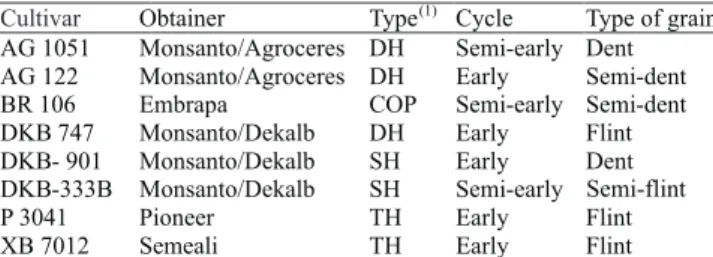
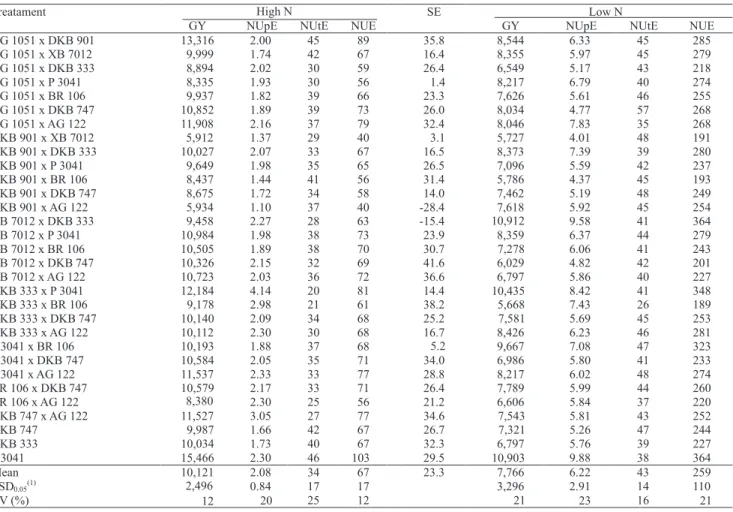
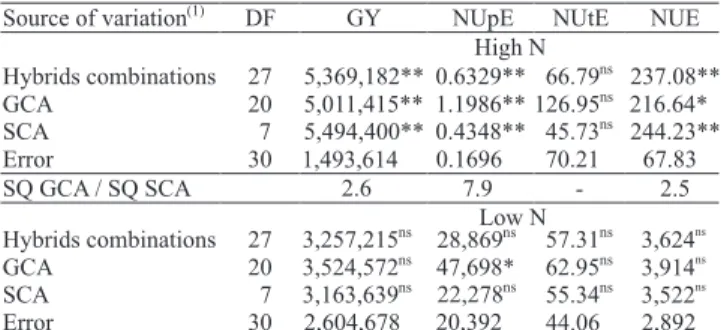
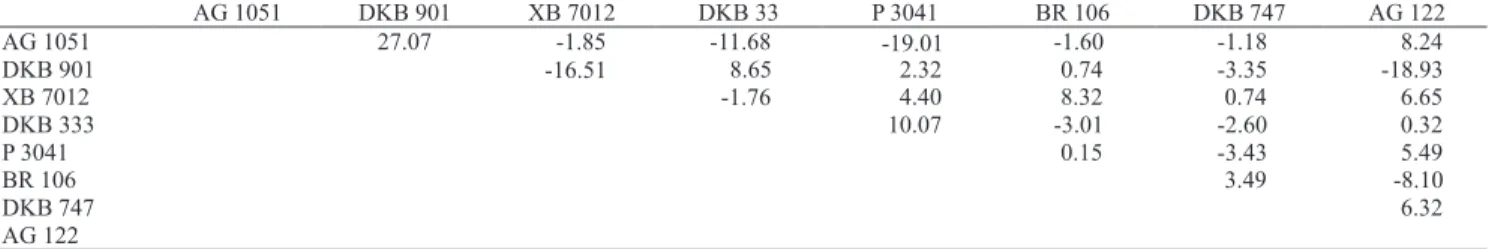
Genetic control of grain yield and nitrogen use efficiency in tropical maize
Texto
Imagem
Documentos relacionados
Em pacientes oncológicos ou em cuidados paliativos devemos agir com cuidado, enfatizando sua qualidade de vida, dor e a fadiga além dos já mencionados
Estimates and significance of the contrast between groups of dent and flint corn hybrids, for grain yield (GY), grain yield in silage maturity (GYS), vitreousness (VIT),
The aim of this study was to estimate the genetic progress attained for popping expansion (PE) and grain yield (GY) after four cycles of recurrent selection and to compare
The objective of this study was to evaluate the water use efficiency, agronomic efficiency of nitrogen, heat units and use of evapotranspiration of sunflower yield and its
A ideia de escrever um artigo deste género tem 4 objectivos principais: 1) dar a conhecer alguns aspectos do processo editorial aos participantes nos Congressos Anuais da
Abstract ‒ The objective of this work was to estimate genetic and phenotypic parameters among progenies of the CNA6 population of upland rice ( Oryza sativa ) for grain yield and
The nitrogen levels significantly affected dry weight, thousand grain weight and oil content in 2012, as well as plant height, number of pods, dry weight, grain yield and oil yield
Abstract – Grain productivities of 100 bean lines were evaluated in the presence and absence of nitrogen fertilizer in order to identify those with high nitrogen use efficiency
