INSILICO SCREENING OF NNRTI AS
POTENTIAL TARGETS FOR
TUMOR CONTROL
DEVADASS CHANDRAMOHAN*
Department of Molecular Biosciences,Bishop Heber College, (Affiliated to Bharathidasan University, Tiruchirappalli) Post Box No. 615, Tiruchirappalli - 620 017, Tamil Nadu, India
KANDASAMY MUTHUKRISHNAKUMAR
^
Department of Biotechnology,Bishop Heber College, (Affiliated to Bharathidasan University, Tiruchirappalli) Post Box No. 615, Tiruchirappalli - 620 017, Tamil Nadu, India
CYNTHIA CHINNAPPARAJ
Assistant Lecturer, Department of Biotechnology, Bishop Heber College, (Affiliated to Bharathidasan University, Tiruchirappalli)
Post Box No. 615, Tiruchirappalli - 620 017, Tamil Nadu, India
MARUDHAMUTHU RAJADURAI#
Assistant Lecturer, Department of Biotechnology and Bioinformatics, Bishop Heber College, (Affiliated to Bharathidasan University, Tiruchirappalli)
Post Box No. 615, Tiruchirappalli - 620 017, Tamil Nadu, India *^ These authors contributed equally to this work
#
Corresponding author
Abstract
Nevirapine, a NNRTI was proven effective in differentiation of transformed cells by arresting the reverse transcriptase activity of telomerase in transformed cells. According to a recent hypothesis, almost all NNRTI functions by a similar mechanism against 1 RT. The presence of evolutionary conserved regions between HIV-1 RT and hTERT, a catalytic subunit of telomerase also implies the importance of those regions in reverse transcriptase activity. Such evolutionary conserved regions and Nevirapine’s activity against cancer cell proliferation will make obvious that NNRTI which are used as anti-HIV drugs could also be used as to target telomerase reverse transcriptase in cancer cells. hTERT is composed of four domains which are known as N-Terminal, RNA binding domain, RT domain, and C-Terminal domain. It is possible to find out the best NNRTI against cancer cell control by docking all available NNRTI against this RT domain of hTERT. Due to the lack of crystallographic and NMR structure of this hTERT, the domain docking approach was handled to overcome structure deficit. This study focused on available NNRTI, to find out the better NNRTI to use as anti-cancer leads or precursor compounds.
Keywords: NNRTI, telomerase, hTERT, RT, Docking, Interactions
1. Introduction
in germ line cells, immortal cells, and most cancer cells but is inactive in normal somatic cells. This specific feature projects telomerase as an attractive and novel target for cancer therapeutics that might have minimal side effects as mentioned by Gellert et al. (2005). Literature evidences substantiate that telomerase reverse transcriptase is similar to conventional reverse transcriptase (Fig. 1).
Fig. 1. Similarity between HIV-1 RT and hTERT with motif alignments
In particular, hTERT exhibit similarities with the functionally important amino acid residues of HIV-1 RT which was studied by Lundblad (1998). Cech et al. (1997) in their review stated that the similarities are obvious, especially among the evolutionarily conserved regions of telomerase reverse transcriptase and HIV-1 RT. Due to the similarity between both reverse transcriptases, researchers evoke thoughts of using non-nucleoside reverse transcriptase inhibitors (NNRTIs) inhibitors to target telomerase reverse transcriptase.
Non-nucleoside reverse transcriptase inhibitors (NNRTIs) were identified as a new class of antiretroviral drugs over the past twenty years by means of dedicated research and development towards HIV treatment, which were explained well by Bethune (2010). These NNRTIs are diverse in their chemical structure as referred by Prajapati et al. (2009). Cremer et al. (2004) explained that these NNRTIs are unique in their inhibitory mechanism in targeting HIV reverse transcriptase enzyme by binding between the “fingers” and “thumb” subdomains of the p66 subunit. Recently, Ivetac and McCammon (2009) proposed a hypothesis of reverse transcriptase inhibition by nevirapine. According to their study, the movement or sliding of two β-sheets connected by a hinge in the p66 domain of hTERT are essential for the reverse transcriptase activity. During nevirapine treatment, this NNRTI binds in between these two β-sheets and block the hinge region, resulting in reverse transcriptase inhibition. Studies on X-ray crystal structures of HIV-1 RT in complex with NNRTI also evidenced that most of the NNRTI has similarities in their geometry of the bound inhibitor as supported by Cremer et al. (2004)
Mangiacasale et al. (2003) observed that nevirapine, a first generation NNRTI induced differentiation of transformed cells, after certain period of exposure to nevirapine treatment. Indeed, it’s a surprising result that the mechanism is due to the inhibition of reverse transcriptase activity as seen in HIV-1 RT. Inhibition of cancer cell’s proliferation by NNRTI and striking similarity between HIV-RT and hTERT hypothesized for docking studies of NNRTIs to target RT domain of hTERT with the ultimate goal of finding comparatively better NNRTI against telomerase inhibition. In this insilico study based on dock score and interactions with RT domain, some NNRTI’s seem to be presumably better than nevirapine against telomerase inhibition.
2. Materials and Methods
The amino acid sequence of human telomerase enzyme (Accession number: AAC51672.1) was obtained from National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov), which provides access to biomedical and genomic information (Wheeler et al. (2007)).
Based on the hypothesis that nevirapine inhibits reverse transcriptase activity of telomerase, RT domain of hTERT was considered as the docking target. Due to the deficit of crystallographic structure of RT domain, it was modelled using protein structure prediction server SAM-T08 (http://compbio.soe.ucsc.edu/SAM_T08/T08-query.html), an iterative hidden Markov model based method (Karplus (2009)). The server has been validated as part of the CASP8 assessment of structure prediction with good performance across all classes of predictions.
Three dimensional structures of 27 NNRTIs were derived from NCBI-PubChem Compound database (http://pubchem.ncbi.nlm.nih.gov/), which is a public repository for biological properties of small molecules hosted by the US National Institutes of Health (also see supplementary 1). Retrieved compound structures were docked against modelled RT domain. The docking was performed with the molecular docking algorithm MolDock using Molegro Virtual Docker (MVD) (http://www.molegro.com) according to instructions given in the MVD tutorial. Molegro Virtual Docker is an integrated platform for predicting protein–ligand interactions (Yu et al. (2009)).
Protein Data Bank (PDB), is an archive for biological macromolecular crystal structures (Berman (2000)). Three dimensional structure of HIV-1 Reverse transcriptase bound with nevirapine complex was derived from Protein Data Bank (PDB) (http://www.rcsb.org/pdb/home/home.do) in .pdb format. The interactions of the nevirapine and its interacting residues within the binding site of hTERT and HIV-1 RT were analyzed using Ligand Explorer (http://ligpro.sdsc.edu), which is integrated with protein data bank. This Ligand Explorer is programmed well, to provide details of hydrophilic and hydrophobic interactions between a protein and a ligand at different cutoff distances (Moreland et al. (2005)). Based on the dock score and hydrophilic interactions, a few candidate NNRTIs are selected as final set of candidates.
clogP values were obtained for the final set of candidates by drawing them in Osiris Property Explorer (http://www.organic-chemistry.org), a basic toxicity prediction tool with integrated facility for molecular weight and cLogP prediction (Malik et al. (2007)). This cLogP prediction is important in drug discovery, because cLogP<5.0, is an indication for a good drug properties, according to Lipinski’s rule of Five (Lipinski et al. (2001)).
3. Results
3.1. Fragmentation of hTERT amino acid sequence into domains
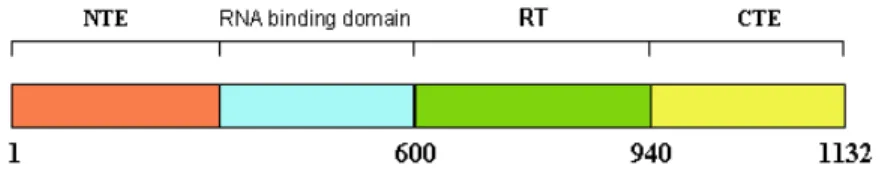
The amino acid sequence of human telomerase enzyme was obtained from NCBI (Accession number: AAC51672.1) which consists of 1132 amino acids (Fig. 2).
The sequence is considered for modeling due to the absence of its three dimensional structure in protein structural databases. Modelling of lengthy amino acid chain proteins (1132 aminoacids) like hTERT is very complex and the resulting structure may not be reliable. To overcome this barrier the entire amino acid sequence was fragmented into four domains (Fig. 3) namely N terminal and RNA binding domain (1aa - 600aa), RT domain (604aa – 939aa), and C Terminal domain (940aa – 1132aa) with reference to telomerase database.
Fig. 3. Diagrammatic representation of Domains in telomerase reverse transcriptase
3.2. Modeling of telomerase RT domain
Reverse transcriptase (RT) domain is highly conserved in both telomerase and HIV-1 RT. In addition, the amino acid sequence corresponding to the binding site of nevirapine in HIV-1 RT is found to be similar to RT domain of hTERT. Due to this observation, RT domain is solely considered for modeling and docking studies. Based on the domain segregation from telomerase database, the amino acid sequences between 604aa to 939aa was considered as RT domain and modelled (Fig. 4) using HMM based SAM T08 server [http://compbio.soe.ucsc.edu/SAM_T08/T08-query.html].
Fig. 4. Ribbon representation of modeled RT domain of hTERT
3.3. Interactions of Nevirapine with HIV-1 RT and TERT
Fig. 5. Bound Nevirapine (green) in RT domain
Binding pocket of nevirapine was traced out in both HIV-1 RT and RT domain of hTERT using Ligand scout (Fig. 6). The results of ligand scout revealed that the binding site of nevirapine in RT domain of hTERT was similar to the binding site of nevirapine in HIV-1 RT, but with slightly distinguished variations. The interactions of nevirapine with both hTERT and HIV-1 RT were analyzed by Ligand Explorer. The results of this interactions study, depicted considerable similarities among their interacting residues, with highly similar residues in their binding region as shown below.
Residues in binding region of HIV-1 RT
Region 1 (226-236): PFLWMGYELHP
Region 2 (176-192): PDIVIYQYMDDLYVGSD
Residues in binding region of RT domain of hTERT
Region 1 (236 – 249): LLCSLCYGDMENKL Region 2 (257 – 271): GLLLRLVDDFLLVTP
Fig. 6. Comparison of Nevirapine binding site in both HIV-1 RT (a) and RT domain of hTERT (b) Nevirapine interacting regions (Region 1 and 2) are marked as circles
responsible for HIV-1 reverse transcriptase activity. The telomerase inhibition by nevirapine may also be due to the contact inhibition arrest of these two regions of hTERT RT domain by nevirapine binding in between them.
3.4. Docking and interaction studies
NNRTIs were successfully docked against RT domain of hTERT using Molegro Virtual Docker and their dock scores were recorded (Table 1).
Table 1. Mol dock score of NNRTIS docked against RT domain of hTERT
S. No NNRTIs Molecular weight Dock Score
1 TIBO derivatives [i.e., 8-chloro-TIBO (R86183, Tivirapine)] 321.868 -104.262 2 Dipyridodiazepinones [Nevirapine (BIRG-587)] 266.298 -80.4485
3 Pyridinones (L-697,661) 352.215 -109.176
4 BHAP derivatives [Delavirdine – (U-90152)] 456.561 -125.486
5 a-APA derivatives [Loviride] 351.227 -107.171
6 PETT derivatives [Trovirdine ] 337.238 -108.308
7 Thiocarboxanilide derivatives (UC-781) 335.848 -111.044
8 Thiazolobenzimidazole (NSC 625487) 288.315 -85.1978
9 Benzothiadiazine NSC 287474 274.361 -87.3746
10 Quinazolinones 323.776 -105.832
11 Benzoxazinones (Efavirenz) 315.675 -102.678
12 Calanolide A 370.439 -83.834
13 Imidazodipyridodiazepine UK-129,485 307.35 -91.4821
14 Imidazopyridazines 387.478 -132.2
15 Thiadiazolyl Dialkylcarbamates (TDA RD-4-2024) 346.232 -105.046 16 Arylpyridodiazepine and –Thiodiazepine derivatives (MEN
10979) 326.439 -84.6699
17 Alkoxy(arylthio)uracils 348.46 -99.8748
18 Indolyl Dipyridodiazepinones 369.419 -97.7664
19 PNU – 32945 273.332 -109.103
20 4-methyl MBI 370.343 -101.795
21 Etravirine TMC 125 435.277 -114.865
22 Rilpivirine 366.419 -114.119
23 RDEA806 557.847 -143.067
24 Lersivirine (UK-453061) 310.35 -129.186
25 Dapivirine 329.398 -105.909
26 Thiazoloisoindolinones 295.399 -97.846
27 Pyrrolobenzoxazepinones 303.354 -80.4397
Table 2. Interacting residues and number of hydrogen bonds formation by NNRTI with RT domain of hTERT
S. No Ligands Interacting Residues (Hydrophilic)
No. of Hydrogen
Bonds
1
TIBO derivatives [i.e., 8-chloro-TIBO (R86183, Tivirapine)]
SER234,LEU237,LEU233 9 2 Dipyridodiazepinones
[Nevirapine (BIRG-587)] SER239, LEU237,LEU240, LEU262, VAL263 10 3 Pyridinones (L-697,661) CYS238,SER234,CYS238,SER239,THR235,VAL263 10 4 BHAP derivatives
[Delavirdine - (U-90152)]
ARG139,ALA75,ASP244,GLY243,THR235,LEU236, LEU237,ARG139,LEU240,LEU236,SER239,LEU233, SER234,THR235
29 5 a-APA derivatives [Loviride] CYS238,LEU240,ALA75, ASP244, LEU237 10 6 PETT derivatives [Trovirdine
] LEU237,ARG139, LEU236 5
7 Thiocarboxanilide derivatives
(UC-781) SER231,ILE232, SER234, PRO228 6
8 Thiazolobenzimidazole
(NSC 625487) THR235,CYS238 2
9 Benzothiadiazine NSC
287474 LEU233,ARG139 5
10 Quinazolinones ARG139,LEU240,SER239 7
11 Benzoxazinones (Efavirenz) ARG139, TYR140 2
12 Calanolide A TYR114,CYS238,SER239,THR235,LEU236 8
13 Imidazodipyridodiazepine
UK-129,485 LEU233,ARG139,ILE232,LEU236 6
14 Imidazopyridazines ARG139,LEU240,SER239 7
15
Thiadiazolyl Dialkylcarbamates (TDA RD-4-2024)
SER239,CYS241,SER234,LEU236, LEU237,TYR242 14
16
Arylpyridodiazepine and – Thiodiazepine derivatives (MEN 10979)
TYR242, SER234 6
17 Alkoxy(arylthio)uracils SER239,ARG139 5
18 Indolyl Dipyridodiazepinones CYS241,LEU240,LEU236,SER239,ALA75,SER76,
LEU240 11
19 PNU – 32945 CYS238,LEU237, LEU240, CYS241, TYR242 6
20 4-methyl MBI CYS238,CYS241 3
21 Etravirine TMC 125 CYS241,TYR242,GLY243,ASP244,CYS238,LEU240,
LEU237 12
22 Rilpivirine TYR242,GLY243,ASP244,SER239,LEU240,LEU236,
CYS241, LEU237, LYS248,TYR289 14
23 RDEA806 TYR114, ARG28, GLN229, THR235, TYR114 5
24 Lersivirine (UK-453061) CYS238,CYS238, LEU240, PRO118, LEU237,
GLY243 10
25 Dapivirine CYS241,TYR242,GLY243,ASP244,CYS238,LEU240,
LEU237 12
26 Thiazoloisoindolinones THR235, LEU236 2
Finally the dock score and number of hydrogen bonds formed by nevirapine with RT domain was kept as a cut off value for screening out the candidates. NNRTIs excelled above these fixed cut off values were tabulated (Table 3) and considered as final set of compounds better than nevirapine to target telomerase inhibition.
Table 3. List of NNRTIs showing better dock score and number of hydrogen bond interactions with RT domain of hTERT
S. No Ligands No. of
hydrogen bonds Dock Score cLogP
1 BHAP derivatives [Delavirdine - (U-90152)] 29 -125.49 1.95
2 Rilpivirine 14 -114.12 6.01
3 Etravirine TMC 125 12 -114.87 5.26
4 a-APA derivatives [Loviride] 10 -107.17 2.64
5 Pyridinones (L-697,661) 10 -109.18 3.44
6 Lersivirine (UK-453061) 10 -129.19 2.14
aCut off values by Nevirapine: (i) Dock score : -80.449, (ii) Number of Hydrogen bond interactions : 10
All these candidates of final set were analyzed for their cLogP values. In this step, 2 compounds (Rilpivirine, and Etravirine TMC 125) got eliminated for their excess cLogP value. The remaining 4 compounds obey the Lipinski’s range for a better drug ie) ClogP<5.0, according to Lipinski’s rule of Five (Table 4).
Table 4. Candidate NNRTI compounds to target RT domain of hTERT
S. No Ligands No. of
hydrogen bonds Dock Score cLogP
1 BHAP derivatives [Delavirdine - (U-90152)] 29 -125.49 1.95
2 a-APA derivatives [Loviride] 10 -107.17 2.64
3 Pyridinones (L-697,661) 10 -109.18 3.44
4 Lersivirine (UK-453061) 10 -129.19 2.14
These four compounds are found to be better compounds to use as initial leads for anti-cancer drug discovery research. These compounds will act as better leads to inhibit excessive telomerase activity of cancer cells with high chance of inhibition over RT domain of hTERT.
4. Discussion
5. Conclusion
Our objective in this docking study is to find out the better NNRTI that could be used as anti-cancer drugs in addition to Nevirapine. From our results, we conclude that the best scored inhibitors will have the better chance of inhibiting cancer cells by targeting telomerase enzyme as well as to use as anti-cancer leads. It is also possible in forth coming future to develop these NNRTI, to be suitable for cancer therapeutics. This goal can be achieved with further research attention over the functional residues lying in the NNRTI binding site of telomerase. Due to the immediate need of better drugs for mostly prevalent problems like cancer, it is essential to screen compound libraries computationally, to speed up the drug discovery process. Such a computational screening and comparative docking of compounds or leads with proven drugs will lead to the identification of new efficient drugs. Awaiting for crystal structures, which are very complex to solve for high molecular weight and large proteins will delay the drug discovery process. For such complex proteins, one can go forward with domain docking, by promoting identification of functionally important regions related to their studies.
References
[1] Berman, H. M.; Westbrook, J.; Feng, Z.; Gillil, G.; Bhat, T. N.; Weissig, H.; Shindyalov, I. N.; Bourne, P. E. (2000): The Protein data bank. Nucleic Acids Res., 28(1), pp. 235-242.
[2] Bethune, M. P. (2010): Non-nucleoside reverse transcriptase inhibitors (NNRTIs), their discovery, development, and use in the treatment of HIV-1 infection: A review of the last 20 years (1989–2009). Antiviral Res., 85(1), pp. 75–90.
[3] Cech, T. R.; Nakamura, T. M.; Lingner, J. (1997): Telomerase is a true reverse transcriptase. A review. Biochemistry (Mosc), 62(11), pp. 1202-1205.
[4] Cremer, N. S.; Temiz, N. A.; Bahar, I. (2004): Conformational Changes in HIV-1 Reverse Transcriptase Induced by non nucleoside Reverse Transcriptase Inhibitor Binding. Curr. HIV Res., 2(4), pp. 323–332.
[5] Gellert, G. C.; Jackson, S. R.; Dikmen, Z. G.; Wright, W. E.; Shay, J. W. (2005): Telomerase as a therapeutic target in cancer. Drug Discovery Today: Disease Mechanisms, 2(2), pp. 159-164.
[6] Ivetac, A.; McCammon, J. A. (2009): Elucidating the inhibition mechanism of HIV-1 non-nucleoside reverse transcriptase inhibitors through multi-copy molecular dynamics simulations. J. Mol. Biol., 388(3), pp. 644–658.
[7] Karplus, K. (2009): SAM-T08, HMM-based protein structure prediction. Nucleic Acids Res., 37, pp. W492–W497.
[8] Kirkpatrick, K. L.; Mokbel, K. (2001): The significance of human telomerase reverse transcriptase (hTERT) in cancer. Eur. J. Surg. Oncol., 27(8), pp. 754–760.
[9] Lipinski, C. A.; Lombardo, F.; Dominy, B. W.; Feeney, P. J. (2001): Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug. Deliv. Rev., 46, pp. 3-26.
[10] Lundblad, V. (1998): Telomerase catalysis: A phylogenetically conserved reverse transcriptase. Proc. Natl. Acad. Sci. U S A, 95(15), pp. 8415–8416.
[11] Malik,I.; Sedlarova, E.; Csollei, J.; Andriamainty,F.; Cizmarik,J. (2007): Relationship between physicochemical properties, lipophilicity parameters, and local anesthetic activity of dibasic esters of phenylcarbamic acid. Chem. Pap., 61(3), pp. 206-213.
[12] Mangiacasale, R.; Pittoggi, C.; Sciamanna, I.; Careddu, A.; Mattei, E.; Lorenzini, R.; Lorena; Travaglini, L.; Landriscina, M.; Barone, C.; Nervi, C.; Lavia, P.; Spadafora, C. (2003): Exposure of normal and transformed cells to nevirapine, a reverse transcriptase inhibitor, reduces cell growth and promotes differentiation. Oncogene, 22(18), pp. 2750-2761.
[13] Mokbel, K. (2003): The evolving role of telomerase inhibitors in the treatment of cancer. Curr. Med. Res. Opin., 19(6), pp. 470–472. [14] Moreland, J. L.; Gramada, A.; Buzko, O. V.; Zhang, Q.; Bourne, P. E. (2005): The Molecular Biology Toolkit (MBT): a modular platform
for developing molecular visualization applications. BMC Bioinformatics, 6, pp. 21.
[15] Podlevsky, J. D.; Bley, C. J.; Omana, R. V.; Xiaodong, Q.; Chen, J. L. (2008): The telomerase database. Nucleic Acids Res., 36, pp. D339– D343.
[16] Prajapati, D. G.; Ramajayam, R.; Yadav, M. R.; Giridhar, R. (2009): The search for potent, small molecule NNRTIs: A review. Bioorg Med Chem., 17(16), pp. 5744–5762.
[17] Wheeler, D. L.; Barrett, T.; Benson, D. A.; Bryant, S. H.; Canese, K.; Chetvernin, V.; Church, D. M.; Dicuccio, M.; Edgar, R.; Federhen, S.; Geer, L. Y.; Kapustin, Y.; Khovayko, O.; Landsman, D.; Lipman, D. J.; Madden, T. L.; Maglott, D. R.; Ostell, J.; Miller, V.; Pruitt, K. D.; Schuler, G. D.; Sequeira, E.; Sherry, S. T.; Sirotkin, K.; Souvorov, A.; Starchenko, G.; Tatusov, R. L.; Tatusova, T. A.; Wagner, L.; Yaschenko E. (2007): Database resources of the National Center for Biotechnology Information. Nucleic Acids Res., 35, pp. D5-D12. [18] Yu, H. N.; Zhang, L. C.; Yang, J. G.; Das, U. N.; Shen, S. R. (2009): Effect of laminin tyrosine-isoleucine-glycine-serine-arginine peptide
on the growth of human prostate cancer (PC-3) cells in vitro. Eur. J. Pharmacol., 616(1-3), pp. 251-255. [19] Ligand Explorer (LigPro) Home page [http://ligpro.sdsc.edu]
[23] Protein Data Bank [http://www.rcsb.org/pdb/home/home.do]
Insilico screening of NNRTI as potential targets for tumor control – Chandramohan et al. Nevirapine CID 4463
Tivirapine CID 3000323
Pyridinones (L-697,661) CID 65002
Delavirdine CID 5625
Loviride CID 3963
Trovirdine CID 3000870
2D and 3D structures of the 27 candidate non-nucleosidic reverse transcriptase inhibitors
Insilico screening of NNRTI as potential targets for tumor control – Chandramohan et al. Benzothiadazine CID 3000415
Quinazolinone CID 72417
Efavirenz CID 64139
Calanolide A CID 64972
Imidazopyridazines CID 189892
Thiadiazolyl Dialkylcarbamates CID 463219
MEN 10979 CID 462243
Insilico screening of NNRTI as potential targets for tumor control – Chandramohan et al. Indolyl Dipyridodiazepinones CID 466843
Etravirine CID 193962
Dapivirine CID 214347
4-methyl MBI CID 468848
PNU – 32945 CID 477596
Rilpivirine CID 6451164
Pyrrolobenzoxazepinones CID 385306
Insilico screening of NNRTI as potential targets for tumor control – Chandramohan et al. RDEA 806 CID 16221816
Thiocarboxanilide derivatives (UC-781) CID 3000926
Imidazodipyridodiazepine UK-129,485 CID 476263
Thiazoloisoindolinones CID 196920
Insilico screening of NNRTI as potential targets for tumor control – Chandramohan et al.
Candidate non-nucleosidic reverse transcriptase inhibitors with their PubChem Compound ID
S. No Ligands PubChem CID
1 Dipyridodiazepinones [Nevirapine (BIRG-587)] CID 4463
2 TIBO derivatives [i.e., 8-chloro-TIBO (R86183, Tivirapine)] CID 3000323
3 Pyridinones (L-697,661) CID 65002
4 BHAP derivatives [Delavirdine - (U-90152)] CID 5625
5 a-APA derivatives [Loviride] CID 3963
6 PETT derivatives [Trovirdine ] CID 3000870
7 Benzothiadiazine NSC 287474 CID 3000415
8 Quinazolinones CID 72417
9 Benzoxazinones (Efavirenz) CID 64139
10 Calanolide A CID 64972
11 Imidazopyridazines CID 189892
12 Thiadiazolyl Dialkylcarbamates (TDA RD-4-2024) CID 463219
13 Arylpyridodiazepine and –Thiodiazepine derivatives (MEN
10979) CID 462243
14 Alkoxy(arylthio)uracils CID 466628
15 Indolyl Dipyridodiazepinones CID 466843
16 Etravirine TMC 125 CID 193962
17 Dapivirine CID 214347
18 4-methyl MBI CID 468848
19 PNU – 32945 CID 477596
20 Rilpivirine CID 6451164
21 Pyrrolobenzoxazepinones CID 385306
22 Lersivirine CID 16739244
23 RDEA806 CID 16221816
24 Thiocarboxanilide derivatives (UC-781) CID 3000926
25 Thiazolobenzimidazole (NSC 625487) CID 122665
26 Imidazodipyridodiazepine UK-129,485 CID 476263