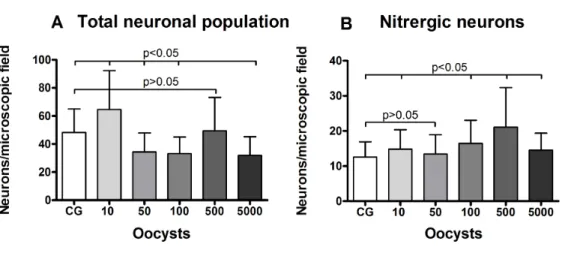
Different inoculum loads of Toxoplasma gondii induce reduction of myenteric neurons of the rat colon
Texto
Imagem

Documentos relacionados
didático e resolva as listas de exercícios (disponíveis no Classroom) referentes às obras de Carlos Drummond de Andrade, João Guimarães Rosa, Machado de Assis,
A concepção de ação social torna-se central dentro desta dinâmica, já que é orientada em uma relação com o outro (Weber, 2014). Weber argumenta que a grande passagem
Despercebido: não visto, não notado, não observado, ignorado.. Não me passou despercebido
The fourth generation of sinkholes is connected with the older Đulin ponor-Medvedica cave system and collects the water which appears deeper in the cave as permanent
The irregular pisoids from Perlova cave have rough outer surface, no nuclei, subtle and irregular lamination and no corrosional surfaces in their internal structure (Figure
i) A condutividade da matriz vítrea diminui com o aumento do tempo de tratamento térmico (Fig.. 241 pequena quantidade de cristais existentes na amostra já provoca um efeito
ABSTRACT Alterations caused by a genotype III strain of Toxoplasma gondii were assessed with respect to the number and the morphometry of the myenteric neurons
Ainda assim, sempre que possível, faça você mesmo sua granola, mistu- rando aveia, linhaça, chia, amêndoas, castanhas, nozes e frutas secas.. Cuidado ao comprar
