ContentslistsavailableatScienceDirect
Neuroscience
Letters
j ou rn a l h om ep a g e :w w w . e l s e v i e r . c o m / l o c a t e / n e u l e t
Research
paper
Effect
of
FGF-2
and
sciatic
nerve
grafting
on
ChAT
expression
in
dorsal
root
ganglia
neurons
of
spinal
cord
transected
rats
Fausto
Pierdoná
Guzen
a,∗,
Dayane
Pessoa
de
Araújo
a,
Eudes
Euler
de
Souza
Lucena
a,
Hécio
Henrique
Araújo
de
Morais
a,
José
Rodolfo
Lopes
de
Paiva
Cavalcanti
a,
Expedito
Silva
do
Nascimento
Jr.
b,
Miriam
Stela
Maris
de
Oliveira
Costa
b,
Jeferson
Sousa
Cavalcante
caLaboratoryofExperimentalNeurology,HealthScienceCenter,StateUniversityofRioGrandedoNorte,Mossoró,RN,Brazil bLaboratoryofNeuroanatomy,DepartmentofMorphology,FederalUniversityofRioGrandedoNorte,Natal,RN,Brazil cLaboratoryofNeurochemicalStudies,DepartmentofPhysiology,FederalUniversityofRioGrandedoNorte,Natal,RN,Brazil
h
i
g
h
l
i
g
h
t
s
•TransectedspinalcordasamodelforstudyofDRGregeneration.
•PeripheralnervegraftsasafavorableenvironmenttoDRGneuroprotection.
•FGF-2potentiatesneuroprotectiveeffectinDRGafterspinalcordinjury.
•FGF-2plussciaticnervefragmentimproveDRGplasticityofratssubmittedtocompletetransectionsofspinalcord.
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received24June2015
Receivedinrevisedform17August2015 Accepted23August2015
Availableonline28August2015
Keywords:
Dorsalrootganglia Fibroblasticgrowthfactor-2 Neuroprotection
Sciaticnervegraft Spinalcord
a
b
s
t
r
a
c
t
NeurotrophicfactorsandperipheralnervesareknowntobegoodsubstratesforbridgingCNStrauma. Theinvolvementoffibroblastgrowthfactor-2(FGF-2)activationinthedorsalrootganglion(DRG)was examinedfollowingspinalcordinjuryintherat.WeevaluatedwhetherFGF-2increasestheabilityof asciaticnervegrafttoenhanceneuronalplasticity,inagappromotedbycompletetransectionofthe spinalcord.Theratsweresubjectedtoa4mm-longgapatlowthoraciclevelandwererepairedwith saline(Salineorcontrolgroup,n=10),orfragmentofthesciaticnerve(Nervegroup,n=10),orfragment ofthesciaticnervetowhichFGF-2(Nerve+FGF-2group,n=10)hadbeenaddedimmediatelyafterlesion. TheeffectsoftheFGF-2andfragmentofthesciaticnervegraftsonneuronalplasticitywereinvestigated usingcholineacetyltransferase(ChAT)-immunoreactivityofneuronsinthedorsalrootganglionafter8 weeks.Preservationoftheareaanddiameterofneuronalcellbodiesindorsalrootganglion(DRG)was seeninanimalstreatedwiththesciaticnerve,aneffectenhancedbytheadditionofFGF-2.Thus,the additionofexogenousFGF-2toasciaticnervefragmentgraftedinagapoftheratspinalcordsubmitted tocompletetransectionwasabletoimproveneuroprotectionintheDRG.Theresultsemphasizedthat themanipulationofthemicroenvironmentinthewoundmightamplifytheregenerativecapacityof peripheralneurons.
©2016PublishedbyElsevierIrelandLtd.
1. Introduction
Trophicfactors,suchasnervegrowthfactor(NGF),brainderived neurotrophicfactor(BDNF)andglia-derivedneurotrophicfactor
∗ Correspondingauthorat:DepartmentofBiomedicSciences,HealthSciences Faculty,StateUniversityofRioGrandedoNorte,Mossoró59607-360,RioGrande doNorte,Brazil.
E-mailaddress:[email protected](F.P.Guzen).
(GDNF)showsimportantandselectiveeffectsonsurvivaland phe-notypicexpressionofprimarysensoryneuronsinthedorsalroot ganglion (DRG)followingnervous systeminjury and peripheral inflammationTrophicfactors,suchasnervegrowthfactor(NGF), brainderived neurotrophic factor(BDNF) andglia-derived neu-rotrophicfactor(GDNF)showimportantandselectiveeffectson survivalandphenotypicexpressionofprimarysensoryneuronsin thedorsalrootganglion(DRG)followingnervoussysteminjuryand peripheralinflammation[1–4].
http://dx.doi.org/10.1016/j.neulet.2015.08.043
positivesubpopulationofsensoryneurons[6].
Evidencefromseveralinvitroandinvivostudieshasshown that FGF-2is not only present in the nervoussystem but also mediatessurvival-promotingeffectsandstimulatesthe transmit-termetabolismofseveralneuronsduringdevelopmentandafter injury[15,16].ApplicationofFGF-2totheproximalstumpofthe transectionedsciaticnervepreventsthelesion-induceddeathof sensoryneuronsofDRGL4-L6[12].
Animal studies have shown that if continuity is restored betweenthespinal cordand theventralroots ofnerves atthe lumbar[17] orcervical[18–20]levelsofthespinalcord, motor neuronaxonscanregrowintotheirrespectiveperipheralnerves withconcomitantrecoveryofmotorfunctions.Thereturnofmotor functionsafterimplantationofavulsedspinalnerverootsintothe spinalcordhasalsobeenreportedinoneclinicalcase[21],aswell asinsomeanimalstudies[22–24].
FGF-1and FGF-2treated cultures can promote a significant increaseinneuriteoutgrowthofventralspinalcordneuronsand stemcells,suggestingthatbothFGFscaninfluenceneuronal devel-opment[25,26].
InordertodeterminefunctionalrolesofFGF-2inthe periph-eralnervoussystemweanalyzedtheexpressionofcholineacetyl transferase(ChAT)inspinalgangliaafterspinalcordlesiontreated withFGF-2.
2. Materialsandmethods
2.1. Animaltreatment
AdultmaleWistarrats(n=30)fromtheFederalUniversityof Pernambuco,Brazil(bodyweight[b.w.]180–200g),wereusedin thepresentstudy.Thestudywasconductedaccordingto proto-colsapprovedbytheAnimalCareandUseEthicCommitteeatthe FederalUniversityof Rio GrandedoNorte usingtheSanPoiley outbreedingmethod[27].
2.2. Microneurosurgery
Ratswerepre-anaesthetizedwithanintramuscularinjectionof ketaminechloridrate10%(AgenerUnião,Brazil,0,1ml/100gb.w.) andxylazine2%(AgenerUnião,Brazil,0,01ml/100gb.w.)andthen anesthetized with isoflurane inhalation (Isoflorine®
) (Cristália, Brazil).Asmalllaminectomyatthetenth/elevenththoracic lev-elswasperformed.Completetransectionscreateda4 mm-long gapattheeleventh/twelfthspinalcordlevels.Agelfoamsoaked in10lof0,9%salinewasleftatthebottomofthegapcloseto
thevertebralbody in10rats(SalineorControlgroup).Another groupof 10ratsreceivedgelfoamfilledwithsciaticnerve frag-ment(Nervegroup).Additionally,afurther10ratsreceivedsciatic nervefragmentand10lofaFGF-2(SantaCruzBiotechnology,
Germany)dissolvedin0.1Mphosphatebufferedsaline(PBS),pH 7.4,for48h.
Agroupof6uninjuredanimalswerealsoperfusedandtheir DRGs fromthe same location in the thoraxwere taken out. A microtome(Leica,SM2000R,Germany)wasusedtoprepare20m
thickentransversalfrozensectionsfromtheDRG.
2.4. ImmunohistochemicalproceduresforChAT
Immunoreactivitywasdetectedusingtheavidin–biotin peroxi-dasetechnique[31–33].Seriesofsectionswerewashed2×10min in0.1MPBS,pH7.4andincubatedwith5%normalgoatserum (NGS,Sigma)for 30minatroomtemperature.Serieswerethen incubatedfor24hat4◦Cwithgoatmonoclonalantiseraagainst ChAT(Millipore, diluted1:100). Theantibodieswere dilutedin PBScontaining0.3%TritonX-100(Sigma)and1%bovineserum albumin (Sigma). The series of sectionswere washed again in PBS (2×10min) and incubated with biotinylated donkey anti-goat(ChAT)immunoglobulinsdiluted1:1000(Jackson,USA)for1 hour.ThesectionswerewashedagaininPBSandincubatedwith anavidin–biotinperoxidasecomplex(bothdiluted1:100, Vectas-tain,Vector) for90min.Immunoreactivitywasvisualizedusing 3-3′-diaminobenzidinetetrahydrochloride(DAB,Sigma)asa chro-mogenandH2O2asasubstrate.
2.5. Morphometric/microdensitometricimageanalysis
TheChATimmunoreactivitywasmeasuredinonesectionper ratintheganglioncentralregion.Anopticalmicroscope (Olym-pusBX41)witha40×objectiveinbrightfieldwasusedtoobtain thedigitalimage.Thecountsandmeasurementsweremadeusing ImageJsoftware.Digitalimagesofrepresentativesectionswere obtainedusingadigitalvideocamera(NikonDXM1200).
2.6. Statisticalanalysis
Statisticalanalyseswereperformedusingtheanalysisof vari-ance(ANOVA)andsignificantinteractionswerefollowed-upwitha TukeyandBonferronipost-testcomparison.Allstatisticalanalyses wereperformedusingSPSS22,andsignificancewassetatp<0.05.
3. Results
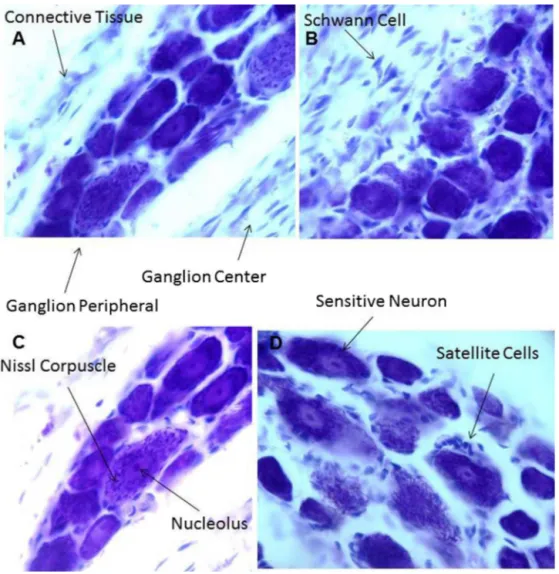
PhotomicrographsinFig.1illustratestheNisslstainedDRG neu-ronsofanimalswithoutinjury(Fig.1).
Uninjured animals used as parameters had a mean area of neuronal cell body of 9.1m, compared to the saline group
(6.86m2), the nerve group (7.85m2) and the nerve+FGF-2
group(8.54m2).Consideringtheinjuredgroups,thegroupthat
Fig.1. PhotomicrographsillustrateNissl-stainedDRGneuronsofanimalswithoutinjury.
Fig.2. AreaofChAT-immunoreactivecellbodiesintheDRGofuninjuredratstreatedwithsaline,graftsoffragmentsofthesciaticnerve(nerve)orfragmentsofthesciatic nervewithaddedFGF-2(nerve+FGF-2).Means±S.E.M.⋆p<0.05,⋆⋆p<0.01and⋆⋆⋆p<0.001accordingtoANOVA–TukeyandBonferroni.
ofcellbodyareacomparedtothesalinegroup(⋆⋆⋆p=0.0001)and
comparedtothenervegroup(⋆⋆p=0.006)(Fig.2).
TheprofilesofChATimmunoreactiveneuronalcellbody diam-eterintheDRG,intheuninjuredcontrolanimals,hadanaverage diameterof2.96mneuronal cellbody.Intheinjuredanimals,
thesalinegrouphad1.75m, inthenervegroup2.55m, and
in the nerve+FGF-2 group had 2.81m. Thus, the group that
Fig.3.DiameterofChAT-immunoreactivecellbodiesintheDRGofanimalstreatedwithsaline,graftsoffragmentsofthesciaticnerveorfragmentsofthesciaticnervewith addedFGF-2.Means±S.E.M.⋆p<0.05,⋆⋆p<0.01and⋆⋆⋆p<0.001accordingtoANOVA–TukeyandBonferroni.
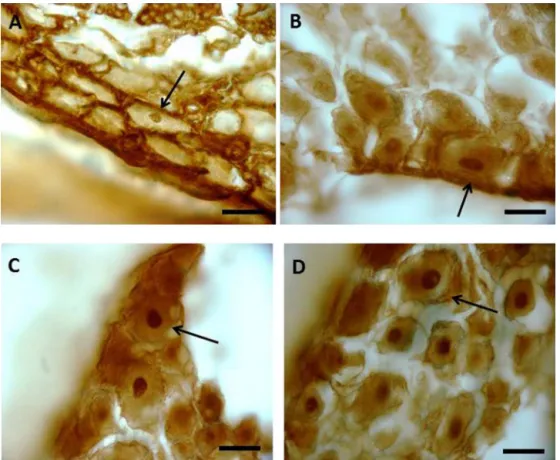
Fig.4.ChAT-immunoreactiveneurons.(A),salinegroup;(B),nervegroup;(CandD),nerve+FGF-2group.ArrowspointrelevantChAT-immunoreactivecellbody.Bars: 20mm2(A–D).
Fig.4A–C shows profiles of ChAT immunoreactive neuronal cell bodies of DRG frominjured groups. Fig. 4A (saline group) showsthecellbodyflatteningattheperipheryofganglion(arrow). Greaterpreservationwasobservedinthegroupsnerve(Fig.4B) andnerve+FGF-2(Fig.4CandD).
Morphologicalanalysisshowsthattheplasticbehaviorsinthe treatedgroupswerehigherthanthecontrolgroup.Thenodesof treatedanimalsshowedamorepreservedarchitecture(Fig.4B–D) thanthecontrol.
4. Discussion
theeffectsofexogenousFGF-2aremediatedmainlybythetyrosine kinaseFGFR1,whichislocalizedintheplasmamembraneaswell asinthenucleiofneuronsintheperipheralnerve[7,38,39].
Somereports have shown that peripheral nerve lesionscan accelerateregenerationofthecentralprimarysensoryneuronsin thedorsalspinalroots[40–42].However,mostoftheaxonsarestill unabletoreenterthespinalcord,andthosethatsucceedpenetrate nofurtherthanthesuperficialdorsalhornofthespinalcord.These reportsalsosuggestthataddingsome“intrinsicorextrinsic fac-tors”[42–44]canleadtotheregenerationofthecentralprocesses oftheDRG,towardgainingentryintothespinalcord.
Genetherapywithadenoviralvectorencodinganeurotrophinis capableofenhancinganddirectingtheregenerationofa subpopu-lationofdorsalrootaxonsthroughtotheCNSenvironmentinrats. However,theregeneratingaxons,inthestudycited,didnotspread widelythroughoutthegraymatterorawayfromtheinjectionsite, andnosensoryrecoverywasmentioned[45].
AnalysisofnervoustissueinratsshowedFGF-2 immunoreactiv-ityinmanydiversesizeneurons.Manyresearchershaveproposed bothneurotrophicandregenerativefunctionsforFGF-2.This sug-geststhatFGF-2mayperformsimilarfunctionsinDRGneurons [46,47].
OurfindingsshowChATimmunoreactivityintheDRGadjacent tothespinalcordinjury.Theanimalsthatreceivedtreatmentwith sciaticnervehadneuroprotectionwhentheareaanddiameterof cellbodieswereanalyzed,aneffectpotentiatedbyFGF-2.
Spinalcordtreatmentwithsciaticnerveandsciaticnerveplus FGF-2 allowedrecovery of hind limb movements compared to control,manifestedbysignificantlyhigherbehavioralscoresand higheramountsofMAP-2andGAP-43immunoreactive.Thus FGF-2addedtothenervegraftfavoredthemotorrecoveryandfiber regrowth[30].
FGF-1enhancesneuriteoutgrowthandstimulatesexpressionof GAP-43andT␣-tubulininculturedneuronsfromadultratdorsal
rootganglia.Aftertransectionandrepair,theanimalsthatreceived treatmentwithNGFshowedrecoveryinbothmotorandsensory nervefunctions[48,20].Duringperipheralnerveregeneration, FGF-2is up-regulatedinboth thecrushednerve andtherespective spinalganglia,suggestingapossiblephysiologicalfunctionofFGF-2 duringtheregenerationprocess[11].Treatmentwithacupuncture hasbeenseentopromoteeffectsontheexpressionofGDNFand FGF-2intheleftsixthlumbarDRGfollowingremovalofadjacent dorsalrootganglia[49].
Thesefindingsindicatethatourtreatmentstrategyusingnerve graftsalongwithFGF-2canenhanceaxonoutgrowthfromtheDRG, protectingthespinothalamicandspinocerebellartractsresulting inimprovementsinsensorystimulation,tone,postureand move-ment.
5. Conclusion
TheadditionofexogenousFGF-2toasciaticnerve fragment graftedintoagapontheratspinalcordwhichhadbeensubmitted tocompletetransectionisabletoimproveneuroprotectioninthe DRG.Theresultsemphasizedthatthemanipulationofthe microen-vironmentatthewoundisabletoamplifytheregenerativecapacity ofperipheralneurons.
Acknowledgments
ThisstudywassupportedbyfundingfromtheNationalCounsel ofTechnologicalandScientificDevelopment(CNPq)andthe Coor-dinationforImprovementofHighLevelStaff(CAPES).TheEnglish versionofthistextwasrevisedbySidneyPratt,Canadian,MAT(The JohnsHopkinsUniversity),RSAdip(TEFL,UniversityofCambridge).
References
[1]R.R.Ji,T.A.Samad,S.X.Jin,R.Schmoll,C.J.Woolf,P38MapkactivationbyNgf inprimarysensoryneuronsafterinflammationincreasesTrpv1levelsand maintainsheathyperalgesia,Neuron36(2002)57–68.
[2]F.Amaya,G.Shimosato,M.Nagano,M.Ueda,S.Hashimoto,Y.Tanaka,H. Suzuki,M.Tanaka,NgfandGdnfdifferentiallyregulatetrpv1expressionthat contributestodevelopmentofinflammatorythermalhyperalgesia,Eur.J. Neurosci.20(2004)2303–2310.
[3]S.Pezet,S.B.McMahon,Neurotrophins:mediatorsandmodulatorsofpain, Annu.Rev.Neurosci.29(2006)507–538.
[4]F.P.Guzen,R.J.deAlmeidaLeme,M.S.deAndrade,B.A.deLuca,G.Chadi,Glial cellline-derivedneurotrophicfactoraddedtoasciaticnervefragmentgrafted inaspinalcordgapamelioratesmotorimpairmentsinratsandincreaseslocal axonalgrowth’,Restor.Neurol.Neurosci.27(2009)1–16.
[5]P.Bohlen.Fibroblasticgrowthfactor.InMacrophage-derivedcellregulatory factors,Ed.Sorg.C,1(1989),204–228.
[6]B.Weise,K.Unsicker,C.Grothe,Localizationofbasicfibroblastgrowthfactor inasubpopulationofratsensoryneurons,CellTissueRes.267(1992) 125–130.
[7]C.Oellig,U.Pirvola,L.Taylor,R.Elde,T.Hokfelt,R.F.Pettersson,AcidicFgfand Fgfreceptorsarespecificallyexpressedinneuronsofdevelopingandadultrat dorsalrootganglia,Eur.J.Neurosci.7(1995)863–874.
[8]C.Grothe,C.Meisinger,P.Claus,Invivoexpressionandlocalizationofthe fibroblastgrowthfactorsystemintheintactandlesionedratperipheralnerve andspinalganglia,J.Comp.Neurol.434(2001)342–357.
[9]H.Liu,Q.F.Wu,J.Y.Li,X.J.Liu,K.C.Li,Y.Q.Zhong,D.Wu,Q.Wang,Y.J.Lu,L. Bao,X.Zhang,Fibroblastgrowthfactor7isanociceptivemodulatorsecreted vialargedense-corevesicles,J.Mol.Biol.Cell.(2015).
[10]R.R.Ji,Q.Zhang,X.Zhang,F.Piehl,T.Reilly,R.F.Pettersson,T.Hokfelt, ProminentexpressionofBfgfindorsalrootgangliaafteraxotomy,Eur.J. Neurosci.7(1995)2458–2468.
[11]C.Grothe,C.Meisinger,A.Hertenstein,H.Kurz,K.Wewetzer,Expressionof fibroblastgrowthfactor-2andfibroblastgrowthfactorreceptor1messenger Rnasinspinalgangliaandsciaticnerve:regulationafterperipheralnerve lesion,Neuroscience76(1997)123–135.
[12]D.Otto,K.Unsicker,C.Grothe,Pharmacologicaleffectsofnervegrowthfactor andfibroblastgrowthfactorappliedtothetransectionedsciaticnerveon neurondeathinadultratdorsalrootganglia,Neurosci.Lett.83(1987) 156–160.
[13]R.Elde,Y.H.Cao,A.Cintra,T.C.Brelje,M.Pelto-Huikko,T.Junttila,K.Fuxe,R.F. Pettersson,T.Hokfelt,Prominentexpressionofacidicfibroblastgrowthfactor inmotorandsensoryneurons,Neuron7(1991)349–364.
[14]B.Weise,T.Janet,C.Grothe,LocalizationofBfgfandFgf-receptorinthe developingnervoussystemoftheembryonicandnewbornrat,J.Neurosci. Res.34(1993)442–453.
[15]C.Unsicker,G.L ¨udecke,D.Otto,R.Westermann,Fibroblastgrowthfactors: theirrolesinthecentralandperipheralnervoussystem,in:S.E.Loughlin,J.H. Fallon(Eds.),NeurotrophicFactors,AcademicPress,NewYork,London,1993, pp.313–338.
[16]C.Grothe,K.Wewetzer,Fibroblastgrowthfactoranditsimplicationsfor developingandregeneratingneurons,Int.J.Dev.Biol.40(1996)403–410.
[17]T.P.Carlstedt,H.Linda,S.Culheim,Reinnervationofhindlimbmusclesafter ventralrootavulsionandimplantationinthelumbarspinalcordoftheadult rat,ActaPhysiol.Scand.128(1989)645–646.
[18]S.Cullheim,T.Carlstedt,H.Linda,M.Risling,B.Ulfhake,Motoneurons reinnervateskeletalmuscleafterventralrootimplantationintothespinal cordofthecat,Neuroscience29(1989)725–733.
[19]T.P.Carlstedt,R.G.Hallin,K.G.Hedstrom,I.A.Nilsson-Remahl,Functional recoveryinprimateswithbrachialplexusinjuryafterspinalcord
implantationofavulsedventralroots,J.Neurol.Neurosurg.Psychiatry(1993) 649–654.
[20]F.Huang,Z.Liu,H.Liu,L.Wang,H.Wang,Z.Li,Gm1andNgfModulateCa2+ homeostasisandGap43Mrnaexpressionincultureddorsalrootganglion neuronswithexcitotoxicityinducedbyglutamate,Nutr.Neurosci.10(2007) 105–111.
[21]T.Carlstedt,P.Grane,R.G.Hallin,G.Noren,Returnoffunctionafterspinalcord implantationofavulsedspinalnerveroots,Lancet346(1995)1323–1325.
[22]T.Y.Chuang,M.C.Huang,K.C.Chen,Y.C.Chang,Y.S.Yen,L.S.Lee,H.Cheng, Forelimbmuscleactivityfollowingnervegraftrepairofventralrootsinthe ratcervicalspinalcord,LifeSci.71(2002)487–496.
[23]C.F.Hoffmann,E.Marani,J.G.vanDijk,W.vdKamp,R.T.Thomeer, Reinnervationofavulsedandreimplantedventralrootletsinthecervical spinalcordofthecat,J.Neurosurg.84(1996)234–243.
[24]J.J.Katims,Electrodiagnosticfunctionalsensoryevalutaionofthepatientwith pain:areviewoftheneuroselectivecurrentperceptionthersholdandpain tolerancethershold,PainDig.8(1998)219–230.
[25]Y.Iwasaki,T.Shiojima,K.Ikeda,N.Tagaya,T.Kobayashi,M.Kinoshita,Acidic andbasicfibroblastgrowthfactorsenhanceneuriteoutgrowthinculturedrat spinalcordneurons,Neurol.Res.17(1995)70–72.
Neurochem.Int.25(1994)39–45.
[33]K.Fuxe,B.Tinner,G.Chadi,A.Harfstrand,L.F.Agnati,Evidenceforaregional distributionofhyaluronicacidintheratbrainusingahighlyspecific hyaluronicacidrecognizingprotein,Neurosci.Lett.169(1994)25–30.
[34]C.Basilico,D.Moscatelli,TheFgffamilyofgrowthfactorsandoncogenes,Adv. CancerRes.59(1992)115–165.
[35]A.J.Furusho,R.J.Franklin,R.Bansal,Fibroblastgrowthfactorsignalingin oligodendrocyte-lineagecellsfacilitatesrecoveryofchronicallydemyelinated lesionsbutisredundantinacutelesions,Glia(2015).
[36]C.Meisinger,C.Grothe,Differentialregulationoffibroblastgrowthfactor (Fgf)-2andFgfreceptor1MrnasandFgf-2isoformsinspinalgangliaand sciaticnerveafterperipheralnervelesion,J.Neurochem.6(1997)1150–1158.
[37]D.E.Johnson,L.T.Williams,Structuralandfunctionaldiversityinthefgf receptormultigenefamily,Adv.CancerRes.60(1993)1–41.
[38]L.Klimaschewski,C.Meisinger,C.Grothe,Localizationandregulationofbasic fibroblastgrowthfactor(Fgf-2)andFgfreceptor-1inratsuperiorcervical ganglionafteraxotomy,J.Neurobiol.38(1999)499–506.
[39]W.Nindl,P.Kavakebi,P.Claus,C.Grothe,K.Pfaller,L.Klimaschewski, Expressionofbasicfibroblastgrowthfactorisoformsinpostmitotic sympatheticneurons:synthesis,intracellularlocalizationandinvolvementin karyokinesis,Neuroscience124(2004)561–572.
[46]B.Malgrange,P.Delree,J.M.Rigo,H.Baron,G.Moonen,Imageanalysisof neuriticregenerationbyadultratdorsalrootganglionneuronsinculture: quantificationoftheneurotoxicityofanticanceragentsandofitsprevention bynervegrowthfactororbasicfibroblastgrowthfactorbutnotbrain-derived neurotrophicfactororneurotrophin-3’,J.Neurosci.Methods53(1994) 111–122.
[47]C.G.Acosta,A.R.Fabrega,D.H.Masco,H.S.Lopez,Asensoryneuron subpopulationwithuniquesequentialsurvivaldependenceonnervegrowth factorandbasicfibroblastgrowthfactorduringdevelopment,J.Neurosci.21 (2001)8873–8885.
[48]L.Mohiuddin,P.Fernyhough,D.R.Tomlinson,Acidicfibroblastgrowthfactor enhancesneuriteoutgrowthandstimulatesexpressionofGAP-43andTalpha 1alpha-tubulininculturedneuronesfromadultratdorsalrootganglia, Neurosci.Lett.215(1996)111–114.