters of multiple needle crystals were observed after 4 days under condition 0.2 M CaCl2, 20% w/v Polyethylene glycol 3,350 (PEG 3,350), pH
6.8. Further optimization of the condition was performed, initially varying the CaCl2and PEG
concentrations, and then screening for additives by using Additive Screens (Hampton Research). The best single crystals were finally obtained with 0.25 M CaCl2and 14% w/v PEG
3,350 with further addition of Polypropylene glycol P 400 (PPG 400) onto the drop to give a final concentration of 5–8% w/v. Crystals from 0.1 mM HsRecA, 10 mM MgCl2,2 mM ADP
solution reached dimensions of 0.8 mm × 0.2 mm × 0.2 mm after 4 days.
Diffraction data were collected using a Bruker diffractometer equipped with a rotating-anode X-ray generator operated at 40 kV and 40 mA and a Smart-6000 detector. The crystals were flash-cooled in liquid nitrogen at 100 K using the reservoir solution supplemented with 25% ethylene glycol as cryoprotectant. The crystals diffracted to a maximum resolution of 1.7 Å. A total of 1056 frames of data were collected with an oscillation angle of 0.5°, an exposure time of 120 s per frame and a crystal-to-detector distance of 60 mm.
Diffraction data were processed with the proteum2 software (Bruker AXS (2010) PRO-TEUM2, Version 2010.5, Bruker AXS Inc., Madison, Wisconsin, USA). Initial phasing was obtained by molecular replacement with the program Phaser [38]. Two domains ofE.coliRecA
(PDB entry 1xmv, residues 3 to 282, NTD and central ATPase domain, and residues 283 to 328, C-terminal) were used as separate search models. The initial structure was built with Phenix Autobuild [39] and it was completed by repeating cycles of manual model building with COOT [40] and refinement with Phenix.refine [41]. Translation, libration, and screw (TLS) groups were determined with TLSMD [42] and were used during refinement with Phenix.refine.
Results
ATPase Assay and Strand Exchange Activity
To investigate how HsRecA dynamically interacts with ssDNA the DNA-dependent ATPase activity of HsRecA was measured in a coupled spectrophotometric assay using circular ssDNA M13mp18 (cssDNA), ATP, an ATP regeneration system in the presence or absence ofE.coli
A maximum steady state ATPase reaction is typically associated with the formation of com-plete and contiguous RecA filaments on the circular ssDNA [35]. The EcRecA and HsRecA apparentkcatwas determined after the steady-state rate of ATP hydrolysis was achieved,
assum-ing one monomer of RecA bound for each three nucleotides of ssDNA. The results exhibited an apparentkcatof 25.63 ± 0.10 and 28.91 ± 0.11 min-1, to EcRecA and HsRecA, respectively (Fig
1A–Reaction 1). The literature reports akcatof about 30 min-1when EcRecA is bound to ssDNA
under similar conditions [35,43,44]. In multiple trials, the ssDNA-dependent ATPase activity of the HsRecA protein was consistently about 10% higher than that exhibited by EcRecA protein.
The SSB protein is typically used inin vitroassays to remove the ssDNA secondary struc-tures and to facilitate formation of contiguous RecA nucleoprotein filaments when it is added after DNA-RecA pre-incubation. However, when SSB is added prior to RecA, it strongly inhib-its RecA filament nucleation and a long lag is seen before ATP hydrolysis is observed [45–47]. We utilized the EcSSB for all experiments, in part because the HsSSB was not available. Studies of RecA proteins from bacterial species ranging fromNeisseria gonorrhoeae(Ng) to Streptococ-cus pneumoniae(Sp) toDeinococcus radiodurans(Dr) have failed to find any difference in SSB effects when EcSSB replaced cognate SSB proteins in standard RecA assays [48–50]. We note that the HsSSB protein is more closely related to the EcSSB (57% identity/69% similarity) than is the SSB from any of the three species mentioned above (50%/63%, 31%/50%, and 38%/49% for NgSSB, SpSSB, and DrSSB, respectively).
To evaluate the effects of SSB on the ATPase activity of HsRecA, we omitted SSB from the reaction described above (Fig 1A–Reaction 2). In this reaction HsRecA showed higher ATPase activity than EcRecA, with an apparentkcatof 16.49 ± 0.11 and 8.89 ± 0.01 min-1, respectively.
However, neither protein reached the same ATPase activity registered when SSB was present during the 70 minutes evaluated. In addition, a linear ATPase profile throughout the reaction indicated that there was no significant increase of RecA bound to the cssDNA over the course of the experiment.
We also investigated the influence of the moment that SSB is added in HsRecA activity. We added SSB, cssDNA and ATP 10 min before RecA addition, to allow SSB coating the cssDNA (Fig 1B). As already reported in the literature [45–47], EcRecA exhibited a low initial ATPase activity, followed by a very gradual increase, indicating a slow displacement of SSB. However, HsRecA protein displaced the SSB bound to the cssDNA much more effectively. The lag in HsRecA protein mediated ATP hydrolysis was short, and a high rate suggesting formation of contiguous HsRecA filaments was achieved within 10–15 min. HsRecA showed an apparent
kcatof 24.66 ± 1.45 min-1, almost as great as thekcatdetermined in the previous assay when SSB
was added after HsRecA-DNA pre-incubation. In comparison, EcRecA could not reach the same ATPase activity during the 80 min evaluated.
We also examined the ability of the EcRecA and HsRecA proteins to catalyze strand exchange between cssDNA and linear dsDNA (ldsDNA) and form nicked circular dsDNA (NC product). Strand exchange promoted by HsRecA was significantly more efficient than the same reaction promoted by EcRecA. The reaction rates appeared to be similar for the reactions promoted by both proteins, with NC product detectable after 5 min incubation, as well as inter-mediates (Fig 2A). The reactions reached an apparent endpoint after 10 min. However, the extent of reaction was greater for the HsRecA protein at all time points (Fig 2B).
Visualization of HsRecA Filaments by EM
In the absence of the EcSSB protein, EcRecA formed small filamented circles, with discon-tinuous regions being common. On the other hand, HsRecA protein formed more open fila-ments with fewer discontinuities. The addition of SSB eliminated detectable differences between the EcRecA and HsRecA filaments, forming large and completely filamented circles
Fig 1. ATPase profile from EcRecA and HsRecA in the presence and absence of SSB protein.(A) Reaction 1: contained 5μM nt M13mp18 cssDNA and 3μM HsRecA or EcRecA, were previously incubated per 20 min at 37°C, following, 3μM ATP and 0.5μM SSB. Reaction 2: reaction 1 without SSB protein addition. Time 0 min indicates the addition of ATP and SSB. (B) The reaction contained 5μM nt M13mp18 cssDNA, 3μM ATP and 0.35μM SSB, were previously incubated per 10 min at 37°C, following addition of 3μM HsRecA or EcRecA (time 0 min).
on the circular ssDNA substrate. Representative filaments are shown inFig 3, panels A-D. The longer filaments formed by HsRecA in the absence of SSB indicate an enhanced capacity to extend filaments into regions of secondary structure. Note that in these images, only the RecA filaments are generally visible. The ssDNA is not seen unless it is bound with SSB (giving it a distinctive beaded appearance).
The general impressions gained from the initial survey of the grids was confirmed by direct filament length measurements. The length of ten filaments each from EcRecA and HsRecA-cssDNA filaments were measured in the presence and absence of SSB. In the presence of SSB, HsRecA and EcRecA exhibited filament lengths that were indistinguishable. There was no sta-tistical significant difference (Tukey p<0.05) between HsRecA and EcRecA filaments lengths Fig 2. DNA strand exchange promoted by the wild-type HsRecA and EcRecA proteins.(A) The three DNA strand exchange containing 10μM nt M13mp18 cssDNA and 3.5μM HsRecA or EcRecA, were previously incubated per 20 min at
37°C, and then 3μM ATP and 1μM SSB were added and incubated for an additional 10 min. The minutes shown represents
the time of reaction after addition of 20μM nt M13mp18 ldsDNA. (B) The percentage of duplex substrate converted into the
nicked circular duplex (NC product) is plotted against the time.
in the presence of SSB. On the other hand, in the absence of SSB, the HsRecA filaments were significantly longer than those formed by the EcRecA (Fig 3E).
X-Ray Crystal Structure of HsRecA Protein
We solved the X-ray crystal structure ofH.seropedicaeRecA to 1.7 Å resolution (Fig 4). The structure was refined with good bond geometry and crystallographic quality statistics, with no residues in disallowed regions of the Ramachandran space (Table 1). The crystals contained a single HsRecA monomer per asymmetric unit.
Electron density maps derived from molecular replacement phasing permitted assembly of a model of the 351-residue HsRecA protein, with the exception of loops for which density was
Fig 3. Electronic microscopy of EcRecA and HsRecA filaments in the absence or presence of EcSSB. (A) cssDNA filaments without EcSSB, (B) HsRecA-cssDNA filaments without EcSSB, (C) EcRecA-cssDNA filaments with EcSSB, (D) HsRecA-EcRecA-cssDNA filament with EcSSB, (E) Avarege length of 10 HsRecA and EcRecA filaments in the conditions assayed. Letters above each bar refer back to panels A-D. Reactions containing 6.7μM HsRecA or EcRecA and 20μM M13mp18 cssDNA were incubated at 37°C for 20 min, with
or without 2μM Ec SSB. After 10 min, filaments were stabilized by a 3 min incubation with 3μM ATPγS and spread on an Alcian grid.
doi:10.1371/journal.pone.0159871.g003
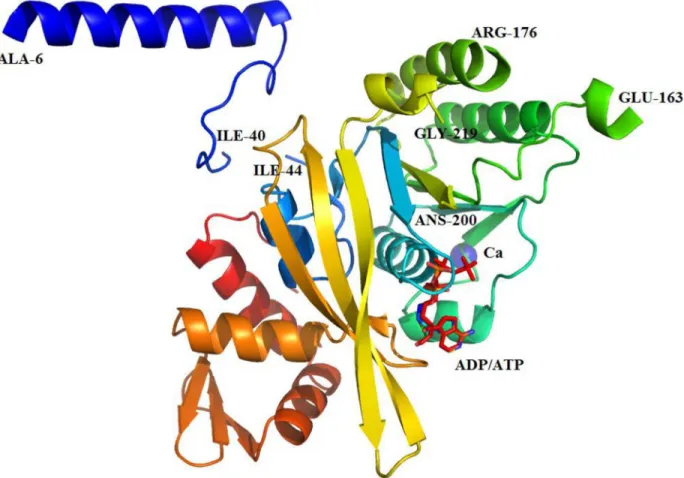
Fig 4. Ribbon diagram of the monomeric crystal structure of HsRecA protein.Regions that could be modeled were indicated by the last residue-number. HsRecA protein is composed of N-terminal domain (NTD), a central core ATPase domain and a large C-terminal domain (CTD). The core ATPase domain contains one Ca2+ion (magenta sphere), coordinated by Asn119 and Asp120 and
the ATPase activity site is partially occupied by ATP and ADP. Figure was prepared using the STRIDE program for secondary structure assignment [55], and visualized using PyMOL [56].
not observed (residues 1–5, 41–43, 164–175, 201–218, and 343–351). As seen in RecA protein structures from other bacterial species [10,11,27,51–53], the tertiary structure of HsRecA can be subdivided into three domains: a small N-terminal domain (NTD), a central core ATPase domain, and a large C-terminal domain (CTD) (Fig 4). The NTD (residues 6–40) consists of a
α-helix and a random coil motif and the CTD (residues 276–343) consists of 3α-helices and one antiparallelβ-sheet motif. The core domain (residues 44–275) consists of a conservedα/β
ATPase motif, which in its turn contains the Walker A (residues 72–81) and B (residues 147–
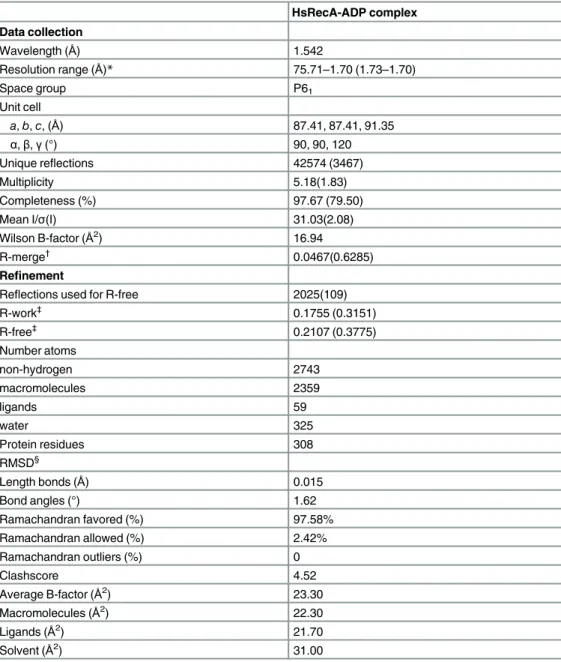
Table 1. Data collection and refinement statistics.
HsRecA-ADP complex Data collection
Wavelength (Å) 1.542
Resolution range (Å)* 75.71–1.70 (1.73–1.70)
Space group P61
Unit cell
a,b,c, (Å) 87.41, 87.41, 91.35
α,β,γ(°) 90, 90, 120
Unique reflections 42574 (3467)
Multiplicity 5.18(1.83)
Completeness (%) 97.67 (79.50)
Mean I/σ(I) 31.03(2.08)
Wilson B-factor (Å2) 16.94
R-merge†
0.0467(0.6285) Refinement
Reflections used for R-free 2025(109) R-work‡ 0.1755 (0.3151) R-free‡ 0.2107 (0.3775) Number atoms non-hydrogen 2743 macromolecules 2359 ligands 59 water 325
Protein residues 308
RMSD§
Length bonds (Å) 0.015
Bond angles (°) 1.62
Ramachandran favored (%) 97.58%
Ramachandran allowed (%) 2.42%
Ramachandran outliers (%) 0
Clashscore 4.52
Average B-factor (Å2) 23.30
Macromolecules (Å2
) 22.30
Ligands (Å2
) 21.70
Solvent (Å2) 31.00
*Statistics for the highest-resolution shell are shown in parentheses.
†R
merge=Σ|I-<I>|/ΣI, where I is the observed intensity. ‡R
workand Rfree=Σ|Fobs-Fcalc|/ΣFobswhere Rfreewas calculated over 5% of the amplitude chosen at random
and not used in the refinement.
§RMSD, root-mean-square deviation.
157) motifs, and the disordered DNA binding loops L1(residues 163–176) and L2(residues
202–219) for which electron density was not observed in our structure. A Ca2+ion is bound by Asn119 and Asp120 in the ATPase core. Proper Ca2+coordination in the structure was
con-firmed by the CheckMyMetal web server [54].
The amino acid sequence alignment from HsRecA and EcRecA, shows that these two pro-teins shares 68.12% identity and 80.6% similarity. The overall structure of HsRecA is similar to that of EcRecA (PDB entry, 1XMV) with root-mean square deviation (RMSD) of 1.324Åfor Cα, 1.335Åfor backbone and 1.486Åfor all atoms. The structural comparison was performed using LSQKAB from the CCP4 program suite [57].
ATPase Active Site
The co-crystallized adenosine diphosphate (ADP) interacts directly with Ser76, Ser77, Thr80, Thr81, Asp107, Tyr110, and Gly272 in the HsRecA structure. Despite the fact that Mg2+ion
was present in the crystallization solution there was no electron density for this ion around the nucleotide ADP to suggest proper coordination. Mg2+ion was also not observed in
Deinococ-cus radioduransandMycobacterium tuberculosisRecA crystal structures [53,58]. We observed an extra electron density beyond theβ-phosphate of ADP which could not be properly
mod-eled with waters, SO43-or PO43-ions. The refinement of PO43-and ADP as the only nucleotide
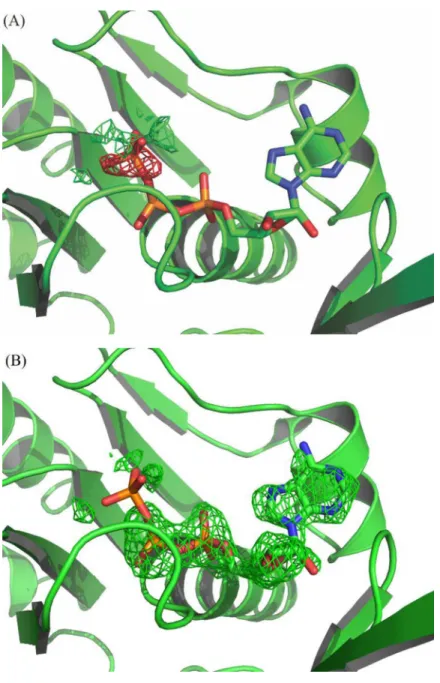
showed electron density that suggests them to be covalently linked, but there also remained residual negative electron density in the Fourier difference map over PO4-3at 100% occupancy
(Fig 5A and 5B). Therefore, we considered that this site is partially occupied by ATP, which might have come from the cells. The refinement with ADP and ATP sharing the position improved the adjustment to the electron density and resulted in occupancies of 61% and 39%, respectively. The amide nitrogen of Gln201 is at 2.88 Å from theγ-phosphate. This residue pre-cedes the DNA binding Loop L2and plays a role in the activation of the RecA function by
induction of an allosteric effect of ATP [59]. Yet in the DrRecA-ATPγS complex this distance is 3.0 Å from theγ-thiophosphate. Both glutamines present the same conformation, different
from the EcRecA structure (pdb entry 1xmv) which is bound to ADP.
Assembly of HsRecA Protein in the Crystal Structure
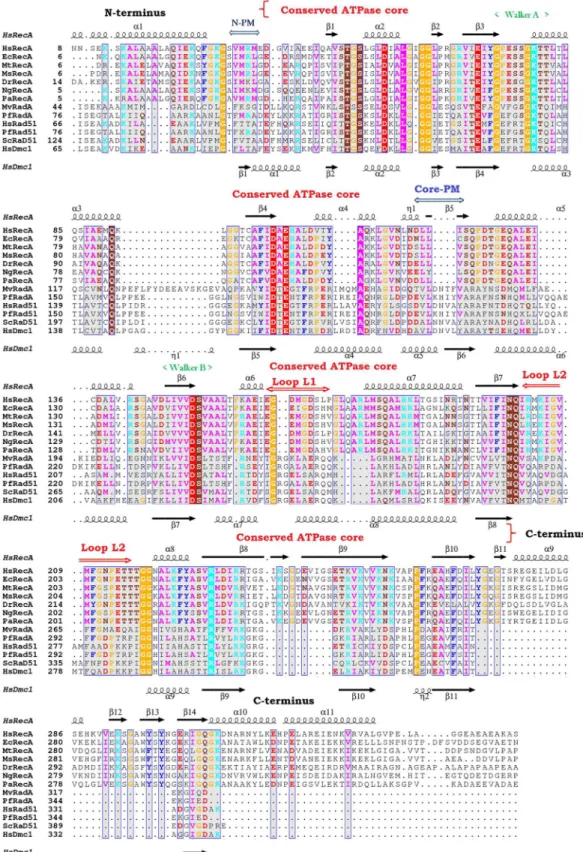
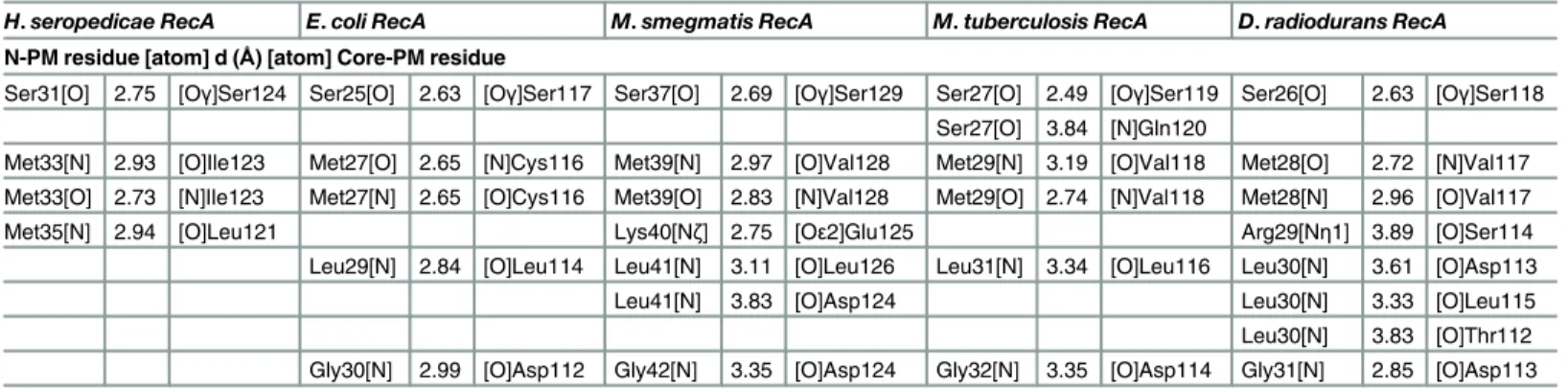
Bacterial RecA and other members of the RecA family (RadA, Dmc1 and Rad51) were aligned using their primary and tertiary structures to identify common regions (Fig 6). The monomer assembly regions were first identified in the RadA/Rad51/Dcm1 proteins [12,13,21,22,60]. They encompass residues in the NTD (Fig 6, N-PM region) and in the core ATPase domain (Fig 6, Core-PM region). The crystal structures of several RecA family members have revealed that the N-PM region is a polymerization motif, which is a shortβ-strand that just precedes the ATPase core and has conserved hydrophobic residues. Yet, using the alignment and the protein structures from the bacteriaE.coli(pdb entry 3CMU, 1XMS, 1U98 and 1XMV),D. radiodur-ans(pdb entry 1XP8),M.smegmatis(pdb entry 1UBE, 1UBF and 1UBG),M.tuberculosis(pdb entry 1MO3, 1MO5, 1MO6 and 1G18) and ours, we noticed that this polymerization motif (N-PM) consists of a conserved sequence (Ser, Val/Ile, Met, Arg/Lys, Leu, Gly; residues 25–30 in the EcRecA sequence after elimination of the N-terminal Met, and residues 30–35 in the HsRecA sequence). This sequence interacts with the core ATPase domain (Core-PM) of the adjacent subunit at another conserved sequence (Asp, Asn, Leu, Leu, Val/Cys, Ser; residues 113–118 and 118–123 in the EcRecA and HsRecA sequences, respectively).
residues. The conserved methionine in the N-PM may play a role in binding/recognition of the hydrophobicβ-strand polymerization motif in the core ATPase domain, promoting thus the polymerization of RecA filaments on the DNA substrate. TheTable 2shows the hydrogen bonds formed between the N-PM and the Core-PM from RecA proteins.
In all inactive RecA protein structures, the shortβ-strand motif becomes aβ-loop motif
con-formation in the NTD (Fig 7A). This motif is observed in the HsRecA structure as well, although in the EcRecA-ssDNA presynaptic filament this stands as aβ-strand motif, as found
in the RecA family proteins (Fig 7B) and it is an antiparallelβ-strand with respect to the second motif in the EcRecA-ssDNA pre-synaptic crystal structure filament.
Fig 5. Omit maps (mFo-DFc) at the ATP binding site:(A) Refinement with ATP at 100% occupancy; the
negative electron density over theγ-phosphate indicates that it should not be at this full occupancy. (B) Refinement with ATP at 39% occupancy; the positive electron density over the corresponding ADP moiety indicates that it should be at full occupancy. The omit maps are contoured at +3 (green) and -3 (red)σlevels.
The Ca2+ion in the HsRecA Core-PM is coordinated by Asn119 and Asp120 and the Mg2+
ion in the EcRecA N-PM is coordinated by Glu18, Lys23 and Ile26 [27]. Both divalent ions may play a role in the formation and stability of RecA protein as a helical nucleoprotein fila-ment. Previous studies showed that addition of excess Mg2+(relative to the available ATP)
helps to produce the active and extended conformation of the filament, required for DNA pair-ing and the strand exchange reactionin vitro[66]. HsRecA E36 is at the subunit interface
bor-der and might interact with excess ions along with other residues nearby, possibly increase the interaction within the subunit-subunit surface.
Electrostatic Potential Surface
We also reconstructed the HsRecA protein on the basis of its electrostatic potentials in order to understand the structural basis of their functional activities and subunit-subunit interactions. The electrostatic potential distribution on the solvent-accessible surface of the HsRecA protein for two subsequent helical hexamers is shown inFig 8.
Due to the CTD portion of each monomer, there is a relatively large negatively charged region on the outer surface in the HsRecA filament structure. As has been observed for many RecA proteins, HsRecA protein has increased positive charge along the inner side of the fila-ment, near the DNA binding sites (Fig 8B). There are subtle differences in the charge density and distribution relative to EcRecA protein,. These have not been analyzed in detail, but may eventually help explain the DNA binding properties of HsRecA protein. Important domains that affect filament polymerization stand orderly along the filament axis, such that there is a charge polarity change from negative to positive that binds the DNA from the 30to 50direction
(Fig 8B and 8C). Therefore, we speculate that electrostatic interactions would facilitate proper filament assembly, after monomers had been stabilized by hydrophobic interactions between the N-PM of one monomer and the Core-PM of the subsequent one.
Helical Pitch
The crystallography61symmetry leads to the formation of a helical filament that has a pitch of
91.3 Å (Fig 9), a value close to the pitch range 90–100 Å determined by electron microscopy
corresponding amino acid sequences: N-PM and core-PM, N-terminal and core polymerization motif, respectively, the ATP binding Walker A and B motifs, the putative DNA binding sites Loop L1and L2. The
secondary structure of HsRecA and HsDmc1 are indicated at the top and bottom, respectively. The 3D structural and amino acid alignment was performed using the MultiProt and T-Coffee programs [61,62]. The results are visualized using ESPript—http://espript.ibcp.fr[63].
doi:10.1371/journal.pone.0159871.g006
Table 2. Summary of hydrogen bonds formed between the N-PM and the Core-PM in RecA protein structures.The residues and their atoms involved in the interaction are indicated, as well as the atomic distances. Interactions were determined using the CCP4 application (Protein Interfaces, Surfaces and Assemblies—PISA) [52,64,65].
H.seropedicae RecA E.coli RecA M.smegmatis RecA M.tuberculosis RecA D.radiodurans RecA N-PM residue [atom] d (Å) [atom] Core-PM residue
Ser31[O] 2.75 [Oγ]Ser124 Ser25[O] 2.63 [Oγ]Ser117 Ser37[O] 2.69 [Oγ]Ser129 Ser27[O] 2.49 [Oγ]Ser119 Ser26[O] 2.63 [Oγ]Ser118 Ser27[O] 3.84 [N]Gln120
Met33[N] 2.93 [O]Ile123 Met27[O] 2.65 [N]Cys116 Met39[N] 2.97 [O]Val128 Met29[N] 3.19 [O]Val118 Met28[O] 2.72 [N]Val117 Met33[O] 2.73 [N]Ile123 Met27[N] 2.65 [O]Cys116 Met39[O] 2.83 [N]Val128 Met29[O] 2.74 [N]Val118 Met28[N] 2.96 [O]Val117
Met35[N] 2.94 [O]Leu121 Lys40[Nζ] 2.75 [Oε2]Glu125 Arg29[Nη1] 3.89 [O]Ser114
Leu29[N] 2.84 [O]Leu114 Leu41[N] 3.11 [O]Leu126 Leu31[N] 3.34 [O]Leu116 Leu30[N] 3.61 [O]Asp113
Leu41[N] 3.83 [O]Asp124 Leu30[N] 3.33 [O]Leu115
for active filaments of EcRecA formed in the presence of DNA, ATP-γ-S or ATP [70]. Inactive and compressed filaments characterized to date have a helical pitch of 65–85 Å and are formed by RecA alone or bound to ADP, either in the absence of DNA or bound to ssDNA or dsDNA [10,26,51,53,58,71]. Despite the fact that HsRecA-ADP/ATP protein was crystallized with ADP and ATP, our structure presented a helical pitch characteristic of an active RecA filament form. However, as noted above, the structural packing is that of the inactive state.
Discussion
In the present work, we report a functional and structural characterization of the RecA protein fromH.seropedicae, an important bacterium to environmental crops in agricultural systems. The work has several conclusions. The HsRecA protein forms filaments on ssDNA, and pro-motes ATP hydrolysis and DNA strand exchange as do other bacterial RecA proteins. The HsRecA distinguishes itself with a somewhat more efficient DNA strand exchange reaction, and a moderately increased rate of ATP hydrolysis relative to that promoted by EcRecA pro-tein. The HsRecA also displaces pre-bound SSB protein much faster than EcRecA. The struc-ture of HsRecA protein, also presented here, is consistent with the RecA strucstruc-tures found for RecA proteins from other bacteria [10,11,27,51–53]. However, in the absence of DNA, the HsRecA crystal structure showed a helical pitch closer to the extended form of a RecA filament, although the structural packing remained that of the inactive state. Structural comparisons offer an opportunity to gain new structure-function insights.
The amino acid sequence alignment fromH.seropedicaeSSB andE.coliSSB, shows that these two proteins are 57.1% identical and 69.0% similar. Our results showed that theE.coli
SSB protein stimulates the formation of contiguous HsRecA nucleoprotein filaments (Fig 1A–
Reaction 1), as previously seen forE.coli,Pseudomonas aeruginosaandNeisseria gonorrhoeae
RecA nucleoproteins [45,72,48,73]. However, the effect of SSB is different inD.radiodurans: eitherE.coliorD.radioduransSSB protein suppresses DrRecA ATPase activityin vitro[74].
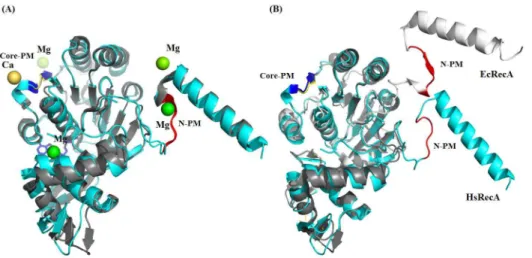
Fig 7. Superposition inactive and presynaptic monomers of EcRecA protein over the inactive HsRecA monomer.EcRecA protein is colored in gray and HsRecA in cyan. (A) Superposition of the inactive EcRecA structure (pdb entry 1XMV) over the HsRecA structure. Both structures have aβ-loop motif in the N-PM. In the EcRecA structure, a Mg2+ion interacts with the N-PM. In the HsRecA structure, a Ca2+ion interacts with
the Core-PM. (B) Superposition of presynaptic monomers of EcRecA structure (pdb entry 3CMU:A) over the HsRecA structure. The N-PM in the active EcRecA structure assumes aβ-strand motif. The nucleotides ADP and ADP-AlF4, Mg2+and Ca2+ions, and the ssDNA bound in the EcRecA structure was removed for clarity.
Figure was prepared using the STRIDE program for secondary structure assignment [55]. The superposition was generated using the MultiProt program [61] and visualized using PyMOL [56].
HsRecA reached an apparentkcatof 28.91 ± 0.11 min-1. When compared under the same
con-ditions, these rates were about 10% higher than those observed when EcRecA was bound to ssDNA.
The HsRecA protein has a higher capacity to bind to regions of secondary structure in ssDNA. Omission of the SSB from the ATPase assay decreases EcRecA ATPase activity about 65% (kcatof 8.89 ± 0.01 min-1), while for HsRecA protein this reduction was about 43% (kcatof
16.49 ± 0.11) (Fig 1A–Reaction 2). Based on the fact the ATPase activity is correlated to the amount of RecA bound to DNA, we suggest that HsRecA can better bind to the secondary struc-ture of ssDNA in absence of SSB than the other proteins previously reported. Electron
Fig 8. Electrostatic potential distribution on the solvent-accessible surface of HsRecA protein structure.(A) Side view of two subsequent helical hexamers. (B) and (C), 50and 30views, relative to the axial direction of the filament, respectively. The surface potential representation has charge levels from -3kT/e(red) to +3kT/e(blue). The electrostatic potential distribution was generated using the APBS program, side chain atom not ordered in the crystal were added using the PDB2PQR program and protonation states at pH 7.5 were assigned with the PROPKA program [67–69], and visualized using PyMOL [56].
microscopy experiments performed to visualize EcRecA and HsRecA interaction with cssDNA and ATPγS also demonstrated a greater capacity to bind secondary structure for the HsRecA
pro-tein. When the SSB protein was absent, HsRecA filament lengths were significantly greater on M13mp18 ssDNA than those observed for EcRecA. When SSB protein was added after RecA and DNA pre-incubation, the filament lengths and characteristics were indistinguishable.
HsRecA protein displaces pre-bound SSB much more quickly than does EcRecA, and reaches a steady state with an apparentkcatof 24.66 ± 1.45 min-1within 10–15 min (Fig 1B).
SSB protein is important in recombination processes because it stimulates the first phase, pre-synaptic polymerization of RecA protein on ssDNA, and the last phase, strand exchange [75–
77]. Previous studies showed that some mutations in EcRecA (recA730 (E38K) [78], recA803 (V37M) [79], recA2020 (T121I) [80] and recA441 (E38K/I298V) [78,81,82]) can produce a similar capacity for bypassing the SSB block to nucleation. In HsRecA protein, these residues are conserved, except V37 and E38, which correspond to I44 and Q45 in HsRecA, respectively (Fig 6). Like EcRecA, the HsRecA has a C-terminus that is predominantly negatively charged (four Glu residues and one Lys among the 16 C-terminal amino acid residues. The concentra-tion of negative charge is greater in the EcRecA C-terminus (7 of the last 17 amino acid resi-dues, with no positive charges). Elimination of the EcRecA C-terminus also results in faster nucleation on SSB-coated ssDNA [20]. We have postulated that an interaction between the RecA C-terminus (unstructured in most RecA protein structures, including this one) and the surface at E38 could mask a RecA surface required for rapid SSB displacement. We postulate that the decrease in negative charge in the HsRecA, coupled with the amino acid residue substi-tutions at positions 44 and 45, may decrease the masking effect of the C-terminus and permit the observed rapid SSB displacement.
The monomeric structure of native HsRecA protein exhibits an architecture similar to that of bacterial RecA proteins previously crystallized. HsRecA protein has a small NTD, a core ATPase domain and a large CTD, with the same secondary structure elements of bacterial RecA proteins [9,51,53,58]. However, the crystallography61symmetry leads to the formation
of a helical filament that has a pitch of 91.3 Å (Fig 9), a value within the pitch range of 90–100 Å determined by electron microscopy for active filaments of EcRecA formed in the presence of DNA, ATP-γ-S or ATP [70]. In contrast, the inactive and compressed filament has a helical
pitch of 65–85 Å and is formed by RecA alone or bound to ADP in the absence of DNA [10,26,51,53,58,71]. Despite the fact that HsRecA-ADP/ATP protein formed an inactive fila-ment in the presence of ADP and absence of DNA, our structure presented a helical pitch of an extended RecA filament form. Although previous studies using single particle analysis revealed that there is a considerable overlap in pitch between the active and inactive states [83], high salt concentrations induce RecA ATP hydrolysis and also facilitate the crystallization of the extended filament in the absence of ssDNA [84–86]. Curiously, polyethylene glycol (PEG) of various sizes also increasedM.voltaeRadA ATPase activity in the absence of DNA [87]. We suggest that the high concentrations of salt and PEG used in the crystallization solution of HsRecA (0.25 M CaCl2,14% w/v PEG 3,350 and 8% w/v PPG 400) could explain the
crystalli-zation of the HsRecA extended filament form even in the absence of DNA.
The mechanism of polymerization of RadA/Rad51 proteins as helical filaments have already been reported [17]. However, analyzing the primary and tertiary structures from bacterial RecA protein we found some differences. A conserved residue sequence in the NTD plays a role in the
Fig 9. Ribbon diagram of two subsequent helical hexamers of HsRecA protein.Each monomer is colored differently. The hexameric structure has 61-fold symmetry, the helical filament has a pitch of 91.3Å. Figures were
polymerization: hydrophobic residues from two motifs (N-PM and Core-PM) stabilize the sub-unit-subunit interaction in inactive structures. The N-PM assumes aβ-loop conformation, while the Core-PM assumes aβ-strand conformation. Sequence alignment revealed the presence of a
conserved methionine in the bacterial N-PM RecAs, which we speculate may play a role in bind-ing/recognition of the hydrophobicβ-strand polymerization motif in the core ATPase domain. We
also noticed that the transition of inactive and compressed to active and extended RecA structure follows the transition from aβ-loop to aβ-strand motif conformation in N-PM. The presence of
the divalent ions Mg2+and Ca2+as observed in the N-PM and Core-PM, respectively, in EcRecA (pdb entry 1XMV) and HsRecA structures may play a role in the structural transition from com-pressed to extended conformation. This is similar to structural transitions in yeast Dmc1 protein, in which Ca2+ion promotes the formation of an extended helical filament onto ssDNA [88].
The polymerization and dissociation of RecA protein from DNA are regulated by an array of proteins [6]. HsRecA was efficient at displacing SSB from ssDNA. In its absence, HsRecA was still able to bind more of the M13mp18 ssDNA circle, with its extensive secondary struc-ture, than was observed with EcRecA when ATP or ATPγS is present. Most bacterial RecAs are
dependent on RecO and RecR to displace SSB from DNA [89,90], but HsRecA protein appears to require less assistance. TheH.seropedicaeSSB, RecO and RecR proteins may play a some-what different role in the recombination processes of this bacterium. Further experiments need to be performed to investigate the role ofH.seropedicaeSSB, RecO and RecR proteins in the mechanism of polymerization and stability of HsRecA filaments.
The HsRecA protein has a greater capacity to bind ssDNA in the presence of SSB, and this may reflect the plant-bacteria interaction and the apparent absence of some auxiliary proteins normally associated with RecA. RecA protein has recently been implicated in bacterial swarming, a flagellar-driven highly coordinated translocation of a bacterial colony across a moist surface [91,92]. This motility has been linked to a chemotaxis signaling pathway which includes methyl-accepting chemotaxis proteins (MCPs), CheW adaptor proteins and CheA kinase [93]. Irazoki
et al. (2016) [94] showed that activation of the SOS response by the presence of a DNA-damaging compound increases the RecA concentration, thereby disturbing the equilibrium between RecA and CheW and resulting in the cessation of swarming. When the DNA-damaging source decreases or disappears, the repair of the DNA damage seems to restore colony swarming ability [94]. Based on the fact thatH.seropedicaeis subjected to many environmental factors that cause DNA-damage, such as solar ultraviolet radiation, reactive oxygen species, pH, among others, the enhanced capacity of HsRecA to bind DNA may facililtate more rapid repair and reestablishment of swarming, providing the bacteria a competitive advantage in colonization of plant root sur-faces. Many genes related to chemotaxis were found inH.seropedicaeSMR1 genome but further experiments are needed to evaluate the role of RecA to swarming in this bacterium.
Acknowledgments
We would like to thank Elizabeth Wood for sub cloning therecAgene ofH.seropedicaeused in this study, Tricia Windgassen for crystallographic advice, and Fabio de Oliveira Pedrosa and Emanuel Maltempi de Souza for scientific and infrastructure support.
Author Contributions
References
1. Kowalczykowski SC, Eggleston a K. Homologous pairing and DNA strand-exchange proteins. Annu Rev Biochem. 1994; 63: 991–1043. doi:10.1146/annurev.bi.63.070194.005015PMID:7979259
2. Cox MM. The bacterial RecA protein as a motor protein. Annu Rev Microbiol. 2003; 57: 551–577. doi:
10.1146/annurev.micro.57.030502.090953PMID:14527291
3. Janion C. Inducible SOS response system of DNA repair and mutagenesis in Escherichia coli. Int J Biol Sci. 2008; 4: 338–44. doi:10.7150/ijbs.4.338PMID:18825275
4. Cox MM. The Bacterial RecA Protein: Structure, Function, and Regulation. Molecular Genetics of Recombination. Berlin, Heidelberg: Springer Berlin Heidelberg; 2007. pp. 53–94. doi:10.1007/4735_ 2006_0205
5. Cox MM. Motoring along with the bacterial RecA protein. Nat Rev Mol Cell Biol. Nature Publishing Group; 2007; 8: 127–138. doi:10.1038/nrm2099PMID:17228330
6. Cox MM. Regulation of Bacterial RecA Protein Function. Crit Rev Biochem Mol Biol. Taylor & Francis; 2008; doi:10.1080/10409230701260258
7. Cox MM, Goodman MF, Kreuzer KN, Sherratt DJ, Sandler SJ, Marians KJ. The importance of repairing stalled replication forks. Nature. 2000; 404: 37–41. doi:10.1038/35003501PMID:10716434
8. Lusetti SL, Cox MM. The bacterial RecA protein and the recombinational DNA repair of stalled replica-tion forks. Annu Rev Biochem. 2002; 71: 71–100. doi:10.1146/annurev.biochem.71.083101.133940
PMID:12045091
9. Story RM, Weber IT, Steitz TA. The structure of the E. coli recA protein monomer and polymer. Nature. 1992; 355: 318–325. doi:10.1038/355318a0PMID:1731246
10. Story RM, Steitz TA. Structure of the recA protein-ADP complex. Nature. 1992; 355: 374–6. doi:10. 1038/355374a0PMID:1731253
11. Chen Z, Yang H, Pavletich NP. Mechanism of homologous recombination from the RecA-ssDNA/ dsDNA structures. Nature. 2008; 453: 489–484. doi:10.1038/nature06971PMID:18497818
12. Chen LT, Ko TP, Chang YC, Lin KA, Chang CS, Wang AHJ, et al. Crystal structure of the left-handed archaeal RadA helical filament: Identification of a functional motif for controlling quaternary structures and enzymatic functions of RecA family proteins. Nucleic Acids Res. 2007; 35: 1787–1801. doi:10. 1093/nar/gkl1131PMID:17329376
13. Wu Y, He Y, Moya IA, Qian X, Luo Y. Crystal structure of archaeal recombinase RADA: a snapshot of its extended conformation. Mol Cell. 2004; 15: 423–35. doi:10.1016/j.molcel.2004.07.014PMID:
15304222
14. Kinebuchi T, Kagawa W, Enomoto R, Tanaka K, Miyagawa K, Shibata T, et al. Structural Basis for Octameric Ring Formation and DNA Interaction of the Human Homologous-Pairing Protein Dmc1. Mol Cell. 2004; 14: 363–374. doi:10.1016/S1097-2765(04)00218-7PMID:15125839
15. Okorokov AL, Chaban YL, Bugreev D V, Hodgkinson J, Mazin A V, Orlova E V. Structure of the hDmc1-ssDNA filament reveals the principles of its architecture. PLoS One. 2010; 5: e8586. doi:10.1371/ journal.pone.0008586PMID:20062530
16. Sandler SJ, Satin LH, Samra HS, Clark AJ. RecA-like genes from three archaean species with putative protein products similar to Rad51 and Dmc1 proteins of the yeast Saccharomyces cerevisiae. Nucleic Acids Res. 1996; 24: 2125–2132. doi:10.1093/nar/24.11.2125PMID:8668545
17. Chang Y-W, Ko T-P, Lee C-D, Chang Y-C, Lin K-A, Chang C-S, et al. Three new structures of left-handed RADA helical filaments: structural flexibility of N-terminal domain is critical for recombinase activity. PLoS One. 2009; 4: e4890. doi:10.1371/journal.pone.0004890PMID:19295907
18. Wu Y, Qian X, He Y, Moya IA, Luo Y. Crystal structure of an ATPase-active form of Rad51 homolog from Methanococcus voltae. Insights into potassium dependence. J Biol Chem. 2005; 280: 722–8. doi:
10.1074/jbc.M411093200PMID:15537659
19. Aihara H, Ito Y, Kurumizaka H, Yokoyama S, Shibata T. The N-terminal domain of the human Rad51 protein binds DNA: structure and a DNA binding surface as revealed by NMR. J Mol Biol. 1999; 290: 495–504. doi:10.1006/jmbi.1999.2904PMID:10390347
20. Eggler AL, Lusetti SL, Cox MM. The C terminus of the Escherichia coli RecA protein modulates the DNA binding competition with single-stranded DNA-binding protein. J Biol Chem. 2003; 278: 16389–
96. doi:10.1074/jbc.M212920200PMID:12598538
21. Lee C-D, Wang T-F. The N-terminal domain of Escherichia coli RecA have multiple functions in promot-ing homologous recombination. J Biomed Sci. 2009; 16: 37. doi:10.1186/1423-0127-16-37PMID:
22. Wang T-F, Chen L-T, Wang AH-J. Right or left turn? RecA family protein filaments promote homolo-gous recombination through clockwise axial rotation. Bioessays. 2008; 30: 48–56. doi:10.1002/bies. 20694PMID:18081011
23. Bell JC, Plank JL, Dombrowski CC, Kowalczykowski SC. Direct imaging of RecA nucleation and growth on single molecules of SSB-coated ssDNA. Nature. Nature Publishing Group, a division of Macmillan Publishers Limited. All Rights Reserved.; 2012; 491: 274–8. doi:10.1038/nature11598PMID:
23103864
24. Lindsleys E. Assembly and Disassembly of RecA Protein Filaments Opposite Filament Ends Occur at. 1990; 265: 9043–9054.
25. Shan Q, Bork JM, Webb BL, Inman RB, Cox MM. RecA protein filaments: end-dependent dissociation from ssDNA and stabilization by RecO and RecR proteins. J Mol Biol. 1997; 265: 519–40. doi:10.1006/ jmbi.1996.0748PMID:9048946
26. Galletto R, Amitani I, Baskin RJ, Kowalczykowski SC. Direct observation of individual RecA filaments assembling on single DNA molecules. Nature. 2006; 443: 875–8. doi:10.1038/nature05197PMID:
16988658
27. Xing X, Bell CE. Crystal structures of Escherichia coli RecA in a compressed helical filament. J Mol Biol. 2004; 342: 1471–85. doi:10.1016/j.jmb.2004.07.091PMID:15364575
28. Roca AI, Cox MM. RecA protein: structure, function, and role in recombinational DNA repair. Prog Nucleic Acid Res Mol Biol. 1997; 56: 129–223. Available:http://www.ncbi.nlm.nih.gov/pubmed/ 9187054. PMID:9187054
29. Galvão CW, Souza EM, Etto RM, Pedrosa FO, Chubatsu LS, Yates MG, et al. The RecX protein inter-acts with the RecA protein and modulates its activity in Herbaspirillum seropedicae. Braz J Med Biol Res. 2012; 45: 1127–34. doi:10.1590/s0100-879x2012007500160PMID:23044625
30. Pedrosa FO, Monteiro RA, Wassem R, Cruz LM, Ayub RA, Colauto NB, et al. Genome of Herbaspiril-lum seropedicae strain SmR1, a specialized diazotrophic endophyte of tropical grasses. PLoS Genet. 2011; 7: e1002064. doi:10.1371/journal.pgen.1002064PMID:21589895
31. Cox MM, McEntee K, Lehman IR. A simple and rapid procedure for the large scale purification of the recA protein of Escherichia coli. J Biol Chem. 1981; 256: 4676–4678. Available:http://www.jbc.org/ content/256/9/4676.abstract?ijkey=ebf61ddcdd61a793ee7acabcf72ece76a020aed5&keytype2=tf_ ipsecsha. PMID:7012155
32. Shan Q, Cox MM, Inman RB. DNA Strand Exchange Promoted by RecA K72R: TWO REACTION PHASES WITH DIFFERENT Mg REQUIREMENTS. J Biol Chem. 1996; 271: 5712–5724. doi:10. 1074/jbc.271.10.5712PMID:8621437
33. Lohman TM, Green JM, Beyer RS. Large-scale overproduction and rapid purification of the Escherichia coli ssb gene product. Expression of the ssb gene under.lambda. PL control. Biochemistry. American Chemical Society; 1986; 25: 21–25. doi:10.1021/bi00349a004PMID:3006753
34. Morrical SW, Lee J, Cox MM. Continuous association of Escherichia coli single-stranded DNA binding protein with stable complexes of recA protein and single-stranded DNA. Biochemistry. 1986; 25: 1482–
94. doi:10.1021/bi00355a003PMID:2939874
35. Brenner SL, Mitchell RS, Morrical SW, Neuendorf SK, Schutte BC, Cox MM. recA protein-promoted ATP hydrolysis occurs throughout recA nucleoprotein filaments. J Biol Chem. 1987; 262: 4011–4016. PMID:2951381
36. Cox MM, Lehman IR. Directionality and polarity in recA protein-promoted branch migration. Proc Natl Acad Sci U S A. 1981; 78: 6018–22. Available:http://www.pubmedcentral.nih.gov/articlerender.fcgi? artid=348968&tool=pmcentrez&rendertype=abstract. PMID:6273839
37. Bedale WA, Cox M. Evidence for the coupling of ATP hydrolysis to the final (extension) phase of RecA protein-mediated DNA strand exchange. J Biol Chem. 1996; 271: 5725–32. doi:10.1074/jbc.271.10. 5725PMID:8621438
38. McCoy AJ, Grosse-Kunstleve RW, Adams PD, Winn MD, Storoni LC, Read RJ. Phaser crystallographic software. J Appl Crystallogr. International Union of Crystallography; 2007; 40: 658–674. doi:10.1107/ S0021889807021206PMID:19461840
39. Terwilliger TC, Grosse-Kunstleve RW, Afonine P V, Moriarty NW, Zwart PH, Hung LW, et al. Iterative model building, structure refinement and density modification with the PHENIX AutoBuild wizard. Acta Crystallogr D Biol Crystallogr. International Union of Crystallography; 2008; 64: 61–9. doi:10.1107/ S090744490705024XPMID:18094468
40. Emsley P, Lohkamp B, Scott WG, Cowtan K. Features and development of Coot. Acta Crystallogr D Biol Crystallogr. International Union of Crystallography; 2010; 66: 486–501. doi:10.1107/
41. Adams PD, Afonine P V, Bunkóczi G, Chen VB, Davis IW, Echols N, et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr D Biol Crystallogr. Inter-national Union of Crystallography; 2010; 66: 213–21. doi:10.1107/S0907444909052925PMID:
20124702
42. Painter J, Merritt EA. Optimal description of a protein structure in terms of multiple groups undergoing TLS motion. Acta Crystallogr D Biol Crystallogr. International Union of Crystallography; 2006; 62: 439–
50. doi:10.1107/S0907444906005270PMID:16552146
43. Bianco PR, Weinstock GM. Interaction of the RecA protein of Escherichia coli with single-stranded oli-godeoxyribonucleotides. Nucleic Acids Res. 1996; 24: 4933–4939. doi:10.1093/nar/24.24.4933PMID:
9016663
44. Weinstock G, McEntee K, Lehman I. Hydrolysis of nucleoside triphosphates catalyzed by the recA pro-tein of Escherichia coli. Hydrolysis of UTP. J Biol Chem. 1981; 256: 8856–8858. Available:http://www. jbc.org/content/256/16/8856.abstract?ijkey=
fa6e333f30de803dbb1d35a529146ef725df2e16&keytype2=tf_ipsecsha. PMID:7021554
45. Kowalczykowski SC, Clow J, Somani R, Varghese A. Effects of the Escherichia coli SSB protein on the binding of Escherichia coli RecA protein to single-stranded DNA. Demonstration of competitive binding and the lack of a specific protein-protein interaction. J Mol Biol. 1987; 193: 81–95. Available:http:// www.ncbi.nlm.nih.gov/pubmed/3295259. PMID:3295259
46. Umezu K, Kolodner RD. Protein interactions in genetic recombination in Escherichia coli. Interactions involving RecO and RecR overcome the inhibition of RecA by single-stranded DNA-binding protein. J Biol Chem. 1994; 269: 30005–13. Available:http://www.ncbi.nlm.nih.gov/pubmed/7962001. PMID:
7962001
47. Shereda RD, Kozlov AG, Lohman TM, Cox MM, Keck JL. SSB as an organizer/mobilizer of genome maintenance complexes. Crit Rev Biochem Mol Biol. 2008; 43: 289–318. doi:10.1080/
10409230802341296PMID:18937104
48. Stohl EA, Gruenig MC, Cox MM, Seifert HS. Purification and characterization of the RecA protein from Neisseria gonorrhoeae. PLoS One. Public Library of Science; 2011; 6: e17101. doi:10.1371/journal. pone.0017101PMID:21359151
49. Steffen SE, Bryant FR. Purification and characterization of the RecA protein from Streptococcus pneu-moniae. Arch Biochem Biophys. 2000; 382: 303–9. doi:10.1006/abbi.2000.2029PMID:11068882
50. Kim J-I, Sharma AK, Abbott SN, Wood EA, Dwyer DW, Jambura A, et al. RecA Protein from the extremely radioresistant bacterium Deinococcus radiodurans: expression, purification, and characteri-zation. J Bacteriol. 2002; 184: 1649–60. doi:10.1128/jb.184.6.1649–1660.2002PMID:11872716
51. Datta S, Krishna R, Ganesh N, Chandra NR, Muniyappa K, Vijayan M. Crystal structures of Mycobacte-rium smegmatis RecA and its nucleotide complexes. J Bacteriol. 2003; 185: 4280–4. doi:10.1128/jb. 185.14.4280–4284.2003PMID:12837805
52. Velankar S, Kleywegt GJ. The Protein Data Bank in Europe (PDBe): bringing structure to biology. Acta Crystallogr D Biol Crystallogr. 2011; 67: 324–30. doi:10.1107/S090744491004117XPMID:21460450
53. Rajan R, Bell CE. Crystal structure of RecA from Deinococcus radiodurans: insights into the structural basis of extreme radioresistance. J Mol Biol. 2004; 344: 951–63. doi:10.1016/j.jmb.2004.09.087PMID:
15544805
54. Zheng H, Chordia MD, Cooper DR, Chruszcz M, Müller P, Sheldrick GM, et al. Validation of metal-bind-ing sites in macromolecular structures with the CheckMyMetal web server. Nat Protoc. 2013; 9: 156–
170. doi:10.1038/nprot.2013.172PMID:24356774
55. Frishman D, Argos P. Knowledge-based protein secondary structure assignment. Proteins. 1995; 23: 566–79. doi:10.1002/prot.340230412PMID:8749853
56. Schrödinger L. The PyMOL Molecular Graphics System, Version 1.3r1. 2010 Aug.
57. Winn MD, Ballard CC, Cowtan KD, Dodson EJ, Emsley P, Evans PR, et al. Overview of the CCP4 suite and current developments. Acta Crystallogr D Biol Crystallogr. 2011; 67: 235–42. doi:10.1107/ S0907444910045749PMID:21460441
58. Chandran A V, Prabu JR, Nautiyal A, Patil KN, Muniyappa K, Vijayan M. Structural studies on Mycobac-terium tuberculosis RecA: molecular plasticity and interspecies variability. J Biosci. 2015; 40: 13–30. doi:10.1007/s12038-014-9497-xPMID:25740138
59. Kelley JA, Knight KL. Allosteric Regulation of RecA Protein Function Is Mediated by Gln194. J Biol Chem. 1997; 272: 25778–25782. doi:10.1074/jbc.272.41.25778PMID:9325305
61. Shatsky M, Nussinov R, Wolfson HJ. A method for simultaneous alignment of multiple protein struc-tures. Proteins. 2004; 56: 143–56. doi:10.1002/prot.10628PMID:15162494
62. Notredame C, Higgins DG, Heringa J. T-Coffee: A novel method for fast and accurate multiple sequence alignment. J Mol Biol. 2000; 302: 205–17. doi:10.1006/jmbi.2000.4042PMID:10964570
63. Robert X, Gouet P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014; 42: W320–4. doi:10.1093/nar/gku316PMID:24753421
64. Krissinel E, Henrick K. Inference of macromolecular assemblies from crystalline state. J Mol Biol. 2007; 372: 774–97. doi:10.1016/j.jmb.2007.05.022PMID:17681537
65. Krissinel E. Macromolecular complexes in crystals and solutions. Acta Crystallogr D Biol Crystallogr. 2011; 67: 376–85. doi:10.1107/S0907444911007232PMID:21460456
66. Lusetti SL, Shaw JJ, Cox MM. Magnesium ion-dependent activation of the RecA protein involves the C terminus. J Biol Chem. 2003; 278: 16381–8. doi:10.1074/jbc.M212916200PMID:12595538
67. Dolinsky TJ, Czodrowski P, Li H, Nielsen JE, Jensen JH, Klebe G, et al. PDB2PQR: expanding and upgrading automated preparation of biomolecular structures for molecular simulations. Nucleic Acids Res. 2007; 35: W522–5. doi:10.1093/nar/gkm276PMID:17488841
68. Dolinsky TJ, Nielsen JE, McCammon JA, Baker NA. PDB2PQR: an automated pipeline for the setup of Poisson-Boltzmann electrostatics calculations. Nucleic Acids Res. 2004; 32: W665–7. doi:10.1093/ nar/gkh381PMID:15215472
69. Baker NA, Sept D, Joseph S, Holst MJ, McCammon JA. Electrostatics of nanosystems: application to microtubules and the ribosome. Proc Natl Acad Sci U S A. 2001; 98: 10037–41. doi:10.1073/pnas. 181342398PMID:11517324
70. Egelman EH, Stasiak A. Structure of helical RecA-DNA complexes. Complexes formed in the presence of ATP-gamma-S or ATP. J Mol Biol. 1986; 191: 677–97. Available:http://www.ncbi.nlm.nih.gov/ pubmed/2949085. PMID:2949085
71. Ellouze C, Takahashi M, Wittung P, Mortensen K, Schnarr M, Norden B. Evidence for Elongation of the Helical Pitch of the RecA Filament Upon ATP and ADP Binding Using Small-Angle Neutron Scattering. Eur J Biochem. 1995; 233: 579–583. doi:10.1111/j.1432-1033.1995.579_2.xPMID:7588804
72. Roy R, Kozlov AG, Lohman TM, Ha T. SSB protein diffusion on single-stranded DNA stimulates RecA filament formation. Nature. Macmillan Publishers Limited. All rights reserved; 2009; 461: 1092–7. doi:
10.1038/nature08442PMID:19820696
73. Baitin DM, Bakhlanova I V, Kil Y V, Cox MM, Lanzov VA. Distinguishing characteristics of hyperrecom-binogenic RecA protein from Pseudomonas aeruginosa acting in Escherichia coli. J Bacteriol. 2006; 188: 5812–20. doi:10.1128/JB.00358-06PMID:16885449
74. Ngo K V, Molzberger ET, Chitteni-Pattu S, Cox MM. Regulation of Deinococcus radiodurans RecA pro-tein function via modulation of active and inactive nucleopropro-tein filament states. J Biol Chem. American Society for Biochemistry and Molecular Biology; 2013; 288: 21351–66. doi:10.1074/jbc.M113.459230
PMID:23729671
75. Radding CM. Homologous pairing and strand exchange in genetic recombination. Annu Rev Genet. Annual Reviews 4139 El Camino Way, P.O. Box 10139, Palo Alto, CA 94303–0139, USA; 1982; 16: 405–37. doi:10.1146/annurev.ge.16.120182.002201PMID:6297377
76. Radding CM, Flory J, Wu A, Kahn R, DasGupta C, Gonda D, et al. Three phases in homologous pairing: polymerization of recA protein on single-stranded DNA, synapsis, and polar strand exchange. Cold Spring Harb Symp Quant Biol. 1983; 47 Pt 2: 821–8. Available:http://www.ncbi.nlm.nih.gov/pubmed/ 6345077.
77. Egner C, Azhderian E, Tsang SS, Radding CM, Chase JW. Effects of various single-stranded-DNA-binding proteins on reactions promoted by RecA protein. J Bacteriol. 1987; 169: 3422–8. Available:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=212412&tool=pmcentrez&rendertype= abstract. PMID:3301800
78. Wang TC, Chang HY, Hung JL. Cosuppression of recF, recR and recO mutations by mutant recA alleles in Escherichia coli cells. Mutat Res. 1993; 294: 157–66. doi:10.1016/0921-8777(93)90024-b
PMID:7687008
79. Madiraju M V, Templin A, Clark AJ. Properties of a mutant recA-encoded protein reveal a possible role for Escherichia coli recF-encoded protein in genetic recombination. Proc Natl Acad Sci U S A. 1988; 85: 6592–6. doi:10.1073/pnas.85.18.6592PMID:2842780
80. Wang TC, Smith KC. recA (Srf) suppression of recF deficiency in the postreplication repair of UV-irradi-ated Escherichia coli K-12. J Bacteriol. 1986; 168: 940–6. Available:http://www.pubmedcentral.nih. gov/articlerender.fcgi?artid=213575&tool=pmcentrez&rendertype=abstract. PMID:3023291
82. Volkert MR, Margossian LJ, Clark AJ. Two-component suppression of recF143 by recA441 in Escheri-chia coli K-12. J Bacteriol. 1984; 160: 702–5. Available:http://www.pubmedcentral.nih.gov/
articlerender.fcgi?artid=214793&tool=pmcentrez&rendertype=abstract. PMID:6094485
83. Yu X, Jacobs SA, West SC, Ogawa T, Egelman EH. Domain structure and dynamics in the helical fila-ments formed by RecA and Rad51 on DNA. Proc Natl Acad Sci U S A. 2001; 98: 8419–24. doi:10. 1073/pnas.111005398PMID:11459984
84. Hewat EA, Ruigrok RW, DiCapua E. Activation of recA protein: the pitch of the helical complex with sin-gle-stranded DNA. EMBO J. 1991; 10: 2695–8. Available:http://www.pubmedcentral.nih.gov/ articlerender.fcgi?artid=452972&tool=pmcentrez&rendertype=abstract. PMID:1868839
85. DiCapua E, Cuillel M, Hewat E, Schnarr M, Timmins PA, Ruigrok RW. Activation of recA protein. The open helix model for LexA cleavage. J Mol Biol. 1992; 226: 707–19. Available:http://www.ncbi.nlm.nih. gov/pubmed/1507222. PMID:1507222
86. Pugh BF, Cox MM. General mechanism for RecA protein binding to duplex DNA. J Mol Biol. 1988; 203: 479–93. doi:10.1016/0022-2836(88)90014-9PMID:3058986
87. Wu Y, He Y, Moya IA, Qian X, Luo Y. Crystal structure of archaeal recombinase RADA: a snapshot of its extended conformation. Mol Cell. 2004; 15: 423–35. doi:10.1016/j.molcel.2004.07.014PMID:
15304222
88. Lee M-H, Chang Y-C, Hong EL, Grubb J, Chang C-S, Bishop DK, et al. Calcium ion promotes yeast Dmc1 activity via formation of long and fine helical filaments with single-stranded DNA. J Biol Chem. 2005; 280: 40980–4. doi:10.1074/jbc.M505896200PMID:16204247
89. Bork JM. The RecOR proteins modulate RecA protein function at 5’ends of single-stranded DNA. EMBO J. 2001; 20: 7313–7322. doi:10.1093/emboj/20.24.7313PMID:11743007
90. Inoue J, Nagae T, Mishima M, Ito Y, Shibata T, Mikawa T. A mechanism for single-stranded DNA-bind-ing protein (SSB) displacement from sDNA-bind-ingle-stranded DNA upon SSB-RecO interaction. J Biol Chem. 2011; 286: 6720–32. doi:10.1074/jbc.M110.164210PMID:21169364
91. Mayola A, Irazoki O, Martínez IA, Petrov D, Menolascina F, Stocker R, et al. RecA Protein Plays a Role in the Chemotactic Response and Chemoreceptor Clustering of Salmonella enterica. Cloeckaert A, edi-tor. PLoS One. Public Library of Science; 2014; 9: e105578. doi:10.1371/journal.pone.0105578PMID:
25147953
92. Gómez-Gómez J-M, Manfredi C, Alonso J-C, Blázquez J, Harshey R, Soutourina O, et al. A novel role for RecA under non-stress: promotion of swarming motility in Escherichia coli K-12. BMC Biol. BioMed Central; 2007; 5: 14. doi:10.1186/1741-7007-5-14PMID:17391508
93. Burkart M, Toguchi A, Harshey RM. The chemotaxis system, but not chemotaxis, is essential for swarming motility in Escherichia coli. Proc Natl Acad Sci U S A. 1998; 95: 2568–73. Available:http:// www.ncbi.nlm.nih.gov/pubmed/9482927. PMID:9482927
94. Irazoki O, Mayola A, Campoy S, Barbé J, Henrichsen J, Butler M, et al. SOS System Induction Inhibits the Assembly of Chemoreceptor Signaling Clusters in Salmonella enterica. Rao C V., editor. PLoS One. Public Library of Science; 2016; 11: e0146685. doi:10.1371/journal.pone.0146685PMID: