___________________________
Corresponding author: Renata Iličić, University of Novi Sad, Faculty of Agriculture, Trg Dositeja Obradovića 8, 21000 Novi Sad, Serbia
Original scientific paper
CHARACTERIZATION OFPseudomonas syringaePATHOVARS FROM DIFFERENT
SWEET CHERRY CULTIVARS BY RAPD ANALYSIS
Renata ILIČIĆ(1), Jelica BALAŽ(1), Vera STOJŠIN(1), Dragana JOŠIĆ(2) (1)University of Novi Sad, Faculty of Agriculture, , Novi Sad, Serbia.
(2)Institute of Soil Science, Genetic Lab, Belgrade, Serbia.
Iličić R., J. Balaž, V.Stojšin, D. Jošić (2016):Characterization of Pseudomonas syringae pathovars from different sweet cherry cultivars by RAPD analysis.- Genetika vol 48, no1, 285-295.
Pseudomonas syringaepvs., isolated from sweet cherry grown on different localities in Serbia, were genetically characterized using RAPD analysis. Four out of eleven tested primers (SPH1, DJP 17, DJ 15, and DJ 16) were selected on the basis of the differences between isolates within two pathovars -syringaeandmorsprunorumrace 1. Cumulative RAPD analysis indicated heterogeneity within the population of both groups of tested isolates, revealing four different patterns in each group.RAPD analysis showed up to 24% differences among pv. syringae isolates, as well as 41% in comparison with the reference strain KFB0103 (pv.syringae), while differences of 15% among isolates pv. morsprunorum 1 race and 36% compared to the reference strain CFBP2119 (pv. morsprunorum1) were observed. Isolates from locality Selenca exhibited three different genotypic patterns of pv.morsprunorumrace 1 and one pattern of pv.syringae. Isolates of pv.morsprunorum collected in the same year from two plant organs (branches and leaves) of the cv. Vanda yielded two different patterns. The pv.morsprunorum on cv. Kordia and pv. syringaeon cv. Regina were detected at Mikicevo locality. The same patterns were observed for isolates of pv.syringaefrom Kanjiza and Selenca, as well as from Gornji Tavankut in two years of isolation. Differences were noted between isolates from the same pathovar originating from Ljutovo and Mikicevo, as well as with respect to all other isolates of same pathovar.
INTRODUCTION
Pseudomonas syringae van Hall (1902) is polyphagous and widely spread species of phytopathogenic bacteria and is recognized as an economically important causal agent of bacterial diseases in numerous plants. According toGARDANet al. (1999), on the basis of DNA homology, bacteria P. syringae is divided into more than 60 pathovars, as well as nine genomospecies. Various molecular techniques have been used to characterize P. s. pv. syringae strains. Polymerase chain reaction (PCR) method for identification of phytopathogenic bacteria at the molecular level has been used in many previous studies (SCORTICHINIet al., 2003;NATALINI et al., 2006; GILBERTet al., 2009; KALUZNA et al., 2010; GAŠIĆ et al., 2012; GAVRILOVIĆ et al., 2012, 2013; IVANOVIĆet al.,2009, 2012). Compared to most classical bacteriological tests, this method allows for better characterization of bacteria. As molecular methods are much more exact, they facilitate reliable differentiation of the bacterial population, especially in the determination ofP. syringae. Nowadays, the most widely utilized approaches in this category include Pulsed-Field Gel Electrophoresis (PFGE), Restriction Fragment Length Polymorphism (RFLP), Random Amplified Polymorphic DNA (RAPD), repetitive – sequence PCR (rep – PCR), Amplified 16S Ribosomal DNA Restriction Analysis (ARDRA), Amplified Fragment Length Polymorphism (AFLP) and MLST (SCORTICHINIet al.,2003;VICENTEet al., 2004;VICENTEandROBERTS, 2007; GILBERTet al., 2009;KALUZNAet al., 2010). Identification of different pathovars from stone fruits was achieved by detection of gene for syringomycin synthesis (syrB) and syringomycin secretion (syrD) specific toP. s.pv.syringaeand production of coronatine (cfl) forP. s.pv.morsprunorum race 1 and P. s. pv. morsprunorum race 2 gene to produce siderophores yersiniabaktin (irp1) (SCORTICHINI et al., 2003; GAŠIĆ et al., 2012). New classification of bacteria based on DNA hybridization revealed thatP. s.pv.morsprunorumrace 1 andP. s.pv.morsprunorumrace 2 are two distinct organisms, because they belong to a genomospecies 2 and 3, whileP. s.pv.syringae belongs genomospecies 1 (GARDAN et al., 1999). Thus, given the significant diversity of P. syringae isolates and the evidence indicating their spreading to new hosts, there is a need to implement methods that allow comparison of isolates of the same or different species of bacteria. RAPD is a valuable method that allows for successful comparison and estimation of differences in the population ofP. syringaepvs. from stone fruit. In addition, application of RAPD analysis can reveal specific genotypic changes inP. syringae(WILLIAMSet al., 1990).
The aim of this study was to characterize isolates of P. syringae pvs. originating from sweet cherry using RAPD method and to estimate genodiversity in two pathovars (syringaeand morsprunorumrace 1).
MATERIALS AND METHODS
For this study, 51 isolates of P. syringaepvs.syringaeand 53 ofmorsprunorumrace 1 were obtained from diseased sweet cherries from several localities in Vojvodina Province. Strains CFBP2119 (pv. morsprunorumrace 1), 765 (pv.lachrymans), Tk21 (pv.syringae) and KFB0103 (pv.syringae) were used as reference. List of representative isolates and reference strains is given in Table 1.
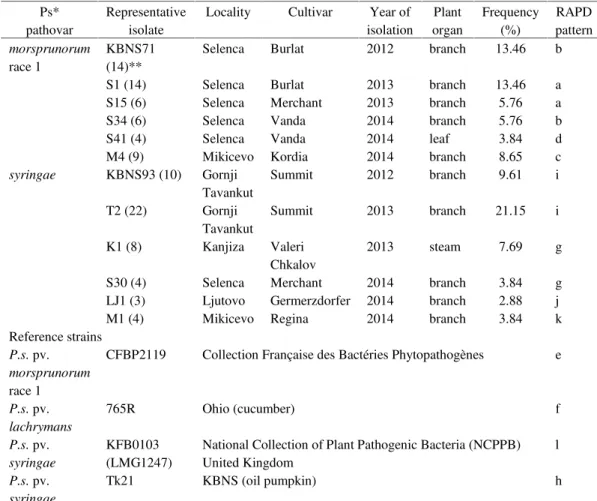
Table 1. Origin and characterization of Pseudomonas syringae isolates and a list of reference strains Ps* pathovar Representative isolate
Locality Cultivar Year of isolation Plant organ Frequency (%) RAPD pattern morsprunorum race 1 KBNS71 (14)**
Selenca Burlat 2012 branch 13.46 b
S1 (14) Selenca Burlat 2013 branch 13.46 a S15 (6) Selenca Merchant 2013 branch 5.76 a S34 (6) Selenca Vanda 2014 branch 5.76 b S41 (4) Selenca Vanda 2014 leaf 3.84 d M4 (9) Mikicevo Kordia 2014 branch 8.65 c syringae KBNS93 (10) Gornji
Tavankut
Summit 2012 branch 9.61 i
T2 (22) Gornji Tavankut
Summit 2013 branch 21.15 i
K1 (8) Kanjiza Valeri Chkalov
2013 steam 7.69 g
S30 (4) Selenca Merchant 2014 branch 3.84 g LJ1 (3) Ljutovo Germerzdorfer 2014 branch 2.88 j M1 (4) Mikicevo Regina 2014 branch 3.84 k Reference strains
P.s.pv. morsprunorum race 1
CFBP2119 Collection Française des Bactéries Phytopathogènes e
P.s.pv. lachrymans
765R Ohio (cucumber) f
P.s.pv. syringae
KFB0103 (LMG1247)
National Collection of Plant Pathogenic Bacteria (NCPPB) United Kingdom
l
P.s.pv. syringae
Tk21 KBNS (oil pumpkin) h
* pathovar identification on the basis of LOPAT and GATTa tests, as well as presence of syrB/syrD or cflgenes (unpublished data); ** number of isolates is given in parenthesis
(unpublished data). The size of amplicon was determined by comparison with the DNA molecular weight markers GeneRuler DNA Ladder mix SM0331 and SM0243 (Thermo Scientific, Lithuania). Four primers (SPH1, DJP17, DJ15 and DJ16) were chosen to estimate a diversity of P. syringaeisolates into each pathovar. All PCR products were visualized by electrophoresis on 1.5% agarose gel in 0.5 Tris – Borate – EDTA buffer at 5Vcm-1 for 3h, stained with ethidium bromide and photographed under UV Transilluminator. The assessment of similarity among the isolates of P. syringae pvs. was based upon the simple matching coefficient (SSM). The formation of clusters was carried out based on the UPGMA (Unweighted Pair Group Method with Arithmetic Mean) analysis using STATISTICA 12 software.
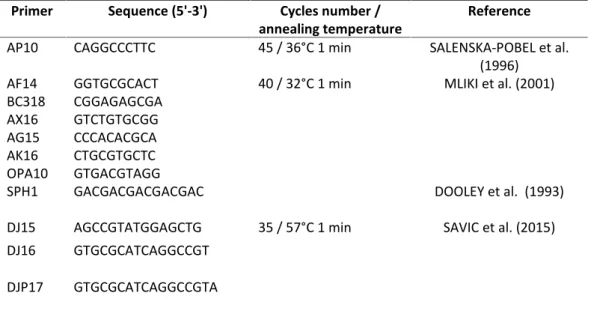
Table 2. Primers and amplification conditions
Primer Sequence (5'-3') Cycles number /
annealing temperature
Reference
AP10 CAGGCCCTTC 45 / 36°C 1 min SALENSKA-POBEL et al.
(1996)
AF14 GGTGCGCACT 40 / 32°C 1 min MLIKI et al. (2001)
BC318 CGGAGAGCGA AX16 GTCTGTGCGG AG15 CCCACACGCA AK16 CTGCGTGCTC OPA10 GTGACGTAGG
SPH1 GACGACGACGACGAC DOOLEY et al. (1993)
DJ15 AGCCGTATGGAGCTG 35 / 57°C 1 min SAVIC et al. (2015) DJ16 GTGCGCATCAGGCCGT
DJP17 GTGCGCATCAGGCCGTA
RESULTS AND DISCUSSION
(8.65%) and d (3.84%). The remaining 49.03% of isolates belong to P. s. pv. syringae, with 30.76% showing genotypic patterni, followed byg(11.53%),k(3.84%) andj(2.88%).
The cumulative analysis findings revealed two clusters, segregating isolates according to pathovars, with 69% differences between them. The first cluster included members of pv. syringae grouped in two distinct subclusters, one of which contained the strain KFB0103 only (with 41% difference). The second subcluster consisted two branches (differing by 24%), with isolates M1, LJ1, T2 and KBNS93 belonging to the first, and K1 and S30, as well as the strain Tk21, to the second. The strain Tk21 exhibited 12% differences relative to the K1 and S30 isolates.
The second cluster consisted two subclusters, differing by 42.5%, dividing the strain 765R (pv.lachrymans) and members of pv.morsprunorumrace 1. The subcluster members of pv. morsprunorum race 1 contained two branches with 36% mutual differences. While only strain CFBP2119 was assigned to the first branch, isolates S41, M4, KBNS71, S34, S15, and S1 were in the second. The difference of 15% was observed among these isolates.
Figure 2. RAPD – Primer DJP17 ofP. syringaeisolates originating from sweet cherry.P. s.pv. syringae: KBNS93, S30, T2, K1, Lj1, M1;P. s.pv.morsprunorumrace 1: S1, S15, S34, S41, M4, KBNS71; Reference strains: CFBP2119 –P. s. pv.morsprunorumrace 1; KFB0103 – P. s.pv.syringae; 765R –P. s.pv.lachrymans; Tk21 –P. s.pv.syringae;Marker SM0331– GeneRuler DNA Ladder Mix Thermo Scientific, Lithuania.
Figure 4. RAPD – primer DJ16 ofP. syringaeisolates originating from sweet cherryP. s. pv. syringae: KBNS93, S30, T2, K1, Lj1, M1;P. s.pv.morsprunorumrace 1: S1, S15, S34, S41, M4, KBNS71; Reference strains: CFBP 2119 –P. s. pv.morsprunorumrace 1; KFB0103 –P. s.pv.syringae; 765R –P. s.pv.lachrymans; Tk21 –P. s.pv.syringae; Marker SM0331– GeneRuler DNA Ladder Mix Thermo Scientific, Lithuania; Marker SM0243 –GeneRuler DNA Ladder Mix Thermo Scientific, Lithuania.
RAPD ZBIRNI Complete Linkage Percent disagreement
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 Linkage Distance
KFB0103 M1 LJ1 T 2 KBNS93 T k21 K1 S30 765R CFBP2119 S41 M4 KBNS71 S34 S15 S1
It is widely accepted that the isolates ofP. s.pv.syringaeare characterized by greater genetic variability compared to those of the pv. morsprunorum(different geographic origin and hosts) (KHAYAMIEet al.,2009;AFROSEet al.,2014). In this investigation, 104 isolates from five localities and eight cultivars were tested. Among 11 primers used in RAPD analysis, AF14, AX16, AG15, AK16 and OPA10 yielded patterns of very poor resolution. On the other hand, AP10 and BC318 amplified different patterns forP. s. pv.syringaeandP. s.pv.morsprunorum race 1; however, they failed to reveal differences among isolates within the same pathovar. Primers SPH1, DJP17, DJ15 and DJ16 amplified high-resolution patterns and thus emphasized differences between the isolates. Based on cumulative analysis using four primers (SPH1, DJP17, DJ15, and DJ16), the differences among tested pv. syringae isolates ranged from 0 to 24%, reaching 41% comparing to the strain KFB0103 (pv.syringae). Differences of up to 15% were observed for pv. morsprunorum race 1 isolates, which increased to 36% when these were compared to the strain CFBP2119. While RAPD analysis suggested certain heterogeneity in both pathovars, genetic diversity was more pronounced among pv.syringaeisolates. Strain 765RP. s. pv. lachrymans showed greater similarity to pv. morsprunorum race 1 isolates and strain CFBP2119, forming a common cluster, but separate subclusters, with differences of about 42.5%. This method has been used in previous investigations ofP. syringaewith different hosts in Serbia (ILIČIĆet al., 2014). Successful identification of the isolates and the presence of greater genetic diversity within the group pv.syringaeoriginating from stone fruit, hazelnut and olive trees using RAPD analysis was reported by KHAYAMIE et al. (2009). According to the same authors, the ability to differentiate isolates is useful in the epidemiology of bacteria canker of stone fruit and host-pathogen interactions relating to stone fruit cultivar infection. It can also find applications in the development of improved strategies for breeding and resistance to P. s.pv.syringae.RAPD can have application in the evaluation of genetic diversity among isolates ofP. s.pv.syringaeand has a particularly significant importance in studying diversity among isolates isolated from the same geographical location, region and host (AFROSEet al., 2014). RAPD analysis is useful for characterizing phytopathogenic bacteria and identifying specific genotypic changes inP. syringae (WILLIAMSet al., 1990).
According to the results pertaining to isolates belonging to pv.morsprunorumrace 1, the same pattern (a) was observed for isolates from the locality Selenca, isolated from diseased branches of sweet cherry cultivars Burlat and Merchant in the same year of isolation (2013), with frequency of 19.22%. In different years of isolation from different cultivars (Burlat in 2012 and Vanda in 2014), isolates KBNS71 and S34 also revealed the same pattern (b, with the frequency of 19.22%). Isolates obtained from diseased branches and leaves of cv. Vanda, grown on Selenca plantation, formed different patterns (isolates S34 and S41, respectively). Isolates of pv. morsprunorum race 1 from locality Mikicevo showed different pattern from those originating from locality Selenca (c, frequency 8.65%).
CONCLUSIONS
Four primers (SPH1, DJP17, DJ15 and DJ16) used for RAPD yielded profiles with high resolution. Differences among isolates of pv.syringaeand pv.morsprunorumrace 1 (up to 24% and 15%, respectively) represented by four genotypic patterns for each pathovar were observed, suggesting a certain heterogeneity; however, the differences were more pronounced in pv. syringae. Both pathovars were detected on the sweet cherry cv. Merchant grown in Selenca collected in two different years, as well as in Mikicevo locality collected in the same year, but on different cvs (Kordia and Regina).
ACKNOWLEDGMENTS
This work was supported by Serbian Ministry of Education, Science and Technological Development, Project No. III46007.
Received November 13st, 2015
Accepted February 16th, 2016
REFERENCES
AFROSE,S.,I.HOSSAIN,M.R.ISLAM,M.D,HOSSAINandM.A.H.KHAN(2014): RAPD analyses reveals the genetic diversity of the litchi leaf blight pathogen,Pseudomonas syringae pv. syringae in Bangladesh. Curr Res Microbiol Biotechnol., Vol 2, No.1, 301-309.
DOOLEY,J.J.,S.P.HARRISON,L.R.MYTTON,M.DYE,A.CRESSWELL,L.SKOT andJ.R.BEECHING(1993): Phylogenetic grouping and identification ofRhizobiumisolates on the basis of random amplified polymorphic DNA profiles. Can J Microbiol.,39: 665-673.
GARDAN,L.,H.SHAFIK,S.BELOUIN,R.BROCH,F.GRIMONTandP.A.D.GRIMONT(1999): DNA relatedness among the pathovars ofPseudomonas syringae and description of Pseudomonas tremaesp. nov. and Pseudomonas cannabinasp. nov. (ex Sutic and Dowson 1959). Int J Syst Bacteriol.,49: 469-478.
GAŠIĆ,K.,A.PROKIĆ,M.IVANOVIĆ,N.KUZMANOVIĆandA.OBRADOVIĆ(2012): Differentiation ofPseudomonas syringae pathovars originating from stone fruits. Pestic Phytomed., 27(3): 219-229.
GAVRILOVIĆ,V.,S.ŽIVKOVIĆ,N.DOLOVAC,N.TRKULJA, E. DOLOVAC– PFAF,T.POPOVIĆandŽ.IVANOVIĆ (2012): Pseudomonas syringae- pathogen of sweet cherry in Serbia. Pestic Phytomed.,27(2): 141-149.
GAVRILOVIĆ, V., Ž. IVANOVIĆ, T. POPOVIĆ, S. ŽIVKOVIĆ, S. STANKOVIĆ, T. BERIĆ and Đ. FIRA (2013): Genetic characterization of pathogenic fluorescent Pseudomonads isolated from necrotic cherry and plum buds in Serbia. Genetika, Vol45, No.3: 953-961.
GILBERT,V.,F.LEGROS,H.MARAITEandA.BULTREYS(2009): Genetic analyses ofPseudomonas syringaeisolates from Belgian fruit orchards reveal genetic variability and isolate-host relationships within the pathovarsyringae, and help identify both races of the pathovarmorsprunorum. Eur J Plant Pathol.,124:199-218.
ILIČIĆ, R., J. BALAŽ and D.JOŠIĆ (2014): Characterization of two Pseudomonas syringae pathovars (syringaeand morsprunorum) from sweet cherry by RAPD. Book of abstact, V Congress of the genetic society, Kladovo, Serbia, September 28th – October 2nd, Ed. Vasiljevic B., S. Mladenovic Drinic. Publisher: Serbian Genetic Society, Belgrade. p. 218.
IVANOVIĆ, Ž., S. ŽIVKOVIĆ, M. STAROVIĆ, D. JOŠIĆ, S. STANKOVIĆ and V. GAVRILOVIĆ (2009): Diversity among Pseudomonas syringaestrains originating from fruit trees in Serbia. Arch Biol Sci.,61 (4): 863-870.
KALUZNA M.,P.FERRANTE,P. SOBICZEWSKIand M.SCORTICHINI (2010): Characterization and genetic diversity of Pseudomonas syringaeisolates from stone fruits and hazelnut using repetitive-PCR and MLST. J Plant Pathol, 92 (3): 781-787.
KHAYAMIE,S.,K.N.NIKNEJAD,B.RABIE,S.SASSANIEandA.A.EBADIE(2009): Genetic characterization ofP. syringaepv. syringaestrains from stone fruits based on RAPD analysis in Iran. Agriculturatropicaetsubtropica,42(4) :162-166.
MLIKI,A.,E.J.STAUB,S.ZHANG YONGandG.ABDELWAHED(2001): Genetic diversity in melon (Cucumis meloL.): An evaluation of African germplasm. Genet Resour Crop Ev.,48: 587-597.
NATALINI,E.,M.P.ROSSI,D.BARIONOVIandM.SCORTICHINI(2006): Genetic and pathogenic diversity ofPseudomonas syringaepv.syringaeisolates associated with bud necrosis and leaf spot of pear in a single orchard. J Plant Pathol.,88(2): 219-223.
SAVIĆ D.,D.JOŠIC,E.RISTANOVIĆ,R.PIVIĆ,A.STANOJKOVIĆ-SEBIĆandZ.LEPSANOVIĆ(2015): Detection of toxin genes and RAPD analysis ofBacillus cereusisolates from different soil types. Genetika,47 (2): 627-638.
SCORTICHINI,M.,U.MARCHESI,M.T.DETTORIandM.P.ROSSI(2003): Genetic diversity, presence of thesyrB gene, host preference and virulence ofPseudomonas syringaepv. syringaestrains from woody and herbaceous host plants. Plant Pathol.,52: 277-286.
SELENSKA– POBELL, S.,E. EVGUENIEVA–HACKERBERG,G. RADEVAandA.SQUARTINI(1996): Characterization of Rhizobium hedusariby RFLP analysis of PCR amplified rDNA and by genomic fingerprinting. J Appl Bacteriol.,50: 517-28.
VICENTE,J. andS.ROBERTS(2007): Discrimination of Pseudomonas syringae isolates from sweet and wild cherry using rep – PCR. Eur J Plant Pathol.,117: 383-392.
VICENTE,J.G.,S.J.ROBERTS,K.RUSSELLandJ.P.ALVES(2004): Identification and discrimination ofPseudomonas syringae isolates from wild cherry in England. Eur J Plant Pathol.,110: 337-351.
KARAKTERIZACIJAPSEUDOMONAS SYRINGAEPATOVARA SA RAZLIČITIH
SORTI TREŠNJE KORIŠĆENJEM RAPD ANALIZE
Renata ILIČIĆ(1), Jelica BALAŽ(1), Vera STOJŠIN(1), Dragana JOŠIĆ(2) (1)Univerzitet u Novom Sadu, Poljoprivredni fakultet, Novi Sad, Srbija
(2)Institut za zemljište, genetička laboratorija, Srbija Izvod
Pseudomonas syringae pvs., izolovani od trešnje na različitim lokalitetima u Srbiji, genetski su okarakterisani pomoću RAPD analize. Četiri od 11 testiranih prajmera (SPH1, DJP17, DJ15 DJ, 16) su odabrani na osnovu razlike između izolata unutar dva patovara -syringaeimorsprunorum rase 1. Zbirna RAPD analiza pokazala je heterogenost unutar populacije obe grupe testiranih izolata, pokazujući četiri različita profila u svakoj grupi. RAPD analiza je pokazala do 24% razlike među izolatima pv. syringae i 41% u poređenju sa referentnim sojem KFB0103 (pv. syringae), dok su razlike od 15% zabeležene među izolatima pv. morsprunorumrase 1 i 36% u odnosu na referentni soj CFBP2119 (pv.morsprunorum rasa 1.). Izolati poreklom iz Selenče su pokazali tri različita genotipska profila pv.morsprunorumrase 1 i jedan profil pv.syringae. Dva različita profila pv.morsprunorumrase 1 su utvrđena na sorti Vanda u istoj godini – poreklom sa obolelih grana i lišća. Patovarmorsprunorumrasa 1 na sorti Kordia i pv.syringaena sorti Regina su utvrđena na lokalitetu Mikicevo. Isti profili utvrđeni su za izolate pv. syringaeporeklom iz Kanjiže i Selenče, kao i za izolate iz Gornjeg Tavankuta u dve godine izolacije. Izolati istog pathovara poreklom iz Ljutova i Mikiceva su međusobno bili različiti kao i između ostalih izolata istog pathovara.