FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
VERTICAL DYNAMICS OF PLANKTONIC
COMMUNITIES AT SOFALA BANK, MOZAMBIQUE
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
VERTICAL DYNAMICS OF PLANKTONIC
COMMUNITIES AT SOFALA BANK, MOZAMBIQUE
Dissertação orientada pelos:
Professor Doutor José Paula
Professora Doutora Vanda Brotas
Miguel Albuquerque da Costa Leal
MESTRADO EM ECOLOGIA MARINHA
Acknowledgements
To all people that in some way contributed to this work, I express my sincere gratitude, especially to:
Professor Doutor José Paula, for taking a chance on me and for all the support, supervision, confidence, friendship and transfer of knowledge, but most of all for his joviality and all the joyfull moments, which turned this hard and serious work into a delightful and pleasant learning process. Professora Doutora Vanda Brotas, for having accepted to guide this thesis and for the support, recommendations and companionship, for believing in me and also for spending some of her precious (and little) time with me.
Carolina Sá and Rafael Mendes, without whom part of this work would never be possible, for all the trust, patience, and time spent on laboratory, as well for the friendship, support and enthusiasm during this work, even in the hardest moments.
All people onboard the R/V “Dr. Fridtjof” Nansen, for the collaboration during sample period, namely Jens‐Otto Krakstad and Diana Zaera, for the nice discussions and help with paperwork, and also to Carlos Bento, Domingos Zacarias e Zé Chamuse for their very important help on the sampling process.
Sónia Nordez and Maurício Lipassula, for their help processing plankton samples in Lisbon; to James Mwaluma, for the larval fish identification; and to Paulo Oliveira for his expertise on current data interpretation.
All the people of the LMG, namely Ricardo Mendes, Gil Penha‐Lopes and Laura Antão, for the friendship and nice chats when difficult choices had to be made.
Bruno Jesus, for his patience in teaching me the world of R and statistics, which definitively was very important for the statistical and graphical analysis.
All my friends, both from the University and Ponto Zero, for all the joy and happiness and also for encouraging and cheering me up in my worst days. Special thanks to my truly friend Vasco Mota, for
ii
all the articles he sent me from the Netherlands and, most of all, for his friendship and nice discussions.
My Family, for the unconditional support, concern, care, recommendations and patience. Special thanks to my mother, father and sister for accepting those (long) periods in which I’m not present, for teaching me the importance of organization at work, discipline and timings, but essentially for what I am today; to my uncle João Martins for the nice chats, holidays and gym classes that were essential to my welfare and mental sanity; to my uncle João Caraça, for the conversations that gave me a scientific view of this world; and also to my uncle Luis Leal, for my initiation to biology and kindle my interest in become a marine biologist, and to the discussions and stories about the natural world, which are the reason for my interest in biology and curiosity to explore and understand our natural world.
At last, but not, by any means, less important, to Marta, my source of motivation to be a better person and a hard working scientist, for all the love, care, patience and trust, for providing me everyday with a smile in my face and for making me understand how much this life is worth living. Thank you for being here!
Resumo
As zonas costeiras são ecossistemas de grande importância para as actividades humanas, onde se desenrolam complexos processos naturais com elevada variabilidade espacial e temporal. Destes processos, com particular importância para a geração de recursos, destacam‐se os fenómenos de maré e as correntes costeiras, que, entre outros factores, estão fortemente associados às variações das propriedades físico‐químicas da massa de água costeira (p.e. temperatura, salinidade e nutrientes). Consequentemente, esta variabilidade influencia as diferentes comunidades que estão na base dos ecossistemas marinhos, como as comunidades fito‐ e zooplanctónicas, assim como as larvas de peixes e de invertebrados marinhos. É de salientar que, no que respeita aos processos bio‐ ecológicos das larvas de peixes e de invertebrados marinhos, a variação na abundância e padrões de distribuição na zona costeira está fortemente dependente de fenómenos como os ventos e correntes que, entre outros factores, condicionam a sobrevivência e, consequentemente, a taxa de recrutamento para as populações de organismos costeiros.
O presente estudo decorreu no Banco de Sofala (Moçambique), uma extensa plataforma continental existente na zona costeira situada em frente ao delta do Rio Zambeze (um dos maiores rios de África), onde ocorrem as mais importantes actividades pesqueiras de Moçambique. Tendo em consideração a importância económica do Banco de Sofala e a grande influência do Rio Zambeze no funcionamento dos processes ecológicos costeiros, os objectivos desta investigação foram direccionados para as variações existentes ao nível hidrológico e biológico que suportam esta intensa actividade pesqueira. Aspectos como as propriedades físico‐químicas da massa de água e correntes costeiras foram estudados conjuntamente com a distribuição e abundância de nutrientes, comunidades fitoplanctónicas (estudados através de assinaturas de pigmentos analisadas por HPLC), zooplâncton e larvas de peixe (especificamente da espécie Herklotsichthys quadrimaculatus), decorrentes no Banco de Sofala. Os trabalhos desenvolvidos foram especialmente direccionados para a distribuição horizontal na plataforma continental (baseada na amostragem de 11 estações radiais distribuídas ao longo de 3 transectos perpendiculares à costa), e para a dinâmica vertical destes processos ao longo de um ciclo de 48 horas (com uma periodicidade de amostragem de 2 h numa estação fixa). O processo de amostragem decorreu ao longo de 3 dias e foi desenvolvido no decorrer de uma missão oceanográfica realizada a bordo do N/O “Dr. Fridtjof Nansen”, integrada num projecto direccionado para o estudo dos ecossistemas costeiros de Moçambique.
iv
A variação horizontal da maioria dos parâmetros analisados revelou a existência de gradientes costa – largo e de descargas do rio Zambeze, evidenciadas pela de intrusão de águas estuarinas na zona costeira. A distribuição superficial das propriedades físico‐químicas analisadas evidenciou a presença de uma massa de água menos salina e mais quente na zona central, associada provavelmente a descargas estuarinas, enquanto que a distribuição das concentrações de nutrientes só evidenciou gradientes perpendiculares à costa para os silicatos, dado que a distribuição de fosfatos foi independente da distância à costa e a concentração de nitratos foi sempre menor que o limite de detecção do método. Relativamente à distribuição superficial de fitoplâncton, foi observada maior biomassa numa das estações mais costeira, visto que foi provavelmente a primeira a ser influenciada pelas descargas do rio Zambeze e, consequentemente, onde maior concentração de nutrientes ficou primeiramente disponível para o fitoplâncton. O único grupo de fitoplâncton com diferente distribuição relativamente à biomassa fitoplanctónica foi o das Cianobactérias, para o qual foram observadas maiores concentrações pigmentares na massa de água mais oceânica.
No que respeita a dinâmica vertical estudada ao longo de 48 horas, as correntes de maré revelaram ser um factor crucial para as variações hidrológicas decorrentes na coluna de água, nomeadamente para o transporte horizontal e vertical da biomassa fitoplanctónica e para a variação da disponibilidade de nutrientes. As concentrações de nutrientes obtidas estão de acordo com resultados obtidos em outras zonas costeiras oligotróficas, contudo os rácios de nutrientes obtidos (N:P e N:Si) foram extremamente baixos e fortemente influenciados pelas reduzidas concentrações de nitratos + nitritos (N). As concentrações de N obtidas foram anormalmente baixas relativamente a muitos ecossistemas costeiros, contudo semelhantes a alguns estudos obtidos no Oceano Índico, nomeadamente na Austrália e no Quénia, onde o N desempenha um papel crucial como factor limitante da biomassa fitoplanctónica. Para além destas concentrações evidenciarem o estado oligrotrófico do Banco de Sofala durante o período de amostragem, sugerem ainda a existência de reduzidas descargas estuarinas para a zona costeira, o que é típico da época seca durante a qual foi realizado este estudo.
As maiores concentrações de pigmentos fitoplanctónicos foram observadas nas maiores profundidades amostradas (30 e 40 m), onde também foram registadas maiores concentrações de N. Durante o decorrer do estudo, a comunidade fitoplanctónica foi dominada por microflagelados, mais concretamente por microalgas do grupo Prymnesiophyceae, o que está de acordo com outros estudos em regiões oligotróficas onde a dominância de microflagelados, e não de microplâncton (diatomáceas e dinoflagelados), é habitualmente observada. De acordo com os resultados obtidos, toda a comunidade fitoplânctónica distribuiu‐se de modo similar na coluna de água ao longo do
espaço de tempo considerado, com a excepção das Cianobactérias que apresentaram uma distribuição vertical mais superficial.
Relativamente à distribuição do zooplâncton, foram observadas elevadas densidades junto ao fundo (30/40 m), onde as maiores concentrações de fitoplâncton foram também verificadas. Durante o período nocturno foram registadas maiores densidades de zooplâncton nas camadas de água mais superficiais (< 10 m) relativamente aos períodos nocturnos, o que evidencia os processos de migração vertical típicos das comunidades zooplanctónicas onde a variação da intensidade luminosa desempenha um papel crucial. Contudo, o movimento vertical observado envolveu apenas uma pequena parte da abundância total de zooplâncton, visto que as maiores densidades observadas à superfície durante a noite foram geralmente inferiores às densidades observadas no estratos mais profundos (20/30 e 30/40 m).
Relativamente à investigação direccionada especificamente para as larvas de Herklotsichthys
quadrimaculatus, os resultados obtidos evidenciaram o padrão típico de migração vertical com
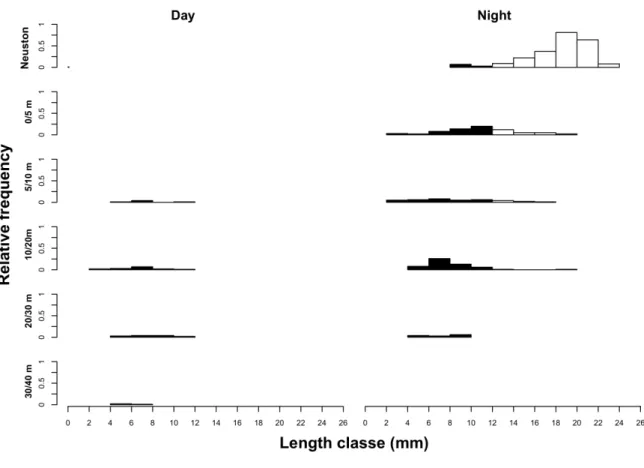
periodicidade diária, realizando movimentos para a superfície e para o fundo durante o anoitecer e amanhecer, respectivamente. Relativamente à variação desta dinâmica vertical em função das variações ontogenéticas, os resultados obtidos evidenciam diferenças na posição vertical das larvas em função do período do dia e do estádio de desenvolvimento. As larvas em estádios de desenvolvimento mais avançados dominaram a massa de água superficial da coluna de água (neuston) durante a noite, enquanto que as larvas mais pequenas estiveram distribuídas na camada de água subjacente (0 – 20 m). Durante o dia, a quantidade de larvas presente nos estratos amostrados foi quase nula, o que evidencia a agregação das larvas de H. quadrimaculatus junto ao fundo (40 – 50 m), onde a penetração luminosa é muito reduzida e, consequentemente, o risco de mortalidade associado a predadores visuais é baixo.
A investigação realizada realça a importância dos processos físico‐químicos nas zonas costeiras e o seu papel na dinâmica vertical das comunidades planctónicas. Este caso de estudo exemplifica os processos base que ocorrem num ecossistema costeiro tropical onde as marés e as descargas estuarinas são factores importantes na dinâmica das comunidades biológicas presentes no plâncton. Os resultados obtidos são então um exemplo dos ecossistemas costeiros do Oceano Índico Ocidental, onde a dinâmica das comunidades planctónicas é ainda pouco conhecida e compreendida.
Palavras‐chave: Pigmentos fitoplanctónicos; Zooplâncton; Estádios larvares; Herklostichthys
Abstract
Coastal ecosystems are largely influenced by the interaction of several factors operating at various temporal and spatial scales, specifically those responsible for primary and secondary production processes that modulate marine resources. Hydrological processes (e.g. tides and coastal currents), nutrients availability, phytoplankton groups, zooplankton and larval fish (specifically the clupeid Herklotsichths quadrimaculatus) abundance and distribution were investigated at the Sofala Bank (Mozambique), with special emphasis on their horizontal distribution and vertical dynamics. Horizontal distribution has shown onshore‐offshore gradients in all analysed parameters, as well as inshore waters intrusion probably related to Zambezi River delta runoff. Tidal currents were responsible for major hydrological vertical variations and for horizontal and vertical advection of phytoplankton biomass in the surface and deepest layers, respectively. Nutrient concentrations were typical from oligotrophic regions, and nutrient ratios were strongly influenced by depleted nitrite + nitrate concentrations, suggesting low estuarine discharges typical from the dry season. Both phytoplankton pigments and zooplankton were found mainly near bottom (40 m deep), despite the latter displayed vertical migrations triggered by light variations. Phytoplankton community was dominated by microflagellates, specifically prymnesiophyceans, and vertical distribution changes were similar for the whole community. Cyanobacteria was the only phytoplankton group that displayed a different vertical distribution pattern, mainly concentrated at mid water column depths (10 – 20 m). Herklotsichthys quadrimaculatus larvae displayed typical diel vertical migrations and were mainly distributed in the upper water column (0 – 20 m) during the night and almost absent from all sampled strata during the day. Larger larvae dominated the neuston layer during the night while during daylight periods remained close to the bottom. This investigation enhances the importance of physico‐chemical phenomena determining the planktonic communities vertical dynamics at a tropical coastal ecosystem of the Western Indian Ocean, where planktonic dynamics are still poorly described and understood.
Keywords: Phytoplankton pigments; Zooplankton; Fish larvae; Herklostichthys quadrimaculatus;
Diel vertical migration; Mozambique
Acknowledgements i
Resumo iii
Abstract vii
CHAPTER 1
1
General introduction 3References 8
CHAPTER 2
11
Distribution and vertical dynamics of planktonic communities at Sofala Bank, Mozambique 13Abstract 13
Introduction 14
Material and Methods 16
Results 18
Discussion 30
Conclusion 34
References 36
CHAPTER 3
41
Vertical dynamics of the gold‐spot herring (Herklotsichthys quadrimaculatus) larvae at Sofala Bank,
Mozambique 43
Abstract 43
Introduction 43
Material and Methods 45
Results 46
Discussion 50
References 52
General introduction
Coastal ecosystems and planktonic communities
Coastal ecosystems are among the most biologically productive environments in the world, providing important services to human populations. Their existence at the interface between the terrestrial and marine environment exposes them to a wide variety of human and natural stressors occurring at different spatial and time scales, which makes the coastal regions complex and dynamic ecosystems (Kennedy et al., 2002).
The natural stressors occurring in the coastal marine environments are typically physical and chemical processes that differently affect biological communities, hence their population dynamics. Some of the most important physical processes are winds, tides and coastal currents, which promote water mass advection, thus spatial and temporal variability on water column physico‐chemical properties (Epifanio and Garvine, 2001; Lutjeharms, 2006; Queiroga et al., 2007). The interactions between physical and chemical variations induce high heterogeneity on coastal shelf waters, specifically near river deltas, where surface and river runoff together with tidal currents are responsible for strong hydrological gradients and cyclic variations (e.g. temperature and salinity), clines vertical oscillation, both water column stratification and vertical mixing processes, and also large nutrient inputs (Verity et al., 1993; Epifanio and Garvine, 2001; Wawrik et al., 2004; Silva et al., 2008).
Nutrient loadings into coastal ocean zones are from various sources and frequently associated to human activities. Surface and river runoff, estuarine discharges and exchanges with the open ocean are commonly the source and sink of major nutrients inputs, but continental shelf sediments and respiration associated with the sediment surface photosynthesis are also thought to be important nutrient sources and recycling (Wawrik et al., 2004; Wassmann and Olli, 2005, Giraud et al., 2008). Additionally, potentially important sources of nutrients are atmospheric deposition and groundwater discharges. All nutrient inputs to coastal zones, together with the diverse physical processes, differently disturb coastal biological communities, and the most directly influenced are possibly from the lower trophic levels (e.g. phyto‐ and zooplankton) (Cottingham, 1999; Tyrrell, 1999; Anderson et al., 2002; Kennedy et al., 2002).
Chapter 1
4
thriving in coastal systems, strongly determining their abundance and distribution variations (Paula et al., 1998; Cottingham, 1999; Calbet et al., 2001; Elliott et al., 2007). The small scale changes on planktonic communities distribution patterns makes them particularly sensitive to hydrological variations, presenting a great variety of both spatial and temporal changes, respectively from thin water column layers to large‐scale gradients (e.g. onshore‐offshore, north‐south) and from hours to seasons (Verity et al., 1993; Paula et al., 1998; Gibson, 2001; Walther et al., 2002; Balkis, 2003; Brunet and Lizon, 2003; Queiroga et al., 2007; Criales‐Hernández, 2008). Accordingly, these variations are often related to environmental drivers, particularly the preference or avoidance to particular physico‐chemical properties and hydrological processes, such as temperature, salinity, oxygen, light penetration, turbulence, currents and nutrients availability.
Spatial and temporal variations of several planktonic organisms (e.g. phytoplankton, zooplankton and invertebrate and fish larvae), specifically their vertical distribution, have been widely studied in different coastal ecosystems, due to their importance to organic matter flows, primary and secondary production, larval recruitment processes and population connectivity, among others (e.g. Steinberg et al., 2002; Brunet and Lizon, 2003; Fiksen et al., 2007; Leis, 2007; Queiroga et al., 2007).
Plankton vertical dynamics
Vertical migration is a commonly observed phenomenon in many species of freshwater and marine plankton and occurs at a variety of periods, including diel, semi‐diel and also tidal periods (Ohman, 1990; Hill, 1991; Epifanio and Garvine, 2001; Brunet and Lizon, 2003; Fiksen et al., 2007; Queiroga et al., 2007). The explanations for vertical migration may include predation avoidance, bioenergetic advantages and regulation of horizontal position. The hypothesis of a predator avoidance mechanism is generally accepted, as vertical migration often results when prey populations are under intense, selective pressures from visually dependent predators (e.g. Zaret and Suffern, 1976; Irigoien et al., 2004). However, some contradictory cases show that predation is not the only adaptive reason for vertical migration, and the attempt to provide only one hypothesis to account for all observed vertical migrations represent a simplistic outlook on natural selection. Other hypothesis is vertical migration as a bioenergetic advantage, resulting of a compromise between reducing energy expenditure and mortality, taking advantage of feeding opportunities (e.g. Ohman, 1990; Fiksen et al., 2007). The other common hypothesis to planktonic vertical migration is the horizontal position regulation. These communities have very limited capacity for horizontal movements, thus position control by vertical migration potentially provides a means to affect transport to specific locations or to promote retention in particular areas (Hill, 1991; Leis, 2007; Queiroga et al., 2007). The integration of several hydrological and biological processes, such as tidal
currents and behaviour, respectively, regulate dispersal, supply to coastal habitats and recruitment of several planktonic organisms, specifically larval stages of fish and crustacean species (Hill, 1991; Epifanio and Garvine, 2001; Queiroga et al., 2006; Santos et al., 2006). Taking into account the several theories and the diversity of organisms displaying vertical movements there should be variability among vertical migrations patterns, since the same vertical behaviour does not produce the same optimum compromise between reduced mortality and energetic cost for different groups, species and stages (Ohman, 1990; Epifanio and Garvine, 2001; Irigoien et al., 2004; Leis, 2007). Furthermore, the mortality risks for each organism are not the same because of the different selectivity of predators.
Different vertical distribution and diel vertical migration (DVM) can be found among plankton communities, from phyto‐ to zooplankton and also invertebrate and fish larvae. Phytoplankton vertical distributions may differ between day and night, indicating either migration or differences in production/mortality rates (Hajdu et al., 2007; Brunet et al., 2008). Accordingly, the vertical distribution differences found on previous studies are mainly triggered by light requirements and nutrient availability in different water column layers. However, sinking cells and water exchange ascribable to tidal currents and vertical mixing processes can easily bias results, since phytoplankton cells are easily advected (Brunet and Lizon, 2003; Hajdu et al., 2007). Furthermore, additional difficulties arise when taking into account phytoplankton − zooplankton interactions, specifically grazing and nutrient regeneration, which affect phytoplankton abundance estimations (Vertiy et al., 1993; Vrede et al., 1999). Zooplankton itself also displays vertical movements, both to predate and avoid predators (Zaret and Suffern, 1976; Ohman, 1990). Accordingly, both normal and reverse DVM are found, strongly determined respectively by diurnal or nocturnal predators, and also by prey distribution. Besides phyto‐ and zooplankton, invertebrate and fish species commonly have planktonic larval stages, which also display DVM (Epifanio and Garvine, 2001; Hare and Govoni, 2005; Fiksen et al., 2007; Queiroga et al., 2007). Due to the importance of larval fish ecology to recruitment success, thus to enhance fish stocks, special attention is given to larval fish vertical migration.
Larval fish vertical dynamics
The fact that most of coastal fish species have a pelagic larval stage has important implications for the dynamics of fish populations and fisheries management (Leis, 2007). Management of coastal fishes must incorporate the scales over which their populations are connected due to dispersal processes and connectivity, therefore it is important to integrate coastal currents knowledge with
Chapter 1
6
larval behaviour, specifically vertical migration, to improve our understanding of larval survival, growth and dispersal (Fiksen et al., 2007).
Larval fish vertical migration is strongly determined by several factors (e.g. predator/prey interactions, bioenergetics and mortality, dispersal processes and connectivity) and much of the existent knowledge was obtained from studies focusing clupeid species, since its importance to fisheries and human populations (e.g. Reid, 2001; Santos et al., 2006; Voss et al., 2007). Clupeid larval stages usually feed on several planktonic species and are strongly predated by visual predators, namely adult fishes (Aksnes et al., 2004; Fiksen et al., 2007). Therefore, during day and night different depth distributions are usually observed, in order to minimize mortality rates without ignoring feeding requirements. Furthermore, according to the larval stage, different vertical positions are observed, since larvae depend upon different coastal currents to disperse and recruit (Hare and Govoni, 2005; Fiksen et al., 2007). Once more, different DVM patterns are observed for different species and larval stages. For example, Hare and Govoni (2005), for species that moved inshore or remained on the shelf, found that larvae were in deeper positions in the water column than larval from species that were exported from the shelf. Santos et al. (2006) found that sardine larvae vertical position was strongly associated to coastal currents, retaining the larvae in high food availability layers. The knowledge of how much energy planktonic species, specifically fish larvae, invest on predation risk and feeding opportunities help us to understand their ecology and life history strategies, as indicators of how much do adults invest in larval stages hence, offspring quality (Fiksen et al., 2007).
Work context
The work developed during this thesis constituted a special study as part of the oceanographic campaign “Ecosystem Survey Mozambique 2007” of the wider Nansen EAF Programme “Strengthening the Knowledge Base for and Implementing and Ecosystem Approach to Marine Fisheries in Developing Countries”. Sampling was made onboard the R/V “Dr. Fridtjof Nansen” and took place at the Sofala Bank, a wide shallow shelf influenced by the Zambezi River delta and other estuaries, and where the most important national fisheries occur. Besides important surface and river runoff discharges, strong tidal currents and currents from the Mozambique Channel have major effects on hydrological and occurring ecological processes (Ridderinkhof et al., 2001; Lutjeharms, 2006).
Studies from Sofala Bank are very scarce and only cruise and fisheries reports can be found (Bandeira et al., 2002, and references therein). It is known that this neritic region has high phytoplankton biomass comparing to the greater part of the Mozambican oligotrophic waters,
mainly because of the nutrient input from the Zambezi river runoff, which enhances the system productivity and supports local fisheries (Gammelsrød, 1992; Lutjeharms, 2006). Therefore, Sofala Bank is an excellent case study to understand the nutrients, phytoplankton, zooplankton and larval fish dynamics in relation to the coastal currents and physico‐chemical environmental variability.
Several studies identified tidal energy, light and diel variations as key factors regulating planktonic communities vertical distribution and diel changes (Hill, 1991; Queiroga et al., 2007; Brunet et al., 2008). At Sofala Bank, the tidal currents and estuarine discharges may strongly determine nutrients availability and water column light penetration due to increased turbidity, thus affecting base processes that modulate planktonic communities dynamics. Stratification of the water column, strongly condition the effectiveness of primary production magnitude at the surface layer and may act as a barrier to vertical movements of zooplankton (Criales‐Hernández et al., 2008). However, due to the strong local tidal currents, vertical mixing might locally induce homogenization of the water column. The investigation of tidal and diel variations and other regulating factors is therefore important to understand fundamental ecological processes that act in generating primary productivity and modulate resources supporting intense local fisheries activity.
Concerning larval fish dynamics, despite numerous worldwide studies on a great variety of species, specifically clupeids, none focused on the tropical herring Herklotsichthys quadrimaculatus (Rüppell 1837), also known as the gold‐spot herring. This species is a major component of tuna baitfish (Lewis, 1990) and consumed by local coastal communities at the Eastern Africa (KMFRI, 1981). At Mozambique, H. quadrimaculatus is broadly present on the coastal zone, namely at the Sofala Bank (INIPM, unpublished data). Knowledge of Herklotsichthys spp. larvae is very scarce (Thorrold and Williams, 1989), and for H. quadrimaculatus from the Western Indian Ocean only Harris and Cyrus (1999) described its high abundance throughout the year near Durban (South Africa). The vertical distribution and behaviour is still unknown for H. quadrimaculatus and information regarding ontogenic changes on depth distribution throughout diel variations are still missing. Furthermore, the hydrological unique properties of Sofala Bank may change the influence of typical factors regulating larvae vertical dynamics (e.g. light and currents variations). Tidal energy and vertical mixing, as well as other local oceanographic features, could therefore have a major role determining H. quadrimaculatus larvae vertical distribution and diel movements.
The scientific papers presented point toward a broadly understanding of the planktonic communities vertical dynamics. The first one (submitted to Estuarine, Coastal and Shelf Science)
Chapter 1
8
investigates the horizontal and vertical oceanographic and planktonic processes at the Sofala Bank shelf waters, specifically aimed to study currents, physico‐chemical water properties (temperature, salinity and nutrients), phytoplankton pigments as chemotaxonomic markers and zooplankton. The second paper (submitted to African Journal of Marine Science) is a specific study of fish larvae vertical dynamics, aimed to study the vertical distribution movements of H. quadrimaculatus larvae, its relation to several hydrological and biological factors (e.g. light variation, vertical mixing and zooplankton abundance) and also vertical distribution changes according to larval ontogeny.
References
Aksnes, D.L., Nejstgaard, J., Soedberg, E., Sørnes, T., 2004. Optical control of fish and zooplankton populations. Limnology Oceanography 49, 233‐238.
Anderson, D.M., Glibert, P.M., Burkholder, J.M., 2002. Harmful Algal Blooms and Eutrophication: Nutrient Sources, Composition, and Consequences. Estuaries 25, 704‐726.
Balkis, N., 2003. Seasonal variations in the phytoplankton and nutrient dynamics in the neritic water of Büyükçekmece Bay, Sea of Marmara. Journal of Plankton Research 25, 703‐717.
Bandeira, S.O., Silva, R.P., Paula, J., Macia, A., Hernroth, L., Guissamulo, A.T., Gove, D.Z., 2002. Marine Biological Research in Mozambique: Past, Present and Future. AMBIO 31, 606‐609. Brunet, C., Lizon, F., 2003. Tidal and diel periodicities of size‐fractionated phytoplankton pigment
signatures at an offshore station in the southeastern English Channel. Estuarine, Coast and Shelf Science 56, 833‐843.
Brunet, C., Gasotti, R., Vantrepotte, V., 2008. Phytoplankton diel and vertical variability in photobiological responses at a coastal station in the Mediterranean Sea. Journal of Plankton Research 30, 645‐654.
Calbet, A., Garrido, S., Saiz, E., Alcaraz, M., Duarte, C.M., 2001. Annual zooplankton succession in coastal NW Mediterranean waters: the importance of the smaller size fractions. Journal of Plankton Research 23, 319‐331.
Cottingham, K.L., 1999. Nutrients and zooplankton as multiple stressors of phytoplankton communities: Evidence from size structure. Limnology Oceanography 44, 810‐827.
Criales‐Hernández, M.I., Schwamborn, R., Graco, M., Ayón, P., Hirche, H.J., Wolf, M., 2008. Zooplankton vertical distribution and migration off Central Peru in relation to the oxygen minimum layer. Helgoland Marine Research 62, S85‐S100
Elliott, M., Burdon, D., Hemingway, K.L., Apitz, S.E., 2007. Estuarine, coastal and marine ecosystem restoration: Confusing management and science – A revision of concepts. Estuarine, Coastal and Shelf Science 74, 349‐366.
Epifanio, C.E., Garvine, R.W., 2001. Larval Transport on the Atlantic Continental Shelf of North America: a Review. Estuarine, Coastal and Shelf Science 52, 51‐77.
Fisken, Ø., Jørgensen, C., Kristiansen, T., Vikebø, F., Huse, G., 2007. Linking behavioural ecology and oceanography: larval behaviour determines growth, mortality and dispersal. Marine Ecology Progress Series 347, 195—205.
Relation to the Zambezi River Runoff. Estuarine, Coastal and Shelf Science 35, 91‐103.
Gibson, R.N., 2001. Go with the flow: tidal migration in marine animals. Hydrobiologia 503, 153‐161. Giraud, X., Quéré, C.L., Cunha, L.C., 2008. Importance of coastal nutrient supply for global ocean
biogeochemistry. Global Biogeochemical Cycles, 22, GB2025.
Hajdu, S., Höglander, H., Larsson, U., 2007. Phytoplankton vertical distributions and composition in Baltic Sea cyanobacterial blooms. Harmful Algae 6, 189‐205.
Hare, J.A., Govoni, J.J., 2005. Comparison of average larval fish vertical distributions among species exhibiting different transport pathways on the southeast United States continental shelf. Fisheries Bulletin 103, 728—736.
Harris, S.A., Cyrus, D.P., 1999. Composition, abundance and seasonality of fish larvae in the mouth of Durban harbour, Kwazulu‐Natal, South Africa. Sout African Journal of Marine Science 21, 19— 39.
Hill, A.E., 1991. Vertical migration in tidal currents. Marine Ecology Progress Series 75, 39‐54.
Irigoien, X., Conway, D.V.P., Harris, R.P, 2004. Flexible diel vertical migration behaviour of zooplankton in the Irish Sea. Marine Ecology Progress Series 267, 85‐97.
K.M.F.R.I., 1981. Aquatic Resources of Kenya. Proc. of the Workshop of Kenya Marine and Fisheries Research Institute, July 13‐19.
Kennedy, V.S., Twilley, R.R., Kleypas, J.A., Cowan, J.H., Hare, S.R., 2002. Coastal and marine ecosystems & Global climate change – Potential Effects on U.S. Resources. PEW Center on Global Climate Change.
Leis, J.M, 2007. Behaviour as input for modelling dispersal of fish larvae: behaviour, biogeography, hydrodynamics, ontogeny, physiology and phylogeny meet hydrography. Marine Ecology Progress Series 347, 185‐193.
Lewis, A.D., 1990. Tropical south Pacific tuna baitfisheries. In: Blaber, S.J.M., Copland, J.W. (Editors) Tuna baitfish in the Indo‐Pacific region. Aust. Coun. Int. agr. Res. Proc. 30, 10—21.
Lutjeharms, J.R.E., 2006. The costal oceans of south‐eastern Africa. In: A.R. Robinson, K.H. Brink (Editors) The Sea Vol 14. Harvard University Press, Cambridge, MA, pp. 783‐834. Ohman, M.D., 1990. The Demographic Benefits of Diel Vertical Migration by Zooplankton. Ecological Monographs 60, 257‐281. Paula, J., Pinto, I., Guambe, I., Monteiro, S., Gove, D., Guerreiro, J., 1998. Seasonal cycle of planktonic communities at Inhaca Island, southern Mozambique. Journal of Plankton Research 20, 2165‐ 2178. Queiroga, H., Almeida, M.J., Alpuim, T., Flores, A.A.V., Francisco, S., Gonzàlez‐Gordillo, I., Miranda, A.I., Silva, I., Paula, J., 2006. Tide and wind control of megalopal supply to estuarine crab populations on the Portuguese west coast. Marine Ecology Progress Series 307, 21‐36.
Queiroga, H., Cruz, T., dos Santos, A., Dubert, J., González‐Gordillo, J.I., Paula, J., Peliz, A., Santos, A.M.P., 2007. Oceanographic and behavioural processes affecting invertebrate larval dispersal and supply in the western Iberia upwelling ecosystem. Progress in Oceanography 74, 174‐191. Reid, D.G., 2001. SEFOS – shelf edge fisheries and oceanography studies: an overview. Fisheries
Research 50, 1—15.
Ridderinkhof, H., Lutheharms, J.R.E., Ruijter, W.P.M., 2001. A research cruise to investigate the Mozambique Current. South African Journal of Science 97, 461‐464.
Chapter 1
10
Santos, A.M.P., Ré, P., Dos Santos, A., Péliz, A., 2006. Vertical distribution of the European sardine (Sardina plichardus) larvae and its implications for their survival. Journal of Plankton Research 28, 523—532.Silva, A., Mendes, C.R., Palma, S., Brotas, V., 2008. Short‐time scale variation of phytoplankton succession in Lisbon bay (Portugal) as revealed by microscopy cell counts and HPLC pigment analysis. Estuarine, Coastal and Shelf Science 79, 230‐238.
Steinberg, D.K., Goldthwait, S.A., Hanself, D.A., 2002. Zooplankton vertical migration and the active transport of dissolved organic and inorganic nitrogen in the Sargasso Sea. Deep‐Sea Research 49, 1445‐1461.
Thorrold, S.R., Williams, D.M., 1989. Analysis of Otolith Microstructure to Determine Growth Histories in Larval Cohorts of a Tropical Herring (Herklotsichthys castelanaui). Canadian Journal Fisheries Aquatic Sciences 46, 1615—1624.
Tyrrell, T., 1999. The relative influences of nitrogen and phosphorus on oceanic primary production. Nature 400, 525‐531.
Verity, P.G., Yoder, J.A., Bishop, S.S., Nelson, J.R., Craven, D.B., Blanton, J.O., Robertson, C.Y., Tronzo, C.R., 1993. Composition, productivity and nutrient chemistry of a coastal ocean planktonic food web. Continental Shelf Research 13, 741‐776.
Voss, R., Schmidt, J.O., Schnack, D., 2007. Vertical distribution of Baltic sprat larvae: changes in patterns of diel migration?. ICES Journal Marine Science, 64.
Vrede,K., Vrede, T., Isaksson, A., Karlsson, A., 1999. Effects of nutrients (phosphorus, nitrogen, and carbon) and zooplankton on bacterioplankton and phytoplankton – a seasonal study. Limnology Oceanography 44, 1616‐1624.
Walther, G.‐R., Post, E., Convey, P., Menzel, A., Parmesan, C., Beebee, T.J.C., Fromentin, J.‐M., Hoegh.Guldberg, O., Bairlein, F., 2002. Ecological responses to recent climate change. Nature 416, 389‐395.
Wassmann, P., Olli, K., 2005. Drainage basin nutrient inputs and eutrophication: an integrated approach. University of Tromsø, Norway, 325 pp.
Wawrik, B., Paul, J.H., Bronk, D.A., John, D., Gray, M., 2004. High rates of ammonium recycling drive phytoplankton productivity in the offshore Mississippi River plume. Aquatic Microbial Ecology 35, 175‐184.
Zaret, T.M., Suffern, J.S., 1976. Vertical migration in zooplankton as a predator avoidance mechanism. Limnology Oceanography 21, 804‐813.
Distribution and vertical dynamics of planktonic communities at
Sofala Bank, Mozambique
M. C. Leal
1, C. Sá
2, S. Nordez
3, V. Brotas
2, J. Paula
11 Centro de Oceanografia, Laboratório Marítimo da Guia, Faculdade de Ciências da Universidade de Lisboa,
Av. Nª Senhora do Cabo, 939, 2759‐374 Cascais, Portugal
2 Centro de Oceanografia, Instituto de Oceanografia, Faculdade de Ciências da Universidade de Lisboa,
Campo Grande, 1749‐016 Lisboa, Portugal
3 Instituto de Investigação Pesqueira de Moçambique, Av. Mao Tse‐Tung nº 389, Maputo, C. P. 4603
Moçambique
Abstract
Coastal ecosystem processes are largely influenced by the interaction of different factors operating at various temporal and spatial scales, specifically those responsible for primary production patterns that modulate zooplankton and subsequent trophic levels. Hydrological processes, such as tidal cycles and coastal currents, nutrients availability, phytoplankton groups (studied through algal pigment signatures analysed by HPLC) and zooplankton abundance and distribution were investigated at the Sofala Bank (Mozambique), with special emphasis on their horizontal distribution and vertical dynamics (48‐hour). Horizontal distribution has shown onshore‐offshore gradients in all analysed parameters, as well as inshore waters intrusion probably related to Zambezi River delta runoff. Tidal currents were responsible for major hydrological vertical variations and for horizontal and vertical advection of phytoplankton biomass in the surface and deepest layers, respectively. Nutrient concentrations were typical from oligotrophic regions, and nutrient ratios were strongly influenced by depleted nitrite + nitrate concentrations, suggesting low estuarine discharges typical from the dry season. The very low N:P ratio obtained, suggests strong nitrogen limitation to phytoplankton communities, supporting the low phytoplankton abundance observed. Both phytoplankton pigments and zooplankton were found mainly near bottom (40 m deep), despite the latter displayed vertical migrations triggered by light variations. Phytoplankton community was dominated by microflagellates, specifically prymnesiophyceans, and behaved as a whole, except Cyanobacteria that displayed vertical distribution movements different from other phytoplankton groups, being mainly concentrated at mid water column depths (10 – 20 m). This investigation enhances
Chapter 2
14
physico‐chemical phenomena and their importance determining the planktonic communities vertical dynamics at a tropical coastal ecosystem of the Western Indian Ocean, where planktonic dynamics are still poorly described and understood.
Keywords: Phytoplankton; Pigments; Zooplankton; Vertical dynamics; Nutrient deficiency;
Mozambique
Introduction
Coastal shelf waters are ecosystems of great human and ecological interest where complex processes occur. The interaction of physical (e.g. coastal currents, upwelling, tides, advection), chemical (variable chemical properties including nutrient inputs) and ecological (e.g. biological production and its dynamics, prey/predator interactions) processes induce high spatial variability on the water column over different time scales (Brunet and Lizon, 2003; Lutjeharms, 2006; Queiroga et al., 2007). This variability determines the abundance and structure of different biological communities present in coastal waters, specifically phyto‐ and zooplankton as oceanic food chain lower levels. Zooplankton abundance and distribution is often related to predator/prey interactions, as a prey for both planktivorous fishes and some planktonic larvae, and as consumers of phytoplankton organisms (e.g. González‐Gordillo and Rodríguez, 2003; Santos et al., 2006). Phytoplankton is therefore regulated by zooplankton and fish herbivory, but also by nutrients availability, mainly nitrogen, phosphate and silica (Vanni, 1987; Svensson and Stenson, 1991; Garnier and Cugier, 2004). The relation between the different nutrient ratios and phytoplankton in coastal systems has been widely studied, and nitrogen generally plays a key role limiting phytoplankton growth thus strongly determining its community structure and chemotaxonomic pigment composition (Hecky and Kilham, 1988; Tyrrell, 1999).
The present study took place at the Sofala Bank (Mozambique), a wide shallow shelf influenced by the Zambezi River delta and other estuaries, and where important fisheries occur. Besides important river runoff discharges, strong tidal currents and currents from the Mozambique Channel have major effects on hydrological and occurring ecological processes (Ridderinkhof et al., 2001; Lutjeharms, 2006). Even though the high importance of this coastal ecosystem for Mozambican fisheries and economy, studies from Sofala Bank are very scarce and only cruise and fisheries reports can be found (Bandeira et al., 2002, and references therein). This neritic region has high phytoplankton biomass comparing to the greater part of the Mozambican oligotrophic waters, mainly because of the nutrient input from the Zambezi river runoff, which is
closely related to rainfall events (Lutjeharms, 2006). Hence, despite that hydrological and planktonic dynamics of several temperate coastal ecosystems are already studied (e.g. Verity et al., 1993; Balkis, 2003; Sabetta et al., 2008; Silva et al., 2008), Sofala Bank is an excellent case study to understand the nutrients, phytoplankton and zooplankton dynamics in relation to the physico‐chemical environmental variability of a tropical coastal ecosystem from the Western Indian Ocean, where scientific knowledge is very scarce.
Several studies identified tidal energy, light and diel variations as key factors changing neritic planktonic vertical distribution (Hill, 1991; Queiroga et al., 2007; Brunet et al., 2008). At Sofala Bank, the strong tidal currents and estuarine discharges may strongly determine nutrients availability and water column light penetration due to increased turbidity, thus affecting base processes that modulate planktonic communities dynamics. Stratification of the water column, specifically the pycnocline depth and its strength, condition the effectiveness of primary production magnitude at the surface layer and may act as a barrier to vertical movements of zooplankton (Criales‐Hernández et al., 2008). However, due to the strong local tidal currents it is probable that vertical mixing locally induces homogenization of the water column. The investigation of tidal and diel variations and other regulating factors is therefore important to understand fundamental ecological processes that act in generating high primary productivity and modulate resources supporting intense local fisheries activity.
This study was a special study as part of the 2007 campaign “Strengthening the Knowledge Base for and Implementing and Ecosystem Approach to Marine Fisheries in Developing Countries” of the Nansen EAF Programme, and its main goal was to investigate the horizontal and vertical oceanographic and planktonic processes in the Sofala Bank shelf waters, specifically the horizontal distribution and vertical temporal variation of (1) hydrological processes and parameters, (2) nutrients concentration, (3) phytopigments composition and abundance and (4) zooplankton abundance along the water column.
Chapter 2
16
Material and Methods
Sampling
The study area was located at the Sofala Bank, in front of the Zambezi river delta. Sampling took place on board the R/V Dr. Fridtjof Nansen from the 6th to the 8th December 2007, coveringin the first 24 hours a total of 11 stations distributed along three transects (Fig. 1), from a bottom depth of 24 to 374 m, and the outermost stations separated by 60 km. Their position was set as to characterize the shelf section where the fixed station sampling was carried out in the following 48‐hour period. The fixed station was positioned over a bottom depth of 50 m and samples were taken every 2 h.
Fig. 1. Radial and fixed stations location. Both radial () and fixed stations () are presented and the arrows indicate sampling order.
Current magnitude and direction were measured with a hull‐mounted Acoustic Doppler Current Profiler (ADCP) and transparency (m) using a Sechi Disk. At every radial and fixed station, CTD profiles were conducted and temperature, salinity and fluorescence (used as a proxy of chlorophyll a) data recorded at every 1 m depth interval. Water samples were taken at five predetermined depths (5, 10, 20, 30 and 40 m), except for shallower stations, using a rosette equipped with Niskin bottles. Immediately after collection, two replicates of 100 ml were collected and stored frozen for posterior nutrient analysis (nitrate + nitrite, phosphate and silicate). For HPLC pigment analysis, two litres of seawater were immediately filtered through glass fibre filters (25 mm φ, 0.7 µm pore ‐ Whatman GF/F) and filters were kept frozen in the dark for posterior analysis.
Depth‐stratified zooplankton samples (0‐5, 5‐10, 10‐20, 20‐30 and 30‐40 m) were collected with a multinet (Midi model, 0.5 x 0.5 m mouth size, Hydro‐bios) with 405 µm mesh size and towed at ~2 knots for 2 min, sampling on oblique hauls in each stratum. Flow rate was monitored by a flowmeter mounted in the mouth of the aperture, and each sample represented approximately 40 m3 of water filtered. A neuston net (0.2 x 1.0 m mouth size) with the same mesh
size and a flowmeter mounted was towed horizontally at similar speed and time, sampling the upper 20 cm of the water column. All zooplankton samples were preserved in 4% borax‐buffered formaldehyde, prepared using seawater.
Laboratory procedures
All the laboratory procedures were done at the Oceanography Centre of University of Lisbon, Portugal. Colorimetric analyses, with a Tecator FIAstarTM 5000 Analyser, were performed to determine nutrient concentrations. Nitrite (NO2‐), nitrate (NO3‐), phosphates (PO43‐, hereafter as
P) and silicates (Si(OH)4, hereafter as Si) were respectively determined according to Grasshoff
(1976), Bendshneider and Robison (1952), Murphey and Riley (1962), and Faning and Pilson (1973). Since water properties from this region are typically oligotrophic (Lutjeharms, 2006), the nitrite and nitrate sum was used (NO2‐ + NO3‐, hereafter as N).
The identification and abundance of phytoplankton functional groups can be achieved by high performance liquid chromatography (HPLC) analytical technique, which is increasingly in use as it is a less time consuming method in relation to microscopy phytoplankton identification and counting. HPLC quantifies chemotaxonomic pigments allowing to estimate the contribution of phytoplankton groups to chlorophyll a (Chl a) using photosynthetic marker pigments, such as alloxanthin for cryptophytes, 19’‐hexanoyloxyfucoxanthin for prymnesiophyceans, and other less specific biomarkers such as fucoxanthin for diatoms (also present in chrysophytes) and zeaxanthin for cyanobacteria (also present in green algae), among others (Jeffrey et al., 1997). In order to extract photosynthetic pigments, frozen filters were disrupted with 2 ml of 95% cold‐buffered methanol (2% ammonium acetate) for 30 min at ‐20º C in the dark. Samples were sonicated for 1 min in the beginning of the extraction period and then centrifuged at 4000 rpm for 15 min, at 4º C. Extracts were filtered (Millipore membrane filters, 0.2 µm) immediately before injection in the HPLC to remove cell and filter debris. Pigment extracts were analysed using a Shimadzu HPLC comprised of a solvent delivery module (IC‐10ADVP) with system controller (SCL‐10AVP) and a photodiode array (SPD‐M10ADVP). The chromatographic separation of pigments was achieved
Chapter 2
18
using the method described in Zapata et al. (2000), which uses a monomeric OS C8 column and a
mobile phase constituted by two solutions: methanol:acetonitrile:aqueous pyridine and, acetonitrile:acetone; a flow rate of 1 mL min‐1 and a run duration of 40 min. Pigments were
identified by comparison of retention times and absorption spectra with pure crystalline standards.
Biovolume, using sedimentation volumes with a conical jar and 24 h settling time, was measured to assess zooplankton abundance, estimated through planktonic organisms larger than 405 µm. Large gelatinous organisms (e.g. jelly fish) were removed because their significant buoyancy makes the method less precise (Postel et al., 2000).
Statistical analysis
In order to test for cyclic phenomena in the variation of the different parameters and their co‐ variations, autocorrelations and linear cross‐correlations were performed. Significant autocorrelations with a 2 h lag were analysed considering the 2‐hour sampling periodicity. Spearman correlation and Student’s t‐test were applied. The study of phytoplankton community was done through the analysis of chemotaxonomic pigments and their ratios to Chl a. Furthermore, the Fp pigment index (Claustre, 1994) was calculated in order to identify the trophic status of this marine ecosystem. The Fp pigment index is given by: Fp = (Σ fucoxanthin + Σ peridinin) x (Σ fucoxanthin + Σ peridinin + Σ 19’‐hexanoyloxyfucoxanthin + Σ 19’‐butanoyloxyfucoxanthin + Σ zeaxanthin + Σ chlorophyll b + Σ alloxanthin) ‐1 All the statistical analyses were carried out using R (R Development Core Team, 2008) while maps for display horizontal distributions were processed using Ocean Data View (Schlitzer, 2008).
Results
Horizontal distribution patterns
The horizontal variations of hydrological data evidenced the onshore‐offshore gradients (Fig. 2A and B). Despite the complex pattern of temperature and salinity distribution, the lowest temperatures were found in the most onshore stations and generally lower salinities were associated to warmer temperatures. However, the more coastal water mass showed higher salinity and lower temperature that the central water mass. Examining the vertical sections of thenorthern and southern transects (Fig. 3), a central less saline water mass with higher temperature was observed only at the surface layers, until 20 m deep. Despite the halocline of this structure was noticeable, vertical temperature profile was stratified along the water column. Fig. 2. Surface distribution of salinity (A) temperature (B, ºC), phosphates (C, µmol l‐1), silicates (D, µmol l‐1), total chlorophyll a (E, fluorescence data) and zeaxanthin (F, µg l‐1).
Chapter 2
20
Fig. 3. Vertical cross‐section of the northern (A) and southern (B) transect perpendicular to shore, as regards to salinity (1), temperature (2, ºC) and total chlorophyll a (3, fluorescence data) variation.
Concerning nutrients concentration, P was higher in the northern transect and Si concentration was higher near coast (Fig. 2C and D), while N was always under method detection limit (< 0.005 µmol l‐1). Shelf vertical cross sections of P concentration presented no vertical trend,
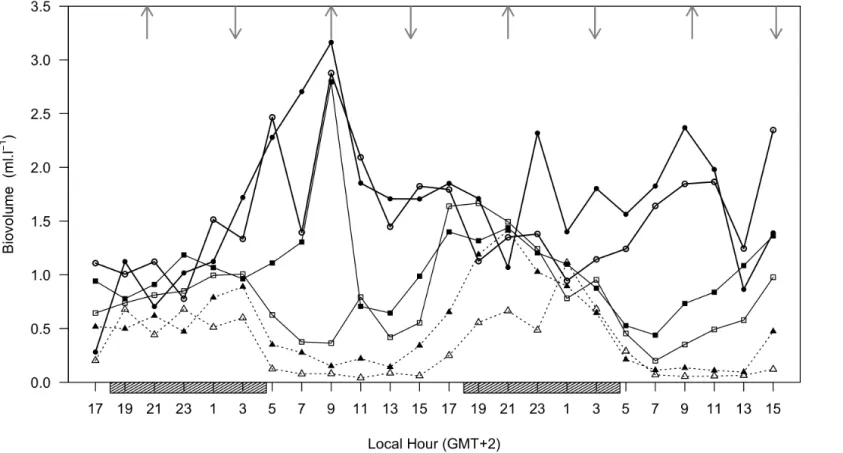
while for S higher concentrations on surface onshore water mass were observed (data not shown). Phytoplankton pigments presented very low concentrations in almost every radial station, with a distribution pattern similar to fluorescence data, where the southern and closest stations to the coast presented higher biomass signal, with a lowering gradient perpendicular to coast (Fig. 2E), and no different pigment composition at the central water mass. Vertical sections of the outermost transects displayed different phytoplankton distributions given by fluorescence signal (Fig. 3), with higher biomass on surface water mass nearer to coast on the southern transect, while in the northern transect higher values were obtain near the bottom, where a discontinuity occurs. Zeaxanthin concentration was the only pigment with a different spatial distribution, since higher concentrations were observed in the most northern and offshore stations (Fig. 2F). Zooplankton total abundance was generally higher in the northern transect (Fig. 4), apart from the most offshore station where total biovolume was extremely low, which was also observed in the other transects. The majority of night sampled stations (two northern transects) presented higher total zooplankton abundances than day stations.
Fig. 4. Horizontal distribution of zooplankton water column total abundance.
Vertical dynamics
Current direction and magnitude measurements were very similar throughout water column and results observed were in accordance with tidal variation (Table 1). Current direction varied mainly between South and West, respectively during flooding and ebbing tides, and speed was significantly higher during flood tides (t value = ‐ 4.7042, p < 0.01), with average current of 252 and 202 mm s‐1, respectively for flood and ebb tides. Minimum velocity values observed werecoincident with predicted tides at Chinde, near the Zambezi delta (Fig. 5), thus showing no significant delay to the sampling area. Current direction was similar in all sampling depths throughout the 48‐hour sampling period and the residual currents were generally over 150 mm s‐1 from South. However, current speed presented vertical variation without apparent consistence, i.e., in some periods higher values were at the 16 m deep while in others at 25 m. It should be noted that the ADCP only registered below the 16 m depth layer, thus results do not show the water mass behaviour above that depth. Water transparency varied mainly between 10 and 20 m deep, presenting no significant periodicity throughout sampling period. 35.5 36.0 36.5 37.0 37.5 -19.6 -19.4 -19.2 -19.0 -18.8 -18.6 -18.4 Biovolume Longitude Latitude 1.6 3.2 4.8 6.4 8 ml.l !1
Chapter 2
22
Table 1. Auto‐ and cross‐correlations of relevant hydrobiological parameters and their significanceParameter Periodicity (h) Significance Depth (m) (Cross ‐ ) correlated with Temperature 6 (opp.) Tide variation 20, 30*, 40* Current direction** (opp. phase; r = 0.681)
Salinity 6 (opp.) Tide variation 20 Current direction** (r = 0.681)
Fluoroscence 6 (opp.) Tide variation 30, 40 ‐
Current magnitude 12 Tidal cycle 0 – 30** ‐
Current direction 12 Tidal cycle All depths*** N* (r = 0.363)
NO2‐ + NO3‐ 6 (opp.) Tide variation 40 Si*** (r = 0.533)
Si(OH)4 6 (opp.) Tide variation 20* P* (r = 0.369)
diadinoxanthin (30 m: r = 0.289; 40 m: r = 0.213) fucoxanthin (30 m: r = 0.366; 40 m: r = 0.468)
Zeaxanthin 4 Water mass 20** ‐
Zooplankton 12 (opp.) Diel cycle All** (except 10/20 m) ‐
* p < 0.1, ** p < 0.05; *** p < 0.01; opp., opposed phase
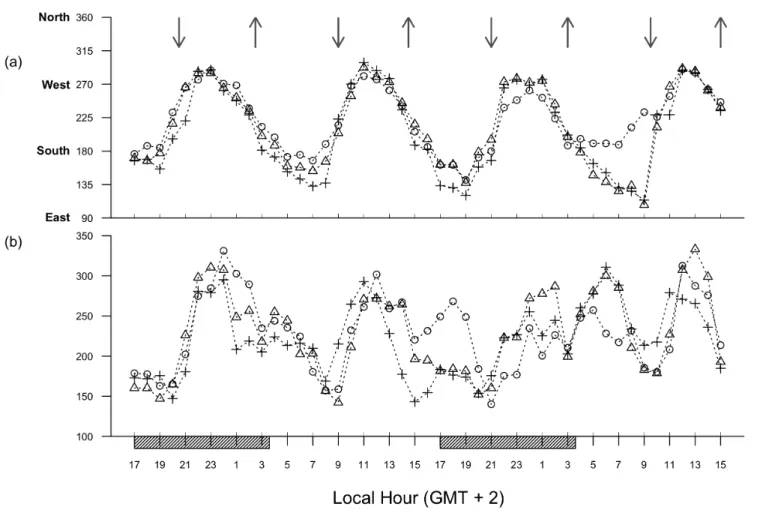
Fig. 5. Variation of 1‐h average currents direction (a, degrees) and magnitude (b, mm s‐1) at several depths ( ‐ 16 m, ‐ 25 m, ‐ 34 m). Upper and downward
Chapter 2
24
Fig. 6. Temperature (a, ºC) and salinity (b) variation during 48‐hour cycle. Upper and downward arrows indicate predicted high and low tides, respectively; shaded rectangles indicate night periods. The water column was vertically stratified below the thermocline during sampling period, as seemed clearly from salinity data (Fig. 6). Thermocline and halocline depths oscillated between 15 and 20 m for most of the cycle, and both temperature and salinity followed the same trend with a 6‐hour opposed‐phase periodicity (Table 1). During the low tide period it was generally observed less saline waters and warmer temperatures down to greater depths in the water column (Fig. 6). Throughout the last period of the 48‐hour cycle the water column physical properties did not show the same dynamics, specifically in the end of the second day where an intrusion of less saline waters was observed deeper (around 30 m).Nutrient concentrations ranged from 0 to 2.01 µmol l‐1 for N, 1.65 to 2.94 µmol l‐1 for P and
4.70 to 15.70 µmol l‐1 for Si (Fig. 7). N concentration was higher near the bottom during low tides,
which is consistent with a significant 6‐hour opposed‐phase periodicity and to the strong correlation found with current direction at the 40 m depth layer (Table 1). Despite Si concentration only showed tidal periodicity at shallower depths, it was positively correlated to N and P at 40 m deep (Table 1). Nutrient ratios N:P and N:Si were calculated and ranged both from 0
to 0.41 and 0.15, respectively. These ratios were strongly determined by N concentration that was generally under the method detection limit.
Fig. 7. Nutrient concentrations (µmol l‐1) during the 48‐hour cycle (a ‐ N;b ‐ P;c ‐ Si). Upper and downward arrows indicate predicted high and low tides, respectively; shaded rectangles indicate night periods.