w ww . e l s e v i e r . c o m / l o c a t e / b j p
Original
Article
Leaf
morphology
and
venation
of
Psidium
species
from
the
Brazilian
Savanna
Elaine
F.
Oliveira
a,
Debborah
G.
Bezerra
a,
Mirley
L.
Santos
a,
Maria
H.
Rezende
b,
Joelma
A.M.
Paula
a,∗ aUniversidadeEstadualdeGoiás,CampusAnápolisdeCiênciasExataseTecnológicas,Anápolis,GO,BrazilbDepartamentodeBotânica,InstitutodeCiênciasBiológicas,UniversidadeFederaldeGoiás,Goiânia,GO,Brazil
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received1December2016 Accepted6March2017 Availableonline14July2017
Keywords: “Arac¸á” Diaphanization Leafvenation Taxonomy Qualitycontrol
a
b
s
t
r
a
c
t
TheBrazilianSavannabiomehasthelargestplantdiversityamong savannasworldwideandis the predominantbiomeinGoiásstate,Brazil.AmongplantspeciespreviouslycataloguedintheBrazilian Savanna,theMyrtaceaefamilyhasreceivedattention,astheseplantsshowgreateconomicpotentialfor itsmedicinalproperties,foodandecologicalimportance,highlightinginthiscontextthePsidiumgenus. Inordertocontributetothedifferentiationofproblematictaxonomicgroups,suchasMyrtaceae,andto qualitycontrolofitsplantmaterial,morphologicalandvenationleafstudiesoffourspeciesofPsidium
wereperformed.Forthispurpose,leafsamplesofPsidiumfirmumO.Berg.,P.myrsinitesDC.,P. laruot-teanumCambes.,andP.guineenseSw.werecollectedfromGoiásStateUniversity,AnápolisAirBase,and SerradeCaldasNovasStateParkandsubmittedtoclassicaltechniquesformorphologicalandleaf vena-tioncharacterizations.TheresultsshowedthatP.firmumpresentsbrochidodromoussecondaryveins, marginallastvenationofthefimbrialtype,anabaxialsurfacewithagroovedmidrib,flatsecondaryveins onbothsides,anapexobtusetomucronate,andaroundedbase.P.guineensepresentstrichomesonboth surfaces,agroovedmidribontheadaxialsurfaceandaprominentmidribontheabaxialsurface,which distinguishesthisspeciesfromallotherMyrtaceaespeciesexaminedinthepresentstudy.ThespeciesP. guineenseandP.firmumpresentedasetofdifferentialleafcharacteristicsamongtheotherstaxaofthe genus,clearlyseparatingtheseplantsinthemorphologicalidentificationkey.
©2017SociedadeBrasileiradeFarmacognosia.PublishedbyElsevierEditoraLtda.Thisisanopen accessarticleundertheCCBY-NC-NDlicense(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Introduction
TheBrazilianSavannaisconsideredahotspot(priorityarea)for globalbiodiversityconservation(Myersetal.,2000);forhostingan estimatednumberof837birdspecies,161mammalspecies,150 amphibianspecies,and120reptilespecies(SilvaandBates,2002); andforhighconcentrationsofendemicplants(Mittermeieretal.,
2005).
Amongthegreat diversity oftheBrazilian Savannaendemic fruits,wehighlightthosebelongingtotheMyrtaceaefamily.The ecologicalimportanceofthisfamilyindifferentbiomesofBrazil hasbeenconfirmedinseveralfloristic,phylogenetic,and phytoso-ciologicalstudies.AccordingtoOliveira-FilhoandFontes(2000), inboththeBrazilianSavannaandAtlanticForest,theMyrtaceae familyhasagreatdiversityofspecies,representing10–15%ofthe vegetalcoverinthesebiomes.
∗ Correspondingauthor.
E-mail:[email protected](J.A.Paula).
One of most comprehensive surveys of the Brazilian Myr-taceaelisted1726species(Berg,1859).AccordingtoLandrumand
Kawasaki(1997),thisfamilyisrepresentedby23generaand1000
species.Amongtheseplants,696speciesareexclusivetoBrazil
(Arantes,1997).InamorerecentstudyinBrazil,Sobraletal.(2013)
recorded23generaandapproximately997species.IntheBrazilian Savanna,Mendonc¸aetal.(2007)listed14generaand211species. AmongtheplantspeciesoftheMyrtaceaefamilypreviously cat-aloguedinopenareasoftheBrazilianSavanna,thePsidiumgenusis distinguishedbyitsgreateconomicpotential,reflectingitsfoodand pharmacologicaluses(SouzaandLorenzi,2008).InBrazilPsidium guajavaL.standsoutasthespeciesofgreatesteconomicinterest ofthePsidiumgenus.However,therehasbeengrowinginterestin the“arac¸azeiros”(Medinaetal.,2011),whichencompassseveral speciesofPsidium.IntheCentralWestregionofBrazil,inaddition toPsidiumguineense,thefollowing“arac¸ás”specieshavealsobeen observed:PsidiumlaruotteanumCambess.,PsidiumfirmumO.Berg., PsidiummyrsinitesDC.,Psidiumsartorianum,andPsidiumsalutare
(Frazonetal.,2009).
WithregardtothemedicinalpotentialofSavanna“arac¸ás”,a recentethnobotanicalsurveyhasindicatedtheuseofleavesand
http://dx.doi.org/10.1016/j.bjp.2017.03.005
Caldas Novas
Caldas Novas State Park
20 Km 10 5 0
Fig.1.MapofBrazilshowingthelocationsofGoiásState,AnápolisandCaldasNovasmunicipalitiesandPsidiumspp.collectionsites.
Table1
Geographiclocation,dateofcollection,geographiccoordinates,andvouchernumberofPsidiumspeciescollectedintheSavannaoftheGoiásstate.
Species Local Collectiondate Geographiccoordinates
Longitude/latitude/altitude(m) Vouchernumber
PsidiumfirmumO.Berg. GSU 08/2013
02/2014 08/2014
16◦22′53.977′′/48◦56′41.779′′/1098 9211a
PsidiummyrsinitesDC. GSU 06/2013
02/2014 08/2014
16◦22′42.058′′/48◦56′40.488′′/1099 9212a
AAB 11/2013
06/2014 08/2014
16◦12′52.991′′/48◦57′55.868′′/1104 9213a
Psidiumlaruotteanum Cambess.
GSU 06/2013
02/2014 08/2014
16◦22′46.885′′/48◦56′39.774′′/1100 9214a
AAB 11/2013
06/2014 08/2014
16◦12′56.088′′/48◦57′56.486′′/1114 9215a
SCNSP 08/2013
11/2013 02/2014 06/2014
17◦28′12.396′′/48◦24′3.604′′/982 9218a
PsidiumguineenseSw. AAB 11/2013
06/2014 08/2014
16◦12′45.507′′/48◦57′23.651′′/1104 9217a
GSU,CampusAnápolisoftheGoiásStateUniversity;SCNSP,SerradeCaldasNovasStatePark;AAB,AnápolisAirBase. aHerbariumoftheGoiásStateUniversity(HGSU).
shootsbyresidentsofurbanareas,settlementsand“quilombola” communitiesoftheGoiásSavannaforthetreatmentof
dysenter-ies(Campos, 2010).Datafromthescientific literaturereinforce
thepharmacologicalpotentialof“arac¸ás”,sincerelevant biologi-calactivitieshavebeenattributedtothefruitsandleavesofthese species,such as: antioxidant,antimicrobial,antiproliferative on humantumorcellsand increasedsleepinducedbyketaminein
mice(Fauthetal.,2002;Corrêaetal.,2011;Medinaetal.,2011;
Voss-Rechetal.,2011;Oliveiraetal.,2012;Patel,2012).
Severalresearchers,includingMcVaugh (1968)and Barroso
et al. (1991), have described the taxonomic problems
involv-ingMyrtaceae, stating that this family isa complex taxonomic group. According to Costa (2004), the difficulty of identifying
BrazilianMyrtaceaecanbeattributedtothespeciation,resulting fromhybridizationandpolyploidy,andthelackofmorphological andanatomicalstudiesforabetterdelimitationofthetaxa.The Psidiumspecies,in particular,havea greatphenotypicplasticity duetothedifferentenvironmentalpressurestowhich theyare subject,causingdifficultiesintheidentificationanddelimitationof thespecies(Costa,2009).Theyarefoundindiverseenvironments suchas,semi-desertandrestingas,withcalcareousandsandysoils, amongothers(Brandãoetal.,2002).
Fig.2.Psidiumspp.leaves:(A)P.firmum,adaxialandabaxialsurfaces;(a1)P.firmumdiaphanizedleaf–brochidodromoussecondaryvenationpattern;(B)P.myrsinites, adaxialandabaxialsurfaces;(b1)P.myrsinitesdiaphanizedleaf–camptodromous–brochidodromoussecondaryvenationpattern;(C)P.laruotteanum,adaxialandabaxial surfaces;(c1)P.laruotteanumdiaphanizedleaf–camptodromous–brochidodromoussecondaryvenationpattern;(D)P.guineense,adaxialandabaxialsurfaces;(d1)P. guineensediaphanizedleaf–camptodromous–brochidodromoussecondaryvenationpattern.
Amongthefoliarmorphologicalcharacteristicscommontothese species, wecan highlightthe simple, opposite,usually crossed, amphiorhypoestomaticleaves,withabundanttectortrichomeson theabaxialsurfaceandrareintheadaxialsurface,and camptodro-mousorbrochidodromoussecondaryveinframework(Soares-Silva
andProenc¸a,2008;Gomesetal.,2009;Campos,2010).
Researchconcerningtheaspectsofleafarchitecturecanbeas fundamentalasthemorphologyofthereproductiveorgansused insystematicstudies(HickeyandTaylor,1991).AccordingtoEllis
et al. (2009), thestudy of leaf architecture is a relatively new
approachthatcanprovideimportantinformationontaxonomically complexfamilies,suchasMyrtaceae.
AccordingtoArantes and Monteiro (2002), despitethe high representationofMyrtaceaeindifferentbiomes,fewtaxonomic studieshavebeenconducted,reflectingthediversityofspeciesand taxonomiccomplexityoftheseplants.Themorphologicalstudies ofthisfamily,andespeciallyofthePsidiumgenus,cancontribute toecological and phytosociological studies, providing subsidies tovalidatethetaxonomicclassification and recognitionofnew species(De-Carvalho,2008).
Morphologicalreferencesareextremelyimportant,particularly formedicinalplants,sothatthesamplescanbeconfirmedandto enabletestsofauthenticity.Thesetestsareneeded,because confu-sionbetweenmorphologicallysimilarspeciesisverycommonand canleadtotheimproperuseofaparticularspecies.Thus,theleaf architectureaddstothesetofinformationthatallowstheaccurate identificationofplantspecies.
The aim of the present study was to describe the morphology and foliar venation of four species of Psid-ium collected from a Savanna biome in Goiás State, Brazil, to contribute to the identification and delineation
of these taxa, and to the quality control of the plant material.
Materialandmethods
LeavesofPsidiumspp.werecollectedfromtheBrazilianSavanna areasinGoiásState,in2013and2014,inthecitiesofAnápolisand CaldasNovas(Fig.1;Table1).
Thebotanicalmaterialcomprisedfullyexpandedadultleaves collected below thethird node. For each individual, a voucher specimenwasdepositedintheHerbariumofGoiásState
Univer-sity–HGSU(Table1),andthenextcharacteristicswererecorded
foreachvoucherspecimen:P.firmumO.Berg.:shrub,about90cm high;stemsmooth,cylindrical,glabrous,andbrown;P.myrsinites DC.:tree,about3mhigh;stemsmoothcylindrical,glabrous,with irregular plate depressions that are detached from the trunk, slightly pink browncoloration; P.laruotteanum Cambess.: tree, about3mhigh;stemwithcrackedrhytidomaandsinuousand dis-continuouscrests,barkwithdarkgrayishcoloration;P.guineense Sw.:tree,about5mhigh;smoothcylindricalstem,yellowishgray coloration.
Leafmorphologicalcharacterizationwasperformedby obser-vationswiththenakedeyeusingastereoscopicmicroscope,when necessary,accordingtoOliveiraandAkisue(2003)andEllisetal.
(2009).
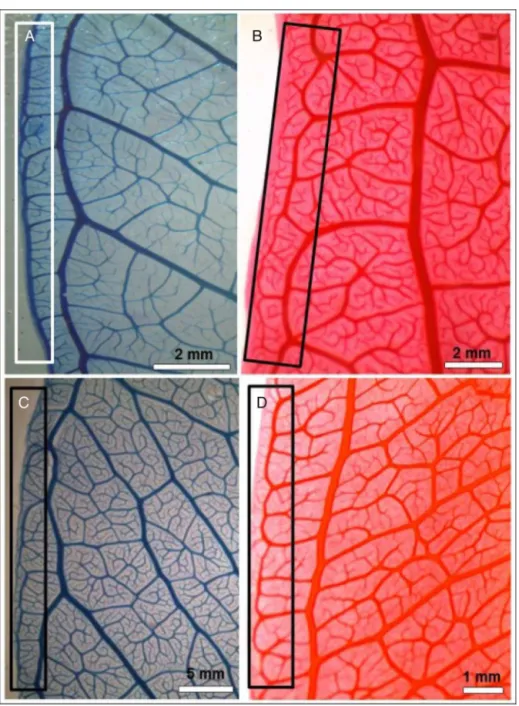
Fig.3.Psidiumspp.leafveincharacters.(A)P.firmum,intersecondaryvein(arrow);(a1)Veinorders(untilthe5thorder)(arrows);(B)P.myrsinites,irregularareoles(square); (C)P.laruotteanum,freelyendingveinlets(FEV)(arrow);(D)Psidiumguineense,irregularareolesandfreelyendingveinlets(FEV)(arrow).Sv,secondaryvein;Tv,tertiary vein;Qn,quaternaryvein;Qun,quinternaryvein;Ve,venule;Mi,midrib;Isv,intersecondaryvein.
hypochlorite.Subsequently,dehydrationwasperformedinaseries of 70% (w/w),90% (w/w) and absoluteethanol, incubating the plantmaterialfor1hin eachsolution.Theleaveswerestained withmethyleneblue/xylol(1:1)orsafranin/xylol(1:1).Eachleaf wasimmersed in xylol and immediatelytransferred to a glass platecontainingresin,mounted ontwoplates and dried. Anal-yses were performed using a LEICA, EZ4D model stereoscopic microscope.
Thepatternvenationregistrationwasgeneratedfrom expan-sionsofdiaphanizedleaves(usingaphotographicamplifierDurst M601),andsmallernetworksoftheveinswererecordedusinga OlympusBX40photomicroscope.Macroandmicroscopic charac-terizationsoftheleafvenationpatternswereperformedaccording
toCardosoandSajo(2004,2006)andEllisetal.(2009).
Resultsanddiscussion
TheleavesofthePsidiumspeciesanalyzedinthepresentstudy weretypicallycoriaceous,withshortpetiolesandaprimary pin-natetypevenationpattern,andcharacterizedbyonlyonemidrib ofahighergauge.
Theleaf laminates have severalshapes, and elliptic or obo-vateshapeswereprimarilyobserved.Theapexshapeswereacute toacuminatedinP.firmum,P.laruotteanum,andP.myrsinites.P. guineensedisplayedapexobtuse tomucronateshapes.Thebase shapeswereacutetocuneateinP.myrsinitesandP.laruotteanum androundedinP.firmumandP.guineense(Box1;Fig.2).
Trichomeswerepresentontheabaxialsurfaceoftheleaflamina ofallspecies.P.guineensedisplayedtrichomesonbothsurfaces (Box1).
Theleafmidribwasmoreprominentontheabaxialsurfacein thefourspecies.P.firmum(Fig.2A)andP.guineense(Fig.2D) pre-sentedgroovedmidribsonadaxialsurface(Box1).
The major secondary vein framework was the
camptodromous–brochidodromous mixed venation pattern, followedbythebrochidodromouspattern(Box1;Fig.2).
ThebrochidodromousvenationpatternwasobservedinP. fir-mum(Fig.2a1),consistentwiththeobservationofCampos(2010)
for this species. This venation pattern is formed when secon-dariesjoininaseriesofprominentarchesandloopsofsecondary gauge(Elliset al.,2009).AccordingtoSoares-Silvaand Proenc¸a
(2008),brochidodromousisthetypicalvenationpatternfor
Psid-ium.CardosoandSajo(2006)reportedthatthebrochidodromous
archesandloopsmightbemoreorlessprominentdependingon theclosingangleofthesecondaryveins.
Box1:LeafmorphologicalcharactersandvenationoffourspeciesofPsidiumspp.
Characters/species P.firmum P.myrsinites P.laruotteanum P.guineense
Shape Elliptic to oblong Elliptic to obovate Elliptic to obovate Elliptic to obovate Leaf Apex Acutetoacuminate Acutetoacuminate Acutetoacuminate Acutetomucronate
Base Rounded Acutetocuneate Acutetocuneate Rounded
Petiole Short-petiolate Short-petiolate Short-petiolate Short-petiolate
Indument Adaxialsurface Glabrous Glabrous Glabrous Pilose
Abaxialsurface Pilose Pilose Pilose Pilose
Mainveinsalience Adaxialsurface Grooved Prominent Prominent Grooved
Abaxial surface Prominent Prominent Prominent Prominent
Primaryvenationpattern Pinnate Pinnate Pinnate Pinnate
Secondaryvenation Venationpattern Brochidodromous Camptodromous– brochidodromous
Camptodromous– brochidodromous
Camptodromous– brochidodromous
Numberofpairs <13 <13 <13 <10
Salienceadaxialsurface Flat Prominent Flat Grooved
Salienceabaxialsurface Flat Flat Flat Prominent
Vein (orders) Until the 5th order Until the 5th order Until the 5th order Until the 5th order
Intersecondaryveins Present Present Present Present
Intramarginalveins Absent Absent Absent Absent
Marginalultimatevenation Fimbrial Incompleteloops Looped Looped
Collectorvein Distancefromtheedge (mm)
<1 1–3 1–3 1–3
Venules Simple/branching Branching Branching Branching Branching
Areoles Present Present Present Present
The camptodromous–brochidodromous secondary venation pattern is common for representatives of Myrtinae, Myrciinae andEugeniinae(Costaetal.,1995;FarmacopeiaBrasileira,2002;
Cardoso and Sajo, 2004, 2006). Costa et al. (1995) described
the camptodromous–brochidodromous standard for six of the elevenEugeniaspeciesoccurringinthesandbankofMarica,Rio de Janeiro State in Brazil. In the Brazilian Pharmacopoeia IV, this secondary venation pattern is registered to P. guajava L.
(Farmacopeia Brasileira,2002).Cardoso and Sajo(2004)
identi-fied the camptodromous–brochidodromoussecondary venation standardinnine(9)Eugeniaspeciesfromthehydrographicbasinof TibagiRiver,ParanáState,Brazil:EugeniaarenosaMattos,E. hamil-tonii(Mattos)Mattos,E.handroanaD.Legrand,E.hiemalisCambess., E.neoverrucosaSobral,E.pyriformisCambess.,E.ramboiD.Legrand, E.speciosaCambess.,andE.unifloraL.
The arrangement of secondary veins relative to the midrib in the leaves of the studied species produces angles with divergences ranging from 45◦ to 75◦. According to Cardoso
and Sajo (2004), the divergence angles ranging from 45◦ to
60◦ are most striking in species withmixed venation patterns (camptodromous–brochidodromous),asrecordedinthepresent studyforP.laruotteanum,P.myrsinites,andP.guineense.The diver-genceanglescannotbeconsideredtobetaxonomiccharactersfor distinguishingspecies,asinthepresentstudy,allspeciesshowed widevariationsamongtheanglesinleavesfromthesame individ-ual.
Theamountofsecondaryveinpairsforeachspeciesisshownin
Box1.P.guineense(Fig.2d1)presentsfewersecondaryveinpairs thanotherspecies(8–10pairs),therebydistinguishingthisspecies. Allspecieshaveveinsuntilthe5thorder(Fig.3a1).
Notethepresenceofintersecondaryveinsinallspecies(Box1;
Fig.3A).AccordingtoEllisetal.(2009),theintersecondaryveins originatefromthemidrib,runningparalleltothemajorsecondary veins,ascharacterizedbyanintermediategaugebetweenthe sec-ondaryandtertiaryveins.Inthepresentstudy,nospeciesshowed intramarginalveins,whichaccordingtoCardosoandSajo(2004), aretheveinsformedbetweenbrochidodromousarchesandtheleaf margin.
Spacing among the secondary veins occurred in decreasing ordertowardtheapex.IrregularspacingwasobservedinP.firmum, P.myrsinites,andP.guineense(Fig.2a1,b1,d1),andregularspacing wasobservedinP.laruotteanum(Fig.2c1).
Theconfigurationofthehighest-orderleafveinsinthe mar-ginormarginalultimatevenationwasabsent,showingincomplete loop,looped,andfimbrialtypes.Intheincompletelooptype,the marginal ultimateveinrecurvestoformincomplete loopsnear theleafmargin;intheloopedtype,thetertiaryveinsanastomose, formingaseriesofarchesneartheedge;thefimbrial veintype occurswhencloseveinsjoinattheedgeandproduceacontinuous veinrunningalongthemargin(CardosoandSajo,2006;Ellisetal.,
2009).
Inthepresentstudy,themarginalultimatevenationofthe fim-brialtypewasobservedonlyinP.firmum(Box1;Fig.4A).InP. myrsinites(Box1;Fig.4B),theincompletelooptypewasobserved, andinP.laruotteanum(Fig.4C)andP.guineense(Fig.4D)thelooped types,whichhaveasmallergaugeclosertothemargin,were ver-ified.AccordingtoGomesetal.(2009)andSilvaetal.(2008),in mostsurveysofPsidium,themarginalultimatevenationpatternis theloopedtype,formedbycompletearches,consistentwiththe resultsobtainedinthepresentstudy.
Themarginal ultimateloopedtype venation patternis com-moninmostspeciesoftheMyrtaceaefamily,followedbyfimbrial andincompletelooptypes(CardosoandSajo,2006;Oliveiraetal.,
2011).De-Carvalho(2008)registeredthefimbrialandloopedtypes
intwoandsevenspecies,respectively,oftheninespeciesofMyrcia DC.(Myrtaceae)studiedinFederalDistrict,Brazil.
Areolesarethesmallestareasoftheleaftissuecompletely sur-roundedbyveinsofthefourthandfifthorders.Thesestructurescan havevariousshapesandarrangements,withorwithoutvenules (simpleorramified).Theareolesareconsideredtoberegular (per-fect)whentheyhavethesameshapeandsize;irregular(imperfect) areolesvaryinsizeand shape;andincompleteareoleshaveno limitationsonthesides(Alvarezetal.,2006;Ellisetal.,2009).
Theresultsofthepresentstudyrevealedirregularareoleswith freelyendingveinlets(FEVs)ofthedendriticramifiedtypeforall Psidiumspecies(Box1;Fig.3B–D).Costaetal.(1995)andKlucking
(1988)highlightedthattheformationofincompleteandirregularly
shapedrandomlydistributedareolesisacommoncharacteristicin theMyrtaceaefamily.
Fig.4. Psidiumspp.marginalultimatevenation.(A)P.firmum,marginalultimatevenationoffimbrialtype(rectangle);(B)P.myrsinites,marginalultimatevenationof incompletelooptype(rectangle);(C)P.laruotteanum,marginalultimatevenationofloopedtype(rectangle);(D)P.guineense,marginalultimatevenationofloopedtype (rectangle).
Box2:MorphologicalidentificationkeyforPsidiumspp.
1 Shrubbyhabit;brochidodromoussecondaryvenationpattern;marginalultimatevenationfimbrialtype P.firmum
1 Tree habit; camptodromous–brochidodromous secondary venation pattern; marginal ultimate venation looped type 2
2 Ridged adaxial surface with prominent midrib on the abaxial surface; pilose on both surfaces; apex obtuse to mucronate; base rounded P.guineense
2 Adaxialsurfacewithprominentmidrib;glabrousadaxialsurfaceandpiloseabaxialsurface;apexacutetoacuminate;baseacutetocuneate 3
3 Secondaryveinsprominentontheadaxialsurfaceandmarginalultimatevenationwithincompleteloops P.myrsinites
3 Secondaryveinsflatontheadaxialsurfaceandmarginalultimatevenationlooped P.laruotteanum
veinshavedifferentspacingpatterns,withacutedivergenceangles tothemidrib,aplaneontheadaxial surfaceand areprimarily prominentontheabaxialsurface.Allplantshaveintersecondary veins.Themarginalultimatevenationoftheloopedtypeappearsin mostspeciesandmightappearincompleteandfimbrial.The areo-lasarewelldevelopedveinsofthefourthandfifthorders,andhave branchedFEVs.
Thedifferentcharactersobservedgeneratedthefollowing iden-tificationkeyforPsidiumspecies(Box2).
Psidiumspecies,furtherconfirmingthatdiaphanizationisan inex-pensive,fastandconclusivetechnique.
Notably,phenotypic variation reflecting theinfluence ofthe environmentwasnotconsideredinthisanalysis,asthesiteswhere thespecieswerecollectedhavesimilarmicroclimates.
Authors’contributions
EFO (Master student) contributed incollecting plantsample andidentification,confectionofherbarium,runningthelaboratory work,analysisofthedataanddraftedthepaper.DGBcontributed incollectingplantsample,confectionofherbarium,runningthe laboratorywork,analysisofthedataanddraftedthepaper.MLS contributedincollectingplantsampleandidentification, confec-tionofherbarium,analysisofthedataandtocriticalreadingofthe manuscript.MHRcontributedtocriticalreadingofthemanuscript. JAMPdesignedthestudy,supervisedthelaboratoryworkand con-tributedtocriticalreadingofthemanuscript.Alltheauthorshave readthefinalmanuscriptandapprovedthesubmission.
Conflictsofinterest
Theauthorsdeclarenoconflictsofinterest.
Acknowledgments
TheauthorswouldliketothankJairEustáquioFariaJúniorfor assistance withmaterial identification, Francisco JuniorSimões Calac¸afordesigningtheboards,CAPESandFundac¸ãodeAmparo àPesquisadoEstadodeGoiásforfinancialsupport.
References
Alvarez,A.S.,Potiguara,R.C.V.,Santos,J.U.M.,2006.Arquiteturafoliardeespéciesde EugeniaL.(Myrtaceae)darestingadeAlgodoal,Maiandeua,PA.Bol.Mus.Para. EmilioGoeldi2,29–36.
Arantes,A.A.,(Masterdissertation)1997.FlorísticadafamíliaMyrtaceaeJuss.na Estac¸ãoEcológicadoPanga,Uberlândia,MG.Unesp,RioClaro,SP.
Arantes,A.A.,Monteiro,R.,2002.AFamíliaMyrtaceaenaEstac¸ãoEcológicadoPanga, Uberlândia,MinasGerais,Brasil.Lundiana3,111–127.
Barroso,G.M.,Peixoto,A.L.,Ichaso,C.L.F.,Costa,C.G.,Guimarães,E.F.,Lima,H.C., 1991.SistemáticadeangiospermasdoBrasil.UniversidadeFederaldeVic¸osa, ImprensaUniversitária,Vic¸osa.
Berg,O.,1859.Myrtaceae.In:Martius,C.P.F.(Ed.),Florabrasiliensis14.,pp.529–656. Brandão,M.,Laca-Buendía,J.P.,Macedo,J.F.,2002.Árvoresnativaseexóticasdo
EstadodeMinasGerais.EPAMIG,BeloHorizonte.
Campos,L.Z.O.,(Masterdissertation)2010. EtnobotânicadogêneroPsidiumL. (Myrtaceae)noCerradobrasileiro.Brasilia.InstitutodeCiênciasBiológicas, Uni-versidadedeBrasília,Brasília,71pp.
Cardoso,C.M.V.,Sajo,M.G.,2004.Vascularizac¸ãofoliareaidentificac¸ãodeespécies deEugeniaL.(Myrtaceae)dabaciahidrográficadoRioTibagi,PR.Rev.Bras.Bot. 27,47–54.
Cardoso,C.M.V.,Sajo,M.G.,2006.Nervac¸ãofoliaremespéciesbrasileirasde Myr-taceaeAdans.ActaBot.Bras.20,657–669.
Corrêa,L.C.,Santos,C.A.F.,Vianello,F.,Lima,G.P.P.,2011.Antioxidantcontentin guava(Psidiumguajava)andarac¸á(Psidiumspp.)germplasmfromdifferent Brazilianregions.PlantGenet.Resour.9,384–391.
Costa,I.R.,(Masterdissertation)2004.Estudoscromossômicosemespéciesde Myr-taceaeJuss.nosudestedoBrasil.InstitutodeBiologia,UniversidadeEstadualde Campinas,Campinas.
Costa,I.R.,(Ph.D.thesis)2009.EstudosevolutivosemMyrtaceae:aspectos cito-taxonômicos e filogenéticos em Myrteae, enfatizando Psidium e gêneros
relacionados.InstitutodeBiologia,UniversidadeEstadualdeCampinas, Cam-pinas,235pp.
Costa,C.G.,Machado,R.D.,Fontenelle,J.B.,1995.Sistemavascularemfolhasde EugeniaL.(Myrtaceae).Bol.Herb.Bradea.6,345–356.
De-Carvalho,P.S.,(Masterdissertation)2008.MyrciaDC.exGuill.(sec¸ãoMyrcia, Myrtaceae)noDistritoFederal,Brasil.UniversidadedeBrasília,Brasilia,68pp. Ellis,B.,Daly,D.C.,Hickey,L.J.,Johnson,K.R.,Mitchell,J.D.,Wilf,P.,Wing,S.L.,2009.
ManualofLeafArchitecture.CornellUniversityPress,Ithaca.
FarmacopeiaBrasileira,2002.MonografiaGoiabeira,4thed.parte2.fasc.4,Atheneu, SãoPaulo.
Fauth,S.,Campos,A.R.,Silveira,E.R.,Rao,V.S.,2002.Efeitosdeóleosessenciaisde plantasnotempodesonoinduzidoporcetaminaemcamundongos.Rev.Bras. Farmacogn.12,112–113.
Frazon,R.C.,Campos,L.Z.O.,Proenc¸a,C.E.B.,Sousa-Silva,J.C.,2009.Arac¸ásdogênero Psidium:principaisespécies,ocorrência,descric¸ãoeusos.EmbrapaCerrados, Brasília(SérieDocumentos,Embrapa).
Gomes,S.M.A.,Somavilla,N.S.D.N.,Gomes-Bezerra,K.M.,Miranda,S.C.,De-Carvalho, P.S.,GracianoRibeiro,D.,2009.AnatomiafoliardeespéciesdeMyrtaceae: contribuic¸õesàtaxonomiaefilogenia.ActaBot.Bras.23,223–238.
Hickey,L.J.,Taylor,D.W.,1991.TheleafarchitectureofTicodendronandthe appli-cationoffoliarcharactersindiscerningitsrelationships.Ann.Mo.Bot.Gard.78, 105–130.
Klucking,E.P.,1988.LeafVenationPatterns.Myrtaceae.J.Cramer,Stuttgart. Kraus,J.E.,Arduin,M.,1997.Manualbásicodemétodosemmorfologiavegetal.EDUR,
Seropédica.
Landrum,L.R.,Kawasaki,M.L.,1997.ThegeneraofMyrtaceaeinBrasil:anillustrated synoptictreatmentandidentificationkeys.Brittonia49,508–536.
McVaugh,R.,1968.ThegeneraofAmericanMyrtaceae:aninterimreport.Taxon 17,354–418.
Medina,A.L.,Haas,L.I.R.,Chaves,F.C.,Salvador,M.,Zambiazi,R.C.,Silva,W.P.,Nora, L.,Rombaldi,C.V.,2011.Arac¸á(PsidiumcattleianumSabine)fruitextractswith antioxidantandantimicrobialactivitiesandantiproliferativeeffectonhuman cancercells.FoodChem.128,916–922.
Mendonc¸a,R.C.,Filgueiras,T.S.,Fagg,C.W.,2007.AnáliseFlorísticadaChapadados Veadeiros.In:Felfili,J.M.,Rezende,A.V.,Silva,J.R.(Eds.),BiogeografiadoBioma Cerrado:Vegetac¸ãoeSolosdaChapadadosVeadeiros.EditoraUniversidadede Brasília/Finatec,Brasília,pp.119–237.
Mittermeier,R.A.,Robles,P.,Hoffmann,M.,Pilgrim,J.,Brooks,T.,Mittermeier,C.G., Lamoreux,J.,Fonseca,G.B.,2005.HotspotsRevisited:Earth’sBiologicallyRichest andMostEndangeredEcoregions.ConservationInternational,MexicoCity. Myers,N.,Mittermeier,R.A.,Mittermeier,C.G.,DaFonseca,G.A.B.,Kent,J.,2000.
Biodiversityhotspotsforconservationpriorities.Nature403,853–858. Oliveira,F.,Akisue,G.,2003.Fundamentosdefarmacobotânica,2nded.Atheneu,
SãoPaulo.
Oliveira-Filho,A.T.,Fontes,M.A.L.,2000.Patternsoffloristicdifferentiationamong AtlanticForestsinSoutheasternBrazilandtheinfluenceofclimate.Biotropica 32,793–810.
Oliveira,M.I.U.,Funch,L.S.,Santos,F.A.R.,Landrum,L.R.,2011.Aplicac¸ãode carac-teresmorfoanatômicosfoliaresnataxonomiadeCampomanesiaRuiz&Pavón (Myrtaceae).ActaBot.Bras.25,455–465.
Oliveira,V.B.,Yamada,L.T.,Fagg,C.W.,Brandão,M.G.L.,2012.Nativefoodsfrom Brazilian diversityasasource ofbioactivecompounds.Food Res.Int. 48, 170–179.
Patel,S.,2012.ExotictropicalplantPsidiumcattleianum:areviewonprospectsand threats.Rev.Environ.Sci.Biotechnol.11,243–248.
Silva,F.A.M.,Assad,E.D.,Evangelista,B.A.,2008.Caracterizac¸ãoclimáticadoBioma Cerrado.In:Sano,S.M.,Almeida,S.P.,Ribeiro,J.F.(Eds.),Cerrado:EcologiaeFlora. Embrapa,Planaltina,pp.69–88.
Silva,J.M.C.,Bates,J.M.,2002.BiogeographicpatternsandconservationintheSouth AmericanCerrado:atropicalsavannahotspot.BioScience52,225–233. Soares-Silva,L.H.,Proenc¸a,C.E.B.,2008.AnewspeciesofPsidiumL.(Myrtaceae)from
southernBrazil.Bot.J.Linn.Soc.158,51–54.
Sobral, M.,Proenc¸a, C.,Souza, M., Mazine,F., Lucas,E., 2013. Myrtaceae. In: Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro, http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB171/(accessedDecember 2013).
Souza,V.C.,Lorenzi,H.,2008.Botânicasistemática:guiailustradoparaidentificac¸ão dasfamíliasdeAngiospermasdaflorabrasileira,baseadoemAPGII.Instituto Plantarum,NovaOdessa.