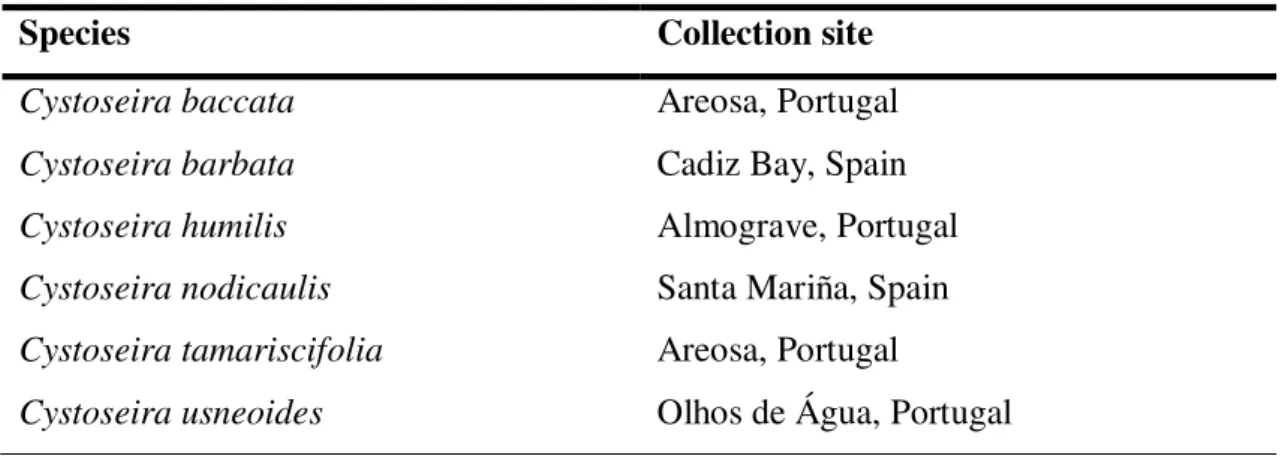
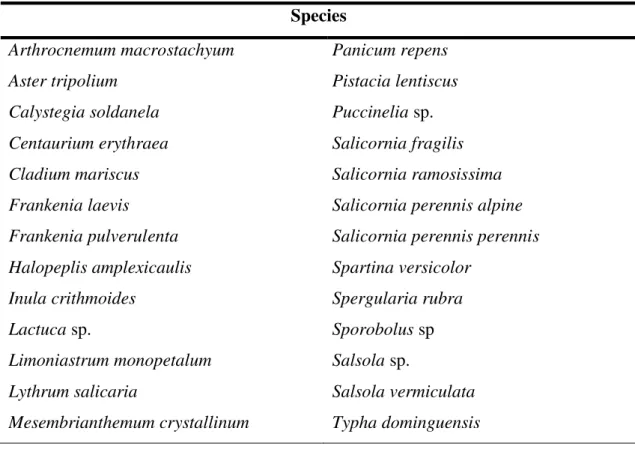
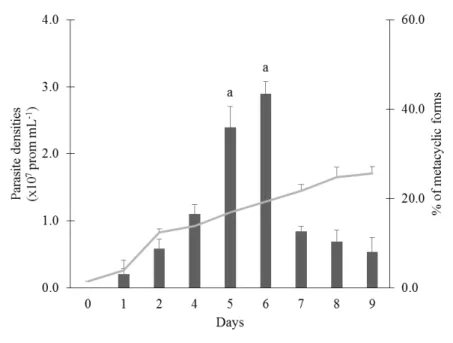
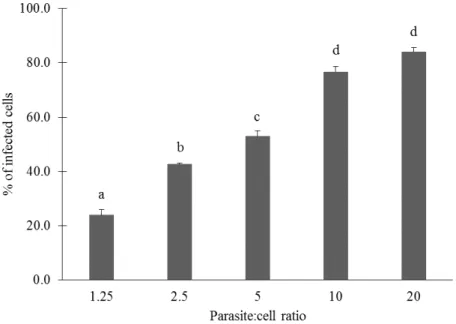
Optimization of screening methods for the evaluation of the antileishmanial potential of halophytes and macroalgae from the Iberian Coast
Texto
Imagem
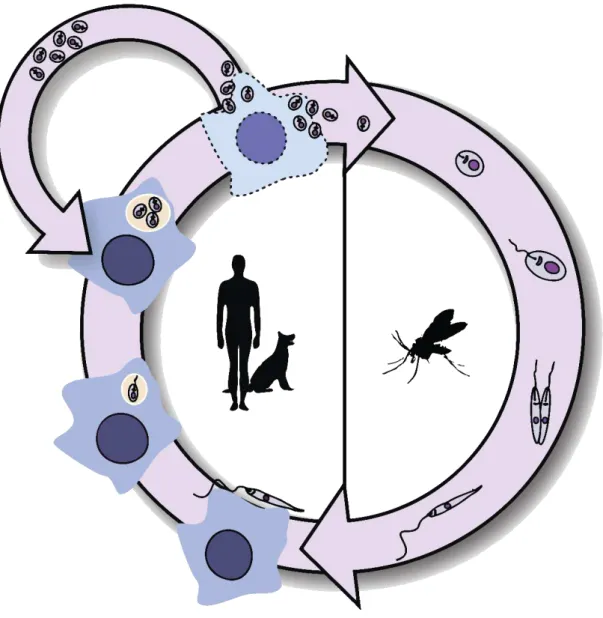
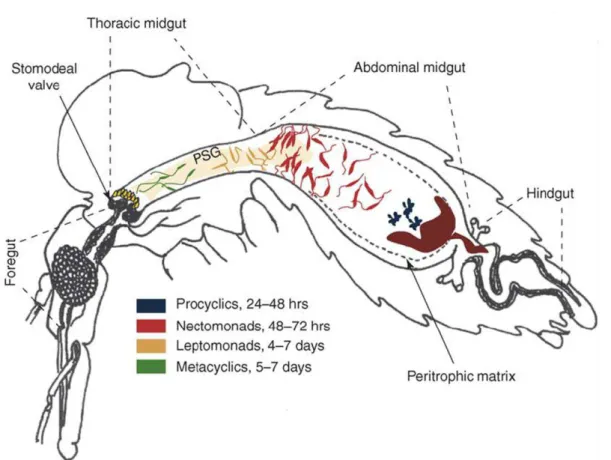
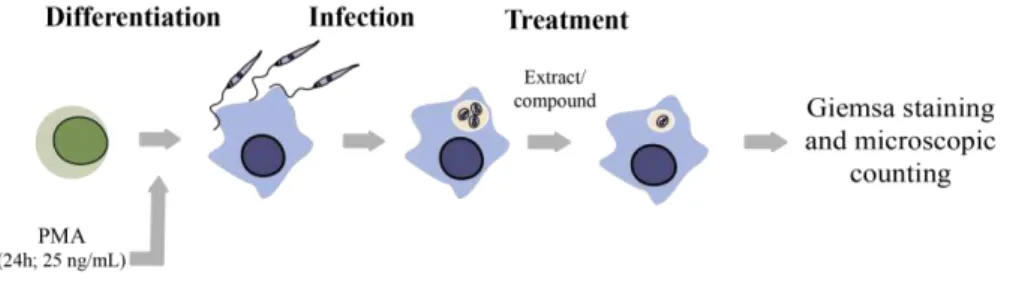
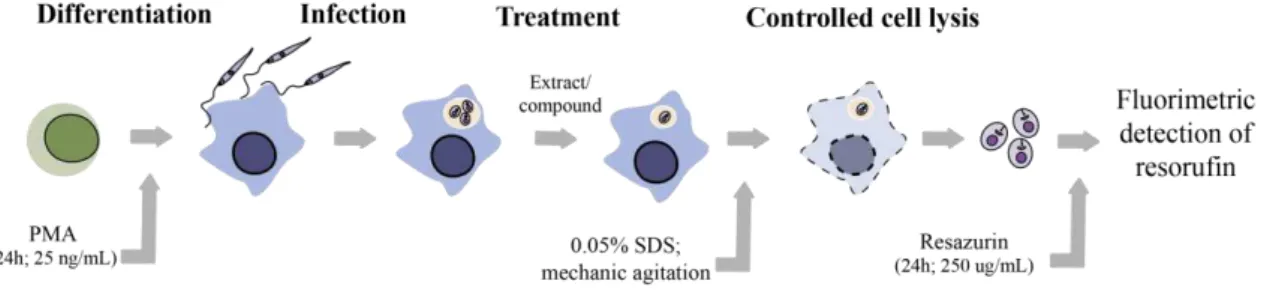
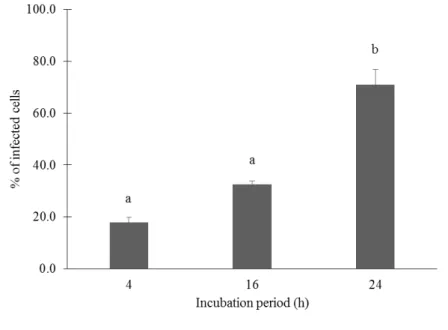
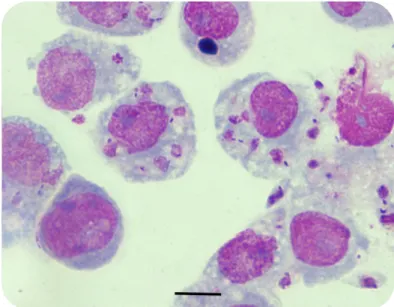
Documentos relacionados
Na hepatite B, as enzimas hepáticas têm valores menores tanto para quem toma quanto para os que não tomam café comparados ao vírus C, porém os dados foram estatisticamente
The irregular pisoids from Perlova cave have rough outer surface, no nuclei, subtle and irregular lamination and no corrosional surfaces in their internal structure (Figure
i) A condutividade da matriz vítrea diminui com o aumento do tempo de tratamento térmico (Fig.. 241 pequena quantidade de cristais existentes na amostra já provoca um efeito
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
didático e resolva as listas de exercícios (disponíveis no Classroom) referentes às obras de Carlos Drummond de Andrade, João Guimarães Rosa, Machado de Assis,
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
Alguns ensaios desse tipo de modelos têm sido tentados, tendo conduzido lentamente à compreensão das alterações mentais (ou psicológicas) experienciadas pelos doentes
