www.cbpv.org.br/rbpv ISSN 0103-846X (Print) / ISSN 1984-2961 (Electronic)
Doi: http://dx.doi.org/10.1590/S1984-29612017062
Seasonality of parasitic helminths of
Leporinus macrocephalus
and their parasitism rates in farming systems in the Amazon
Sazonalidade de helmintos parasitos de
Leporinus macrocephalus
e seus índices parasitários
em sistemas de cultivo na Amazônia
Williane Maria de Oliveira Martins1*; Márcia Cristina Nascimento Justo2; Melissa Querido Cárdenas2;
Simone Chinicz Cohen2
1 Instituto Federal do Acre – IFAC, Cruzeiro do Sul, AC, Brasil
2 Laboratório de Helmintos Parasitos de Peixes – LHPP, Instituto Oswaldo Cruz – IOC, Fundação Oswaldo Cruz – FIOCRUZ,
Rio de Janeiro, RJ, Brasil
Received June 9, 2017 Accepted September 26, 2017
Abstract
The objective of the present study was to analyze the seasonality of parasitic helminths of Leporinus macrocephalus
from fish farms in the municipality of Cruzeiro do Sul, Acre, Brazil, and their parasitism rates. Between June 2014 and March 2015, 200 specimens were sampled from two fish farms: one with a semi-intensive system and the other with an extensive system (100 fish from each farm: 50 during the dry season and 50 during the rainy season). Fifteen species of parasites were found, with seasonal variations of some according to the farming system. In the semi-intensive fish farm, there was greater prevalence of infection during the dry season. Also, Urocleidoides paradoxus, Procamallanus
(Spirocamallanus) inopinatus, Goezia leporini and Rhabdochona (Rhabdochona) acuminata presented differences in their parasitism rates between the seasons. In the extensive fish farm, no variation in the prevalence of infection was observed between the seasons and two species Tereancistrum parvus and G. leporini demonstrated differences only regarding the mean intensity of infection. The data presented here may help fish farmers to understand the parasite dynamics of
L. macrocephalus in farming systems during the dry and rainy seasons in the state of Acre. Keywords: Amazon, fish farm, parasites, seasonal variation.
Resumo
O presente estudo teve como objetivo analisar a sazonalidade dos helmintos parasitos de Leporinus macrocephalus
provenientes de pisciculturas no município de Cruzeiro do Sul, Acre, Brasil e seus índices parasitários. Durante o período de junho de 2014 a março de 2015 foram coletados 200 espécimes obtidos de duas pisciculturas, uma de sistema semi-intensivo e outra de sistema extensivo, sendo 100 peixes em cada, durante as estações seca e chuvosa, 50 espécimes por estação. Foram registradas quinze espécies de parasitos, com variação sazonal de ocorrência de alguns nos sistemas de cultivo. Na piscicultura semi-intensiva houve maior prevalência de infecção na seca, e Urocleidoides paradoxus,
Procamallanus (Spirocamallanus) inopinatus, Goezia leporini e Rhabdochona (Rhabdochona) acuminata apresentaram diferenças dos índices parasitários entre as estações. Na piscicultura extensiva não foi detectada variação da prevalência de infecção entre as estações e duas espécies, Tereancistrum parvus e G. leporini assinalaram diferença apenas na intensidade média de infecção. Os dados apresentados neste estudo podem auxiliar os piscicultores na compreensão da dinâmica parasitária de L. macrocephalus em sistemas de cultivos durante as estações seca e chuvosa no estado do Acre.
Palavras-chave: Amazônia, piscicultura, parasitos, variação sazonal.
*Corresponding author: Williane Maria de Oliveira Martins. Instituto Federal do Acre – IFAC, Estrada da Apadec, 1192, Nova Olinda, CEP 69980-000, Cruzeiro do Sul, AC, Brasil. e-mail: [email protected]
Introduction
Although fishery resources are abundant in the Amazon region, depletion of natural stocks of some species of fish, together with availability of water resources and increased demand for fish, has
2006; PAVANELLI et al., 2013; ZAGO et al., 2014), culminating in significant production losses.
The Amazon region presents clear seasonality: a rainy season that usually occurs between November and April, and a dry season from May to October (FIGUEROA & NOBRE, 1990). At this latitude, there is little variation in temperature and intensity of sunlight (SILVA et al., 2008). The mean temperature is above 30 °C, which is a factor that can cause stress among fish, thus favoring multiplication of parasites (MARINHO et al., 2013). Abiotic variations, such as high temperatures and low levels of oxygen can influence the abundance of some species of parasites or the abundance of intermediate hosts (TAVARES-DIAS et al., 2001; SCHALCH & MORAES, 2005; MARINHO et al., 2013; ZAGO et al., 2014; JERÔNIMO et al., 2016).
Investigations in the Amazon region have only addressed seasonal variations of parasite communities in rivers and lakes (VITAL et al., 2011; TAVARES-DIAS et al., 2014; VASCONCELOS & TAVARES-DIAS, 2014). Little is known about the influence of seasonality on occurrences of parasites and rates of parasitism in fish farms. This is particularly true regarding species that were introduced to farms in this region, such as Leporinus macrocephalus
Garavello and Britisky 1988 (Anostomidae), commonly known as piavuçu.
Among the species of fish that are farmed in this region,
L. macrocephalus has gained prominence through its high production
capacity, with harvesting in approximately six months. However, various diseases and parasite infestations can affect the production of this fish, including species of Monogenoidea and Nematoda (TAVARES-DIAS et al., 1999; MARTINS & YOSHITOSHI, 2003; SCHALCH & MORAES, 2005).
Parasites are abundant in tropical regions and their presence is one of the main causes of death among fish (AGUINAGA et al., 2015). Thus, understanding the agents that cause parasitic diseases and the complex relationship between environmental factors and hosts and their parasites is important. With such knowledge, interventions in the system using adequate prevention techniques to create control programs for these diseases become possible. Within this context, the objective of the present study was to analyze the seasonality of helminths that parasitize L. macrocephalus in fish
farms in the municipality of Cruzeiro do Sul, Acre, Brazil, and their parasitism rates.
Materials and Methods
Sampling and fish farms
Between June 2014 and March 2015, 200 specimens of
L. macrocephalus were collected from two fish farms. One of these fish farms used a semi-intensive system, while the other used an extensive farming system. One hundred fish were sampled from each of these farms: 50 specimens during the dry season and 50 during the rainy season.
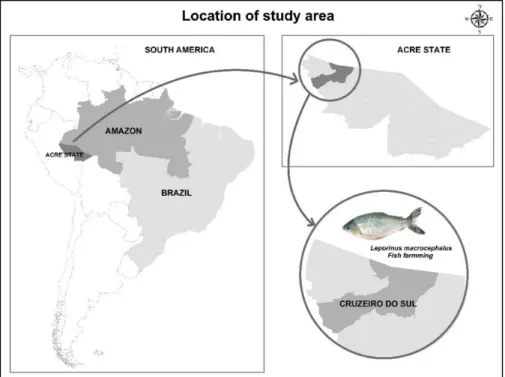
The fish were collected from a single reservoir in each farming system and each fish corresponded to one experimental unit. These fish farms are located in the municipality of Cruzeiro do Sul, State of Acre, Brazil (07° 37’ 52”S and 72° 40’ 12”W)
(Figure 1). The characteristics of the fish farms assessed is presented in Table 1. This study was authorized by the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA, license nº. 396871-1/2013).
Water quality
Water quality was measured during sample collection, always at the same time. Dissolved oxygen (O2D), hydrogen potential (pH), water temperature (T°C) and electric conductivity (EC) were measured at the edge of the breeding area, using the HANNA
multiparameter device (Hanna Instruments, USA).
Collection, processing and identification of parasites
The fish were kept in thermal boxes and transported to the “Laboratório de Processamento de Pescado” of the Federal Institute of Acre, Campus Cruzeiro do Sul, mensured the weight (g) and lenght (cm). For collection of Monogenoidea, the gills were removed and placed in bowls containing water at 65 °C and shaken; then, alcohol was added to reach a concentration of 70%. In the laboratory, the parasites were collected with the aid of a stereoscopic microscopic and stored in the same fixative. The Digenea were cold fixed in AFA (2% glacial acetic acid, 3% formaldehyde, and 95% of 70o alcohol), under light cover
glass pressure. Nematoda were rinsed in 0.7% NaCl solution and fixed in hot AFA. The material was taken to the “Laboratório de Helmintos Parasitos de Peixes” (LHPP), at the Oswaldo Cruz Institute in Rio de Janeiro and the helminths were processed for light microscope studies.
Some specimens of Monogenoidea were mounted unstained in Hoyer’s medium for study of the sclerotized parts and others were stained with Gomori’s trichrome and mounted in Canada balsam. The Digenea were stained with Langeron alcoholic acid carmine, dehydrated by means of alcohol series, cleared using beechwood creosote and mounted in Canada balsam as permanent slides. Nematoda were clarified and mounted in semi-permanent slides in phenol 50%. Specimens were studied on a light microscope Zeiss Axioscope 2 from the LHPP.
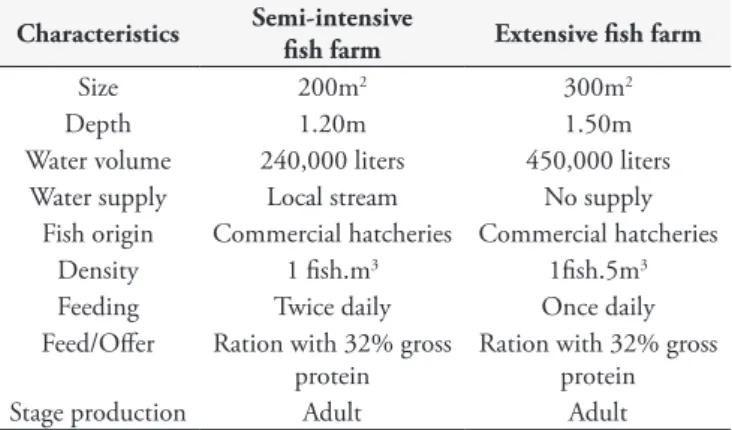
Table 1. Characteristics of fish farms systems. Characteristics Semi-intensive
fish farm Extensive fish farm
Size 200m2 300m2
Depth 1.20m 1.50m
Water volume 240,000 liters 450,000 liters
Water supply Local stream No supply
Fish origin Commercial hatcheries Commercial hatcheries
Density 1 fish.m3 1fish.5m3
Feeding Twice daily Once daily
Feed/Offer Ration with 32% gross protein
Ration with 32% gross protein
Data analysis
The total length (Lt) and total weight (Wt) were obtained and data were used to estimate the relative condition factor (Kn) of the fish. The weight-length relation was calculated using the expression P = a Cb, where P = total weight, C = total length, a = intercept
and b = angular coefficient (CREN, 1951). The coefficients a and b were estimated after logarithmic transformation of the
weight and length data and were later used in the calculation of the theoretically expected values of total weight (We), through the formula We = a.Ltb. To determine the value of We, the Kn of
the animals were calculated, in which Kn=Wt/We.
The data on total length, weight and condition factor were subjected to the assumptions of normality of residues through the Shapiro & Wilk (1965) test. Variance homogeneity was assessed through the Bartlett (1937) test. Differences between seasons were evaluated through ANOVA. Possible differences in water quality parameters between the seasons were also analyzed through ANOVA. The statistical significance level was taken to be p ≤ 0.05.
Parasite prevalence, mean abundance and intensity of infection were calculated according to Bush et al. (1997). Differences in parasite prevalences between the seasons were evaluated using the Chi-Square test (χ2) with the Yates correction (RÓZSA et al., 2000).
For the species that occurred on both seasons and parasitizing more than one host specimen, the variation of mean abundance and mean intensity of infection were compared through the Mann-Whitney U-test (ZAR, 2010). The Spearman´s correlation
coefficient (rs) was used to determine possible correlations between
mean abundance and total length according to seasonality and was considered significant when presented a p≤0.05.
Results
Water quality
Table 2 presents the water quality parameters per season. At the semi-intensive fish farm, the water temperature was significantly different between the seasons, with a higher mean during the dry season (p = 0.01), while dissolved oxygen reached higher levels during the rainy season (p = 0.03). Hydrogen potential (pH) and electric conductivity did not vary seasonally. At the extensive fish farm, a significant difference was observed in relation to water temperature (p = 0.01), which was higher during the dry season, while for the other parameters there were no significant differences.
Fish biometry
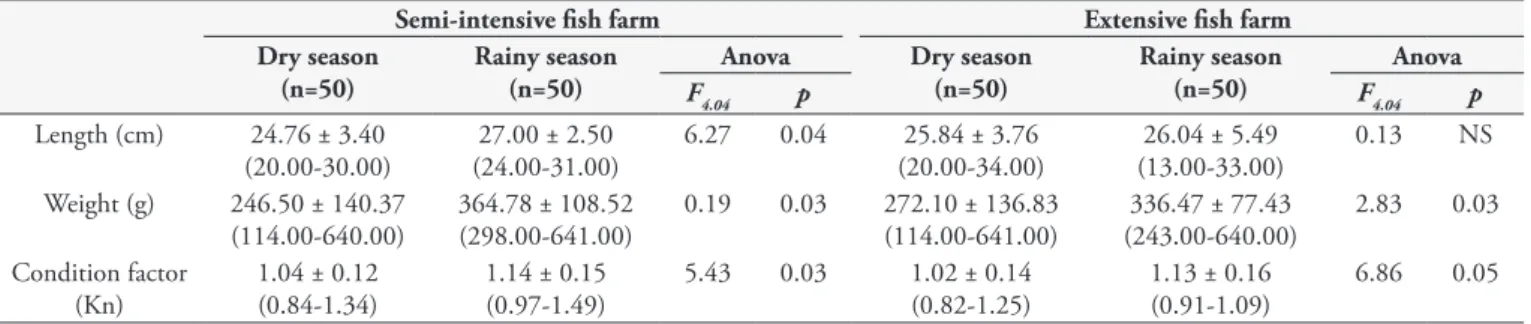
The biometric characteristics of the fish presented seasonal differences between the two farms, except regarding the total length of fish from the extensive system (Table 3). Fish from the semi-intensive fish farm were significantly larger (p = 0.04) and heavier (p = 0.03), and presented a higher condition factor (p = 0.03) during the rainy season. Similarly, in the extensive fish farm, the fish were also heavier (p = 0.03) and presented a higher condition factor (p = 0.05) during the rainy season, but with no significant difference regarding total length.
A positive and significant correlation was observed at the semi-intensive fish farm during the dry season between total length and mean abundance of parasites (rs = 0.36; p = 0.04). At the extensive fish farm, no such correlation was found (rs = 0.72; p = 0.06).
Parasites
In this study, the following species were identified:
Urocleidoides paradoxus Kritsky, Thatcher and Boeger, 1986; Urocleidoides eremitus Kritsky, Thatcher and Boeger, 1986; Jainus leporini Abdallah, Azevedo and Luque, 2012; Tereancistrum parvus Kritsky, Thatcher and Kayton, 1980; Tereancistrum paranaensis Karling, Lopes,
Takemoto and Pavanelli, 2014 e Kritskyia eirasi Kritsky, Thatcher and Kayton, 1980; Dactylogyridae sp. 1; Dactylogyridae sp. 2; Dactylogyridae sp. 3; Microcotyle sp.; Prosthenhystera obesa (Diesing,
1850) Travassos, 1922; Procamallanus (Spirocamallanus) inopinatus
Travassos, Artigas and Pereira, 1928; Rhabdochona (Rhabdochona)
acuminata (Molin, 1860); Goezia leporini Martins and Yoshitoshi, 2003; and Brevimulticaecum sp. (larva).
Six species were common to the two fish farms and occurred in the two seasons: U. paradoxus, U. eremitus, T. parvus, P. (S.) inopinatus,
R. (R.) acuminata and G. leporini. In the semi-intensive fish farm two species were observed only in the dry season: T. paranaensis e
K. eirasi, and three species were identified only in the rainy season:
J. leporini, Dactylogyridae sp. 1 and Dactylogyridae sp. 2. In the
extensive fish farm, only in the rainy season were recorded J. leporini, Dactylogyridae sp. 1, Dactylogyridae sp. 2, Dactylogyridae sp. 3,
Microcotyle sp., P. obesa and Brevimulticaecum sp. (larva).
Parasite descriptors
At the semi-intensive fish farm, there was a significant difference in the general prevalence of infection between the seasons (χ2 = 56.54;
p < 0.001). The hosts presented higher levels of infection during the dry season (P = 96%) than during the rainy season (P = 32%). Considering the parasitism rates of the species that occurred during both seasons (Table 4), U. paradoxus (p = 0.04), P. (S.)
inopinatus (p < 0.001), R. (R.) acuminata (p < 0.001) and G. leporini
(p < 0.001) presented seasonal variation of their prevalence, with higher rates during the dry season.
Regarding mean abundance, P. (S.) inopinatus (p < 0.001) was more frequent during the dry season, while U. paradoxus
(p = 0.02), Rhabdochona (R.) acuminata (p < 0.001) and G. leporini
(p < 0.001) were more abundant during the rainy season (Table 4).
P. (S.) inopinatus (p < 0.001) presented higher mean intensity of infection during the rainy season, while U. paradoxus (p < 0.001),
T. parvus (p = 0.02), R. (R.) acuminata (p < 0.001) and G. leporini
(p < 0.001) presented higher values during the dry season (Table 4).
Urocleidoides eremitus did not present significant differences between the seasons in relation to any of the indexes analyzed.
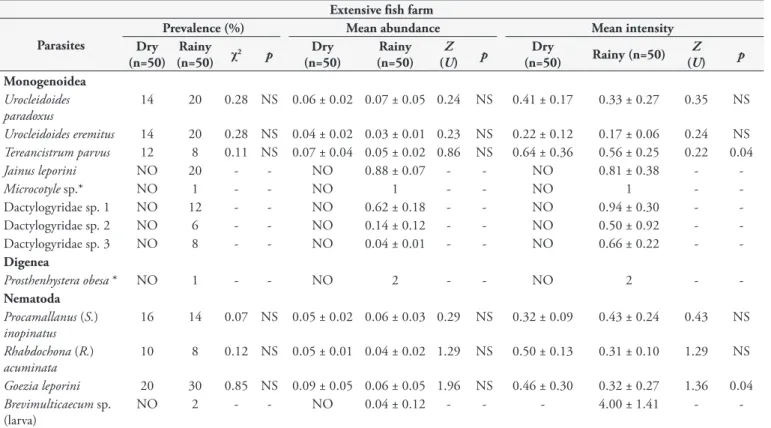
The general prevalence of infection in the extensive fish farm was 20% during the dry season and 19% during the rainy season, which did not represent a significant difference (χ2 = 0.06;
p = 0.09). For the species that occurred in both seasons, there was no significant seasonal variation in either prevalence or mean abundance (Table 5). However, T. parvus (p = 0.04) and G. leporini
(p = 0.04) were the only species that presented increased mean intensity of infection during the dry season (Table 5).
Discussion
The incidence of diseases in fish farms can be correlated with abiotic factors regarding to the environment and biotic factors related to the host, and it may be influenced by seasonality throughout the fish production cycle. According to Tavares-Dias et al. (2014), seasonal occurrences of parasites may signal periods of epizootic outbreaks, thus demonstrating the importance of understanding this variability, in order to prevent economic losses.
Table 2. Parameters of water quality in dry and rainy seasons of fish farms of Leporinus macrocephalus.
Parameters
Semi-intensive fish farm Extensive fish farm
Dry season Rainy season Anova Dry season Rainy season Anova
F4.17 p F4.17 p
Dissolved oxygen (mg.L-1) 4.85 ± 0.42 5.40 ± 0.41 8.04 0.03 5.09 ± 041 5.51 ± 0.42 3.80 NS
pH 5.79 ± 0.53 5.64 ± 0.52 0.49 NS 5.71 ± 0.56 5.73 ± 0.44 0.01 NS
Temperature (ºC) 26.61 ± 1.13 22.22 ± 0.97 79.18 0.01 24.46 ± 1.46 21.27 ± 1.06 42,06 0.01 Eletric conductivity (µS/cm) 16.23 ±3.15 17.87 ± 3.99 1.02 NS 16.02 ± 3.23 19.07 ± 4.09 3.33 NS Note: (NS) Not significant. (p ≤ 0.05) indicate significant difference between seasons.
Table 3. Means, standard deviation (minimum and maximum values in parentheses) of length, weight and factor condition (Kn) of fishes collected in dry and rainy seasons in fish farms in Acre, Brazil.
Semi-intensive fish farm Extensive fish farm
Dry season (n=50)
Rainy season (n=50)
Anova Dry season
(n=50)
Rainy season (n=50)
Anova
F4.04 p F4.04 p
Length (cm) 24.76 ± 3.40 (20.00-30.00)
27.00 ± 2.50 (24.00-31.00)
6.27 0.04 25.84 ± 3.76 (20.00-34.00)
26.04 ± 5.49 (13.00-33.00)
0.13 NS
Weight (g) 246.50 ± 140.37 (114.00-640.00)
364.78 ± 108.52 (298.00-641.00)
0.19 0.03 272.10 ± 136.83 (114.00-641.00)
336.47 ± 77.43 (243.00-640.00)
2.83 0.03
Condition factor (Kn)
1.04 ± 0.12 (0.84-1.34)
1.14 ± 0.15 (0.97-1.49)
5.43 0.03 1.02 ± 0.14 (0.82-1.25)
1.13 ± 0.16 (0.91-1.09)
6.86 0.05
Table 4. Seasonal variation of prevalence (%), mean abundance (mean ± standard deviation) and mean intensity (mean ± standard deviation) of parasitic helminths of Leporinus macrocephalus in the semi-intensive fish farm in the state of Acre.
Semi-intensive fish farm
Parasites
Prevalence (%) Mean abundance Mean intensity
Dry (n=50)
Rainy
(n=50) χ2 p
Dry (n=50)
Rainy (n=50)
Z
(U) p
Dry (n=50)
Rainy (n=50)
Z
(U) p
Monogenoidea Urocleidoides paradoxus
36 18 4.11 0.04 0.07 ± 0.04 0.10 ± 0.02 2.23 0.02 0.57 ± 0.15 0.20 ± 0.11 2.27 <0.001
Urocleidoides eremitus
30 18 1.37 NS 0.06 ± 0.02 0.03 ± 0.01 2.38 NS 0.18 ± 0.08 0.20 ± 0.09 2.82 NS
Tereancistrum parvus
20 18 0.02 NS 0.03 ± 0.01 0.04 ± 0.01 0.76 NS 0.19 ± 0.09 0.11 ± 0.04 1.32 0.02
Tereancistrum paranaensis
6 NO - - 0.06 ± 0.19 NO - - 0.43 ± 0.18 NO -
-Kritskyia eirasi 4 NO - - 0.06 ± 0.02 NO - - 3.00 ± 1.00 NO -
-Jainus leporini NO 6 - - NO 0.05 ± 0.02 - - - 1.50 ± 0.50 -
-Dactylogyridae sp.1 NO 2 - - NO 0.08 ± 0.40 - - - 1.66 ± 0.54 -
-Dactylogyridae sp.2 NO 4 - - NO 0.10 ± 0.02 - - - 1.40 ± 0.08 -
-Nematoda Procamallanus (S.)
inopinatus
96 32 41.70 <0.001 0.14 ± 0.07 0.08 ± 0.06 2.49 <0.001 0.32 ± 0.09 0.43 ± 0.24 24.00 <0.001
Rhabdochona (R.)
acuminata
56 10 15.76 <0.001 0.07 ± 0.04 0.09 ± 0.02 1.63 <0.001 0.50 ± 0.13 0.31 ± 0.10 3.30 <0.001
Goezia leporini 80 20 33.64 <0.001 0.18 ± 0.04 0.22 ± 0.11 0.92 <0.001 0.46 ± 0.30 0.32 ± 0.27 21.00 <0.001 Note: (NO) Not observed. (NS) Not significant. (p ≤ 0.05) indicate significant difference between seasons.
Table 5. Seasonal variation of prevalence (%), mean abundance (mean ± standard deviation) and mean intensity (mean ± standard deviation) of parasitic helminths of Leporinus macrocephalus in the extensive fish farm in the state of Acre.
Extensive fish farm
Parasites
Prevalence (%) Mean abundance Mean intensity
Dry (n=50)
Rainy
(n=50) χ2 p
Dry (n=50)
Rainy (n=50)
Z
(U) p
Dry
(n=50) Rainy (n=50)
Z
(U) p Monogenoidea
Urocleidoides paradoxus
14 20 0.28 NS 0.06 ± 0.02 0.07 ± 0.05 0.24 NS 0.41 ± 0.17 0.33 ± 0.27 0.35 NS
Urocleidoides eremitus 14 20 0.28 NS 0.04 ± 0.02 0.03 ± 0.01 0.23 NS 0.22 ± 0.12 0.17 ± 0.06 0.24 NS
Tereancistrum parvus 12 8 0.11 NS 0.07 ± 0.04 0.05 ± 0.02 0.86 NS 0.64 ± 0.36 0.56 ± 0.25 0.22 0.04
Jainus leporini NO 20 - - NO 0.88 ± 0.07 - - NO 0.81 ± 0.38 -
-Microcotyle sp.* NO 1 - - NO 1 - - NO 1 -
-Dactylogyridae sp. 1 NO 12 - - NO 0.62 ± 0.18 - - NO 0.94 ± 0.30 -
-Dactylogyridae sp. 2 NO 6 - - NO 0.14 ± 0.12 - - NO 0.50 ± 0.92 -
-Dactylogyridae sp. 3 NO 8 - - NO 0.04 ± 0.01 - - NO 0.66 ± 0.22 -
-Digenea
Prosthenhystera obesa * NO 1 - - NO 2 - - NO 2 -
-Nematoda Procamallanus (S.)
inopinatus
16 14 0.07 NS 0.05 ± 0.02 0.06 ± 0.03 0.29 NS 0.32 ± 0.09 0.43 ± 0.24 0.43 NS
Rhabdochona (R.)
acuminata
10 8 0.12 NS 0.05 ± 0.01 0.04 ± 0.02 1.29 NS 0.50 ± 0.13 0.31 ± 0.10 1.29 NS
Goezia leporini 20 30 0.85 NS 0.09 ± 0.05 0.06 ± 0.05 1.96 NS 0.46 ± 0.30 0.32 ± 0.27 1.36 0.04
Brevimulticaecum sp. (larva)
NO 2 - - NO 0.04 ± 0.12 - - - 4.00 ± 1.41 -
Although some authors have previously evaluated seasonal effects on the parasite community of L. macrocephalus in the state of São Paulo (TAVARES-DIAS et al., 2001; SCHALCH & MORAES, 2005), no studies of this nature have been conducted in the humid tropical climate of the Amazon, particularly in relation to this species, which was introduced to the region for farming systems.
Some species of helminths have been reported among other fish of the genus Leporinus in the reservoir behind the Nova Pontes dam in Minas Gerais and on the floodplain of the upper Paraná River (KRITSKY et al., 1980; FELTRAN et al., 2004; GUIDELLI et al., 2006; TAKEMOTO et al., 2009; AZEVEDO et al., 2010; GUIDELLI et al., 2011). Two species of helminths Rhinoxenus sp. and G. leporini have been reported parasitizing L. macrocephalus
in fish farms and in the natural environment (MARTINS & YOSHITOSHI, 2003; TAKEMOTO et al., 2009). In the present study, 15 species were identified and some of them showed seasonal variation of occurrence. T. paranaensis and K. eirasi occurred only in
the semi-intensive fish farm during the dry season, while J. leporini,
Dactylogyridae sp. 1 and Dactylogyridae sp. 2 only occurred during the rainy season. In the extensive fish farm, J. leporini,
Dactylogyridae sp. 1, Dactylogyridae sp. 2, Dactylogyridae sp. 3, Microcotyle sp., P. obesa and Brevimulticaecum sp. (larva) were
observed only during the rainy season.
The occurrence of six species in both fish farms and during both sampling periods indicated that these parasites can at least infect hosts regardless of seasonal variations and the environmental conditions of the farming systems. According to Schalch & Moraes (2005), occurrences of these species may be associated with the life cycle characteristics of these parasites, such that recurring and continuous infections are enabled.
The high prevalence of infection in the semi-intensive fish farm during the dry season (96%) was higher than that reported by Schalch & Moraes (2005) (87.2%) for the same host (L. macrocephalus) in a fish-out pond establishment in the state of São Paulo. This pattern of infection, according to these authors, follows an increase in temperature, which combined with poor environmental quality, nutritional deficiency and inadequate management, favors occurrences of parasite diseases of greater severity. In the present study, these factors also explained the high prevalence during the dry season, since the temperature was higher and oxygen levels were lower. Moreover, the stocking density of the fish in the semi-intensive fish farm (1 fish.m3) was greater
than in the extensive farm (1 fish.5 m3), which also influenced
the greater prevalence of infection in this system. According to Sanches (2008), high density promotes transmission of the infectious forms of parasites among hosts.
Several studies (TAVARES-DIAS et al., 2001; SCHALCH & MORAES, 2005; ZARGAR et al., 2012; JERÔNIMO et al., 2016; MARCHIORI et al., 2015) have suggested that natural abiotic factors such as temperature, oxygen, salinity and hydrogen ion concentration have a positive influence over parasite populations. In the Amazon region, some authors (ARAÚJO et al., 2009; MARINHO et al., 2013; DIAS et al., 2015; SILVA et al., 2016) have reported that high prevalence of parasites in fish farms was associated with low levels of dissolved oxygen in the water, high temperatures and inadequate sanitary conditions. These authors
also reported that replenishing the breeding areas with water from natural environments also influenced occurrences and levels of infection.
In the semi-intensive fish farm this also contributed towards high prevalence of infection, because the water source was a local stream, with no treatment or filtration. This was especially observed during the dry season, when there was greater loss of water due to evaporation. Although the water temperature was higher during the dry season in the extensive fish farm, higher prevalence of infection was not observed during this period, which could be related to the lower stocking density of fish in this system. Moreover, the other water quality parameters presented similar values and did not undergo significant variation between the seasons, which may have led to lower stress for the fish and better conditions for their development.
Some species of Monogenoidea demonstrated seasonal variation, with higher parasite descriptors during the dry season. Similar results were observed by Schalch & Moraes (2005) in the municipality of Guariba, São Paulo, with seasonal variations of infestation rates and higher occurrence of Monogenoidea during the summer, when temperatures were higher. According to these authors, the reproduction of this group of parasites is favored by higher temperatures, greater fish population density and lower oxygen levels in the water.
In the semi-intensive fish farm, the prevalence of U. paradoxus
in the dry season was similar to that observed by Guidelli et al. (2006) for L. lacustris and L. friderici, with prevalences of 32% and 46.1%, respectively, for specimens in the upper Paraná river floodplain. For T. parvus, these infection rates in both seasons (dry = 20%, rainy = 18%) were higher than what was observed by these authors (L. lacustris = 6.66%, L. friderici = 3.84%). However, the environmental conditions of the areas studied are different. Moreover, farming systems are known to potentially present higher prevalence of infection due to the greater concentration of fish.
Nematodes cause severe infections among farmed fish (MARTINS et al., 2004). Although damage to the hosts was not observed in the present study, G. leporini was the parasite with greatest infection rates. Moreover, this species showed high prevalence (80%) at the semi-intensive fish farm during the dry season. This nematode is a highly pathogenic parasite in
L. macrocephalus. It can negatively influence the health of this fish through reducing its growth, and in severe cases can cause death (MARTINS & YOSHITOSHI, 2003).
Procamallanus (S.) inopinatus is a common and abundant species among Brazilian fish. The parasitism rates observed in the present study were higher than those reported for other fish of the Anostomidae family (MORAVEC, 1998; FELTRAN et al., 2004; GUIDELLI et al., 2006; TAKEMOTO et al., 2009; AZEVEDO et al., 2010; GUIDELLI et al., 2011). However, farming and management conditions vary among these species, and this can influence the parasite population.
In this study, with the exception of Brevimulticaecum sp. (larva),
the highest prevalence and mean abundance rates were observed for the endohelminths P. (S.) inopinatus and G. leporini. This may
the fish, especially during the dry season in the semi-intensive fish farm, where the rates were higher.
Concerning the biometric characteristics of the fish, in both systems the weight and condition factor were higher in the rainy season, coinciding with the period of greater availability of naturally produced foods in fish farms, which provided greater accumulation of fat in this season. In addition, the water temperature was lower in the rainy season, which may have provided better environmental conditions, influencing fish development.
Also in this study, the data obtained indicated that seasonality can influence the levels of parasitism. During the dry season, the fish presented higher rates of parasitism. Although no clinical signs of disease caused by parasites were observed, the data suggest that prophylactic measures should be applied in these farms, especially during the dry season.
The data presented here can aid fish farmers in understanding the parasite dynamics in farming systems during different seasons. Thus, these data assist towards improvement of conditions in the breeding areas with regard to fish population density. This study is the first report assessing parasite seasonality for a fish species that was introduced to the Amazon region for farming. Additional studies should be pursued with the intent of improving farming conditions for this promising species in the state of Acre.
References
Aguinaga JY, Marcusso PF, Claudiano GS, Lima BT, Sebastião FA, Fernandes JB, et al. Parasitic infections in ornamental cichlid fish in the Peruvian Amazon. Rev Bras Parasitol Vet 2015; 24(1): 82-86. PMid:25909258. http://dx.doi.org/10.1590/S1984-29612014076.
Araújo CSO, Gomes AL, Tavares-Dias M, Andrade SMS, Belem-Costa A, Borges JT, et al. Parasitic infections in pirarucu fry, Arapaima gigas
Schinz, 1822 (Arapaimatidae) kept in a semi-intensive fish farm in Central Amazon, Brazil. Vet Arh 2009; 79(5): 499-507.
Azevedo RK, Abdallah VD, Luque JL. Acanthocephala, Annelida, Arthropoda, Myxozoa, Nematoda and Platyhelminthes parasites of fishes from the Guandu river, Rio de Janeiro, Brazil. Check List 2010; 6(4): 659-667. http://dx.doi.org/10.15560/6.4.659.
Bartlett MS. Properties of sufficiency and statistical tests. Proc R Soc Lond
1937; 160(1): 268-282. http://dx.doi.org/10.1098/rspa.1937.0109.
Bush AO, Lafferty KD, Lotz JM, Shostak AW. Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol 1997; 83(4): 575-583. PMid:9267395. http://dx.doi.org/10.2307/3284227.
Cren EDL. The length-weight relationship and seasonal cycle in gonad weight and condition in the perch (Perca fluviatilis). J Anim Ecol 1951; 20(2): 201-219. http://dx.doi.org/10.2307/1540.
Dias MKR, Neves LR, Marinho RGB, Pinheiro DA, Tavares-Dias M. Parasitismo em tambatinga (Colossoma macropomum x Piaractus brachypomus, Characidae) cultivados na Amazônia, Brasil. Acta Amazon
2015; 45(2): 231-238. http://dx.doi.org/10.1590/1809-4392201400974.
Feltran RB, Marçal Júnior O, Pinese JF, Takemoto RM. Prevalência, abundância, intensidade e amplitude de infecção de nematóides intestinais em Leporinus friderici (Bloch, 1794) e L. obtusidens (Valenciennes, 1836) (Pisces, Anostomidae), na represa de Nova Ponte (Perdizes, MG). Rev Bras Zoocienc 2004; 6(2): 169-179.
Figueroa SN, Nobre CA. Precipitation distribution over Central and Western Tropical South America. Climanálise 1990; 5(6): 36-45.
Guidelli G, Tavechio WLG, Takemoto RM, Pavanelli GC. Fauna parasitária de parasitária de Leporinus lacustris e Leporinus friderici (Characiformes, Anostomidae) da planície de inundação do alto rio Paraná, Brasil. Acta Sci Biol Sci 2006; 28(3): 281-290. http://dx.doi.org/10.4025/actascibiolsci. v28i3.228.
Guidelli G, Tavechio WLG, Takemoto RM, Pavanelli GC. Relative condition factor and parasitism in anostomid fishes from the floodplain of the Upper Paraná River, Brazil. Vet Parasitol 2011; 177(1-2): 145-151. PMid:21176864. http://dx.doi.org/10.1016/j.vetpar.2010.11.035.
Jerônimo GT, Pádua SB, Ventura AS, Gonçalves ELT, Ishikawa MM, Martins ML. Parasitological assessment in the hybrid surubim (Pseudoplatystoma reticulatum x P. corruscans), with uncommon occurrence of Monogenea parasites. Rev Bras Parasitol Vet 2016; 25(2): 179-186. PMid:27334818. http://dx.doi.org/10.1590/S1984-29612016037.
Kritsky DC, Thatcher VE, Kayton RJ. Neotropical Monogenoidea. 3. Five new species from South America with the proposal of Tereancistrum
gen. n. and Trinibaculum gen. n. (Dactylogyridae: Ancyrocephalinae).
Acta Amazon 1980; 10(2): 411-417. http://dx.doi.org/10.1590/1809-43921980102411.
Marchiori NC, Gonçalves ELT, Tancredo KR, Pereira-Júnior J, Garcia JRE, Martins ML. Effect of water temperature and salinity in oviposition, hatching success and infestation of Aphanoblastella mastigatus (Monogenea, Dactylogyridae) on Rhamdia quelen.Braz J Biol 2015;75(4 Suppl 1): 245-252. PMid:26628218. http://dx.doi.org/10.1590/1519-6984.14014.
Marinho RGB, Tavares-Dias M, Dias-Grigório MKR, Neves LR, Yoshioka ETO, Boijink CL, et al. Helminthes and protozoan of farmed pirarucu (Arapaima gigas) in eastern Amazon and host-parasite relationship. Arq Bras Med Vet Zootec 2013; 65(4): 1192-1202. http://dx.doi.org/10.1590/ S0102-09352013000400035.
Martins ML, Tavares-Dias M, Fujimoto RY, Onaka EM, Nomura DT. Haematological alterations of Leporinus macrocephalus (Osteichtyes: Anostomidae) naturally infected by Goezia leporini (Nematoda: Anisakidae) in fish pond. Arq Bras Med Vet Zootec 2004; 56(5): 640-646. http:// dx.doi.org/10.1590/S0102-09352004000500011.
Martins ML, Yoshitoshi ER. A new nematode species Goezia leporini n. sp. (Anisakidae) from cultivated freshwater fish Leporinus macrocephalus
(Anostomidae) in Brazil. Braz J Biol 2003; 63(3): 497-506. PMid:14758709. http://dx.doi.org/10.1590/S1519-69842003000300016.
Moravec F. Nematodes of freshwater fishes of the neotropical region. Praha: Academy of Sciences of the Czech Republic; 1998.
Pavanelli GC, Takemoto RM, Eiras JC. Parasitologia de peixes de água doce do Brasil. Maringá: Eduem; 2013.
Rózsa L, Reiczigel J, Majoros G. Quantifying parasites in samples of hosts. J Parasitol 2000; 86(2): 228-232. PMid:10780537. http://dx.doi. org/10.1645/0022-3395(2000)086[0228:QPISOH]2.0.CO;2.
Sanches EG. Controle de Neobenedenia melleni (MacCallum,1927) (Monogenea: Capsalidae) em garoupa-verdadeira, Epinephelus marginatus
(Lowe, 1834), cultivada em tanques-rede. Rev Bras Parasitol Vet 2008; 17(3): 145-149. PMid:19245760. http://dx.doi.org/10.1590/S1984-29612008000300005.
Shapiro SS, Wilk MB. An analysis of variance test for normality. Biometrika
1965; 52(3-4): 591-611. http://dx.doi.org/10.1093/biomet/52.3-4.591.
Silva AEP, Angelis CF, Machado LAT, Waichaman AV. Influência da precipitação na qualidade da água do Rio Purus. Acta Amaz 2008; 38(4): 733-742. http://dx.doi.org/10.1590/S0044-59672008000400017.
Silva MT, Pinto GP, Cavalcante PHO, Santos FGA, Moutinho VAC, Portes CS. Helminth community structure of Arapaima gigas in semi-intensive and semi-intensive fish farming systems in the southwestern Brazilian Amazon. Neotropic Helminthol 2016; 10(2): 219-231.
Takemoto RM, Pavanelli GC, Lizama MAP, Lacerda ACF, Yamada FH, Moreira LHA, et al. Diversity of parasites of fish from the Upper Paraná River floodplain, Brazil. Braz J Biol 2009;69(2 Suppl): 691-705. PMid:19738975. http://dx.doi.org/10.1590/S1519-69842009000300023.
Tavares-Dias M, Moraes FT, Martins ML, Kronka SN. Fauna parasitária de peixes oriundos de “pesque-pagues” do município de Franca, São Paulo, Brasil. II. Metazoários. Rev Bras Zool 2001;18(suppl 1): 81-95. http://dx.doi.org/10.1590/S0101-81752001000500006.
Tavares-Dias M, Oliveira MSB, Gonçalves RA, Silva LMA. Ecology and seasonal variation of parasites in wild Aequidens tetramerus, a Cichlidae from the Amazon. Acta Parasitol 2014; 59(1): 158-164. PMid:24570063. http://dx.doi.org/10.2478/s11686-014-0225-3.
Tavares-Dias M, Schalch SHC, Martins ML, Silva ED, Moraes FR, Perecin D. Hematologia de teleósteos brasileiros com infecção parasitária. I. Variáveis do Leporinus macrocephalus Garavelo e Britski, 1988 (Anostomidae) e
Piaractus mesopotamicus Holmberg, 1887 (Characidae). Acta Scientiarum
1999; 21(2): 337-342. http://dx.doi.org/10.4025/actascibiolsci.v21i0.4440.
Vasconcelos HCG, Tavares-Dias M. Influência da sazonalidade na infestação de Ergasilus turucuyus (Copepoda: Ergasilidae) em Acestrorhynchus falcirostris e Hemiodus unimaculatus (Osteichthyes: Characiformes) do Reservatório Coaracy Nunes, estado do Amapá, Brasil. Biota Amaz 2014; 4(1): 106-110. http://dx.doi.org/10.18561/2179-5746/biotaamazonia. v4n1p106-110.
Vital JF, Varella AMB, Porto DB, Malta JCO. Sazonalidade da fauna de metazoários de Pygocentrus nattereri (Kner, 1858) no lago Piranha (Amazonas, Brasil) e a avaliação de seu potencial como indicadora da saúde do ambiente. Biota Neotrop 2011; 11(1): 199-204. http://dx.doi. org/10.1590/S1676-06032011000100021.
Zago AC, Franceschini L, Garcia F, Schalch SHC, Gozi KS, Silva RJ. Ectoparasites of Nile tilapia (Oreochromis niloticus) in cage farming in a hydroelectric reservoir in Brazil. Rev Bras Parasitol Vet 2014; 23(2): 171-178. PMid:25054495. http://dx.doi.org/10.1590/S1984-29612014041.
Zanolo R, Yamamura MH. Parasitas em tilápias-do-nilo criadas em sistema de tanques-rede. Semina: Ciênc Agrár 2006; 27(2): 281-288. http://dx.doi.org/10.5433/1679-0359.2006v27n2p281.
Zar JH. Biostatistical analysis. New Jersey: Prentice Hall; 2010.
Zargar UR, Chishti MZ, Yousuf AR, Fayaz A. Infection level of monogenean gill parasite, Diplozoon kashmirensis (Monogenea, Polyopisthocotylea) in the Crucian Carp, Carassius carassius from lake ecosystems of an altered water quality: What factors do have an impact on the Diplozoon infection?