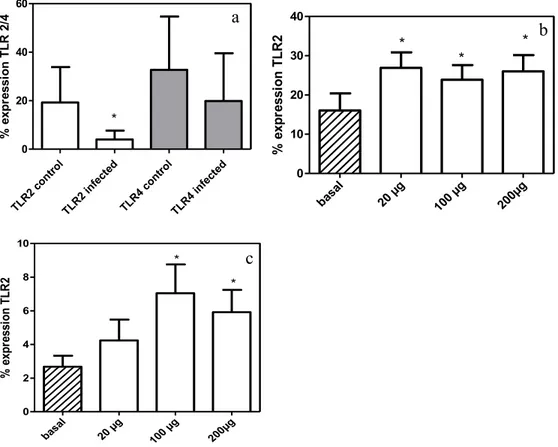
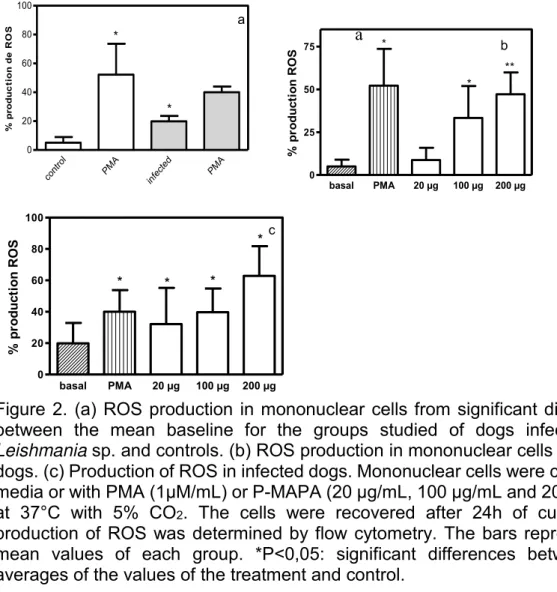
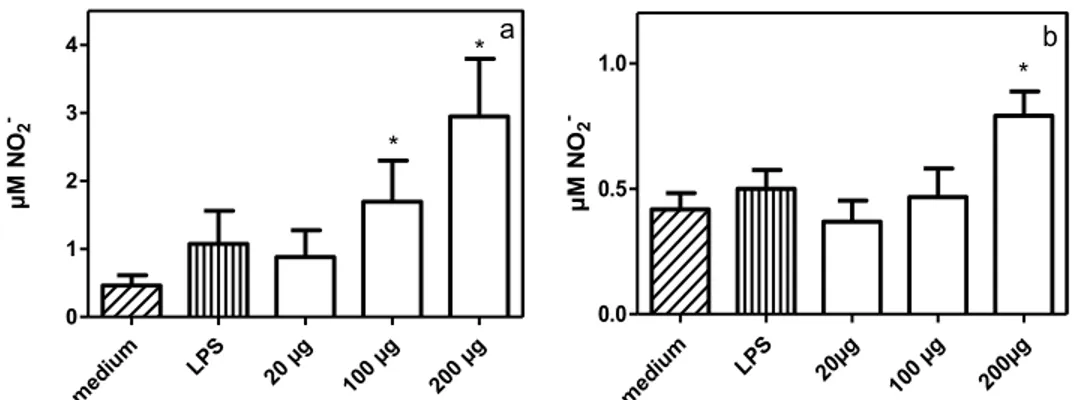
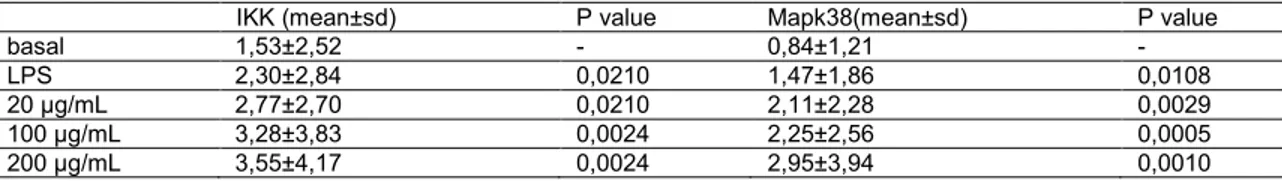
TLR 2, TLR 4, ROS, óxido nítrico, P38 e IKK em células mononucleares de cães com leishmaniose Visceral tratadas com o imunomodulador P-MAPA: Larissa Martins Melo. -
Texto
Imagem
Outline
Documentos relacionados
Animals or coelomocytes were exposed to different salinity (25‰ to 45‰) and phagocytic parameters , production of reactive oxygen species (ROS), mitochondrial activity and
We further studied the possible association of key signaling kinases (protein kinase C, tyrosine kinases, phosphoinositide 3-kinase, and Rho kinase) and reactive oxygen species
Um relatório de sustentabilidade fornece uma declaração equilibrada e razoável do desempenho de sustentabilidade da organização, incluindo contribuições positivas e
A presente dissertação tem por objetivo o estudo da implementação das decisões da Corte Interamericana de Direitos Humanos no âmbito do Direito brasileiro. A
GLUT4 translocation precedes the stimulation of glucose uptake by insulin in muscle cells: potential activation of GLUT4 via p38 mitogen-activated protein
TAO (thousand-and-one amino acid) protein kinases mediate signaling from carbachol to p38 mitogen-activated protein kinase and ternary complex factors. Cleavage orientation and the
Considerando que as feiras de arte e artesanato são definidas segundo a UNCTAD (2010) como serviços criativos, de ordem cultural e recreativa, observa-se que a
It is now becoming clear that ROS play an important role in the acute inflammatory process after intense exercise [45] and that the skeletal muscle inflammatory process
