Pró-Reitoria de Pós-Graduação e Pesquisa
Stricto Sensu em Ciências Genômicas e Biotecnologia
ANÁLISE MOLECULAR DOS IMPACTOS DO
CULTIVO DE DENDEZEIROS E DO
AMARELECIMENTO FATAL SOBRE AS
COMUNIDADES DE ARQUEIAS DE SOLO
AMAZÔNICO
Autora: Daiva Domenech Tupinambá
Orientadora: Prof
aDra. Betania Ferraz Quirino
Brasília - DF
DAIVA DOMENECH TUPINAMBÁ
ANÁLISE MOLECULAR DOS IMPACTOS DO CULTIVO DE
DENDEZEIROS E DO AMARELECIMENTO FATAL SOBRE AS COMUNIDADES DE ARQUEIAS DE SOLO AMAZÔNICO
Tese apresentada ao Programa de Pós-Graduação Stricto Sensu em Ciências Genômicas e Biotecnologia da Universidade Católica de Brasília, como requisito parcial para a obtenção do Título de Doutor em Ciências Genômicas e Biotecnologia.
Orientadora: Profa. Dra. Betania Ferraz Quirino.
7,5cm
Ficha elaborada pela Biblioteca Pós-Graduação da UCB
T928a Tupinambá, Daiva Domenech.
Análise molecular dos impactos do cultivo de dendezeiros e do amarelecimento fatal sobre as comunidades de arqueias de solo amazônico. / Daiva Domenech Tupinambá – 2015.
115 f.; il.: 30 cm
Tese (Doutorado) – Universidade Católica de Brasília, 2015.
AGRADECIMENTOS
Agradeço:
• A Dra. Betania Ferraz Quirino, por esses quatro anos de orientação; sua ajuda e apoio;
• Ao membros da banca: Dra. Cristine Chaves Barreto, Dra. Cynthia Maria Kyaw e Dr. Ricardo Henrique Kruger; por suas inestimáveis contribuições para este trabalho. Em especial, para a Dra. Cristine, quem sempre se fez presente nestes quatro anos;
• Ao Dr. Esdras, pela indicação da minha orientadora e seu apoio em um momento difícil;
• A Universidade Católica de Brasília e seus professores;
• A Embrapa Cerrados, por permitir meu crescimento profissional;
• A Embrapa Agroenergia, pelo apoio a este projeto de tese;
• Um agradecimento muito especial ao Dr. Maurício Egídio Cantão por ser um professor nato, além de um bioinformata excepcional e, principalmente, por dispor de seu tempo para me ajudar inúmeras vezes. Sem sua colaboração este trabalho não seria possível.
• Aos colegas de laboratório, em especial à Jessicão, que me recebeu e me treinou;
• Um agradecimento carinhoso à Betúlia, uma pessoa de gentileza infinita, que sempre está disponível para a ajudar a todos;
• Aos meus queridos Ohanets, Daniel/James, Tainá e Laninha: sem a sua amizade creio que este projeto não seria possível;
• Um agradecimento mais que especial para Ohana, minha companheira de bancada e desesperos diários, além de coautora e ás do Photoshop! O que seria deste trabalho sem suas figuras?
• Ao meu querido amigo Dr. Robson Sereno, por seu apoio e presença constante;
• Aos meus amigos e familiares, que (quase) sempre entenderam minhas ausências e me ajudaram a tornar essa caminhada mais leve;
• Ao Cláudio, grande companheiro de jornada, desde as noites tristes na orla de Fortaleza, até os dias felizes de hoje; irmão de sangue e alma, obrigada!
• A minha mãe, por sempre se interessar pelas “minhocas” e crer mais em mim do que eu mesma;
• Ao meu pai, não mais comigo em corpo, mas sempre ao meu lado em espírito; pensar em você nos momentos difíceis sempre torna tudo mais leve;
RESUMO
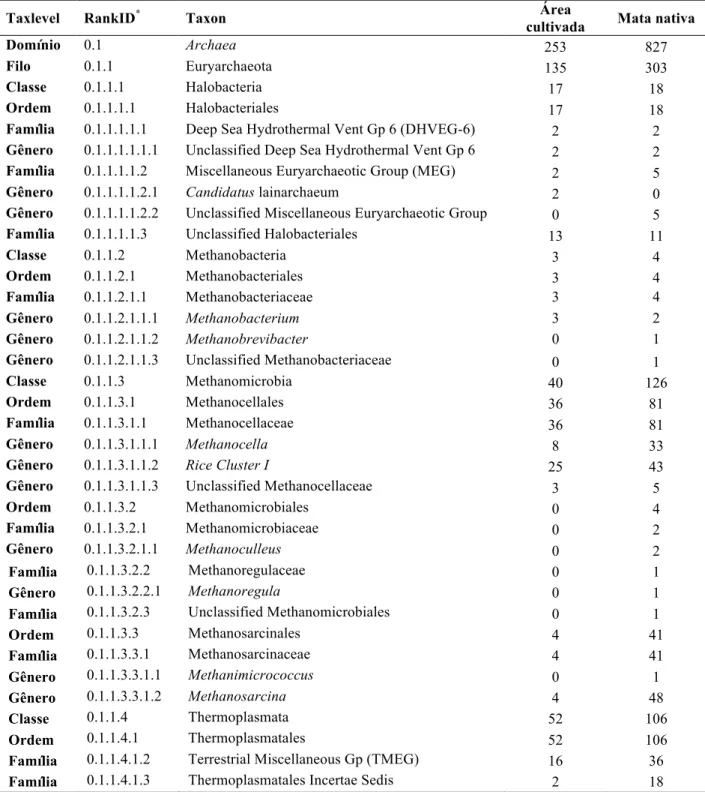
A floresta Amazônica é berço de enorme diversidade de macroespécies. Entretanto, pouco se sabe sobre a diversidade microbiana. O efeito do uso da terra após o desmatamento das florestas é de grande importância no desenvolvimento de políticas públicas. A partir do metagenoma extraído do solo de mata nativa Amazônica e de uma área adjacente cultivada com dendezeiros foi utilizado o pirosequenciamento do 16S rRNA das comunidades de arqueias presentes nesses solos para caracterização filogenética e análise comparativa das comunidades de arqueias. Todas as OTUs dos solos de mata nativa e área cultivada com dendezeiros foram classificadas em apenas dois filos: Euryarchaeota e Thaumarchaeota. O filo Thaumarchaeota foi predominante apenas na mata nativa, sendo Euryarchaeota, especialmente arqueias metanogênicas, predominantes nos solos cultivados com dendezeiros. Diversos gêneros envolvidos com os ciclos biogeoquímicos, como arqueias oxidadoras de amônia e metanogênicas, foram identificados nas duas amostras. Na mata nativa os gêneros classificados que apresentam a maior representação foram Candidatus Nitrosotalea e
Candidatus Nitrososphaera, AOAs. Já na área cultivada com dendezeiros o gênero de maior
representação foi Rice Cluster I. Foi encontrada um correlação direta entre os níveis de matéria orgânica e carbono total e a diversidade de arqueias nos solos amazônicos. Além disso, a antropização também apresentou impacto sobre essa diversidade. Este é o primeiro estudo de caracterização da microbiota de arqueias em solos amazônicos usando primers específicos e sequenciamento de alto desempenho. Este trabalho também caracterizou as comunidades de arqueias em solos cultivados com dendezeiros com e sem sintomas de Amarelecimento Fatal. O crescimento da demanda energética mundial e preocupação com as mudanças climáticas levou a um aumento da busca mundial por fontes alternativas de energia, o que está levando diversos países a buscarem na bioenergia uma alternativa. Entretanto, ainda existem diversas limitações na produção de biocombustíveis, seja na eficiência e custo do processo produtivo, seja na qualidade das fontes energéticas disponíveis. O dendê é uma das fontes mais promissoras de óleo para a produção de biodiesel no Brasil, sendo o Amarelecimento Fatal, doença com fator etiológico desconhecido, um limitante no uso do dendê. A partir do metagenoma extraído dos solos associados a dendezeiros com e sem sintomas de AF foi utilizado o pirosequenciamento do 16S rRNA das comunidades de arqueias para caracterização filogenética. Foi realizada uma análise comparativa das comunidades de arqueias de solos de dendezeiros com e sem sintomas de AF, numa tentativa de associação de algum microrganismo com essa doença. Na comparação entre dendezeiros com e sem sintomas de AF, os três grupos estudados diferiram entre si; o grupo 8 apresentou maior diversidade e obteve menor cobertura. Todos os grupos apresentaram dois filos: Thaumarchaeota e Euryarchaeota. Houve prevalência do segundo em todos os grupos, com aumento na abundância de arqueias metanogênicas com o AF. Na análise entre gêneros, foram observadas diferenças significativas entre os grupos, especialmente para os gêneros
Rice Cluster I e Ca. Nitrosotalea, que apresentaram um aumento em suas abundâncias
diretamente proporcional ao aumento dos sintomas do AF. Os gêneros Ca. Nitrososphaera e Methanocella apresentaram uma relação inversa; uma queda na abundância com o aumento dos sintomas do AF. Entretanto, não se pode afirmar que estes grupos estão relacionados ao AF. Este trabalho é complementar ao estudo da microbiota bacteriana desses solos, já realizado; e pelo estudo da microbiota fúngica, em andamento. Trata-se de um estudo inédito, que irá contribuir para os estudos futuros sobre o Amarelecimento Fatal.
ABSTRACT
The Amazon rainforest is home to huge diversity of macro-species. However, little is known about the microbial diversity. The effect of land-use after deforestation is of great importance in the development of public policies. The metagenome were extracted from soils of native forest and an adjacent cultivated area with oil palm and pyrosequencing of 16S rRNA genes of archaea communities present in those soils was used for phylogenetic characterization of the archaeal microbiota, in an unprecedented characterization of native Amazonian soil and soils cultivated with oil palm. All OTUs of the native forest soils and cultivated area with oil palm were classified into two phyla: Euryarchaeota and Thaumarchaeota. Thaumarchaeota phylum was predominant only in native forest. Euryarchaeota, especially methanogenic archaea, were prevalent in cultivated area with oil palm. Various genera involved in biogeochemical cycles, as AOA and methanogenic archaea, were identified in all samples. In native forest the genera with larger representation were Candidatus Nitrosotalea and
Candidatus Nitrososphaera, AOAs. In the cultivated area with oil palm the genus with larger
representation was Rice Cluster I. There is a direct correlation between levels of organic matter and total carbon and the diversity of archaea in Amazonian soils. In addition, anthropization also showed impact on this diversity. This is the first study to characterize the microbiota of archaea in Amazonian soils using specific primers and high-throughput sequencing. This work also characterize the archaeal communities in soils cultivated with oil palm with and without symptoms of Fatal Yellowing. The growth of world energy demand and concern with climate changes lead to a worldwide increase in the search for alternative sources of energy. Within this scenario, agroenergy presents itself as a viable alternative. However, there are still several limitations to the production of biofuels, such as efficiency and cost of the production process as well as the quality of the energy feedstock available. Palm oil is one of the most promising sources of oil for biodiesel production in Brazil, and the Fatal Yellowing (FY), a disease with unknown etiology, is limiting the use of palm. From the metagenome extracted from soils associated to oil palms with and without symptoms of FY was used pyrosequencing of 16S rRNA genes of archaeal communities for phylogenetic characterization, in an attempt of an association of some microorganism with FY, and an unprecedented characterization of soils cultivated with oil palms with and without FY. In the comparison among oil palms with and without FY symptoms, the three groups were different among then; group 8 showed higher diversity and had lower coverage. All groups presented two phyla: Thaumarchaeota and Euryarchaeota. There was prevalence of the second in all groups, with an increase in abundance of methanogenic archaea with FY. In the analysis of genera, significant differences between the groups were observed, especially for genera Rice
Cluster I and Ca. Nitrosotalea, which showed an increase in abundance directly proportional
to the increase of the FY symptoms. The genera Ca. Nitrososphera and Methanocella showed the opposite; a decrease in abundance with the increase of FY symptoms. However, it’s not possible to say that these genera are related to FY. This work is complementary to the study of bacterial microbiota of these soils, already performed; and the study of fungal microbiota, in progress. This is an unpublished study, which will contribute to future studies on the Fatal Yellowing.
LISTA DE ILUSTRAÇÕES
Figura 1 – Locais de produção das principais oleaginosas com potencial para produção de
biodiesel no Brasil. ... 21
Figura 2 - Dendezeiro adulto. ... 24
Figura 3 - Dendezeiro acometido por AF em estágio 8 e folhas de dendezeiro com sintomas de AF. ... 25
Figura 4 – Árvore filogenética universal. (Fonte:(WOESE, 2000)). ... 30
Figura 5 - Desenho esquemático da amostragem dos solos. ... 40
Figura 6 - Reações de PCR da amostra 8.3.1 usando os primers 340f/1000r... 48
Figura 7 - Gel de agarose 1% da quantificação da amostra 8.3.1 purificada... 48
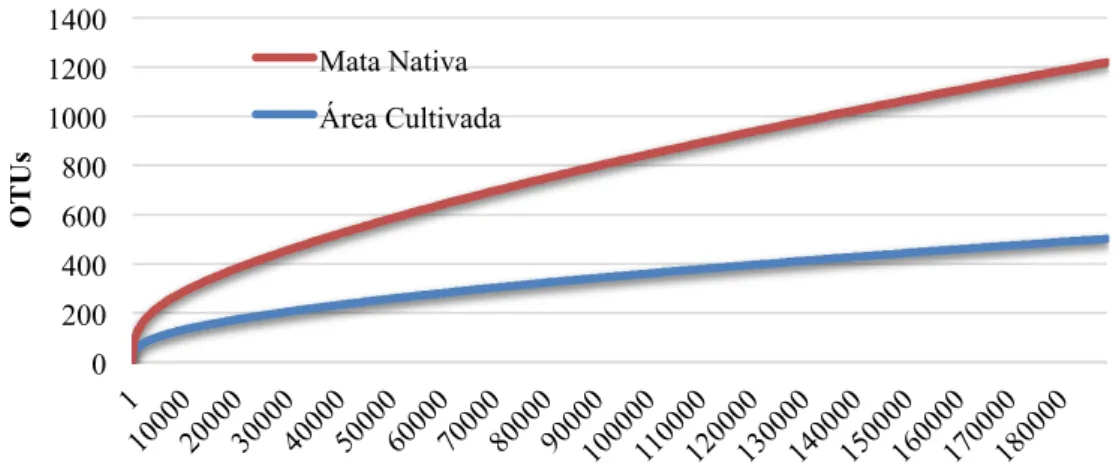
Figura 8 – Curvas de rarefação das sequências e OTUs observadas nas amostras de mata nativa (vermelho) e área cultivada com dendezeiros (azul), por MOTHUR. ... 49
Figura 9 - Número de OTUs compartilhadas e únicas nas amostras de mata nativa e área cultivadas com dendezeiros... 52
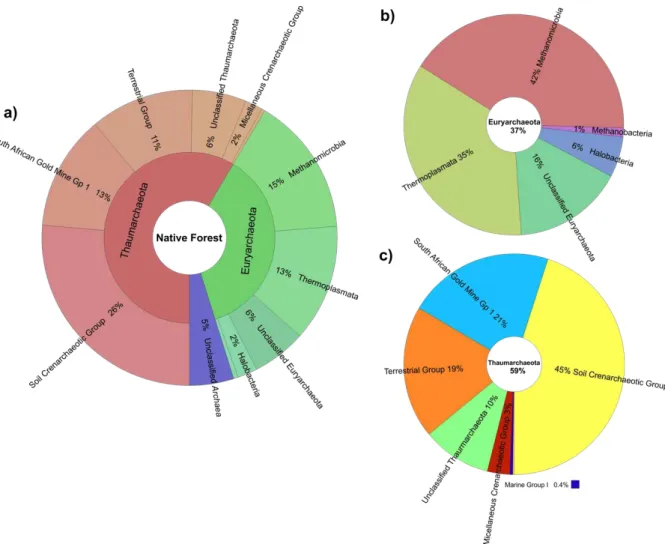
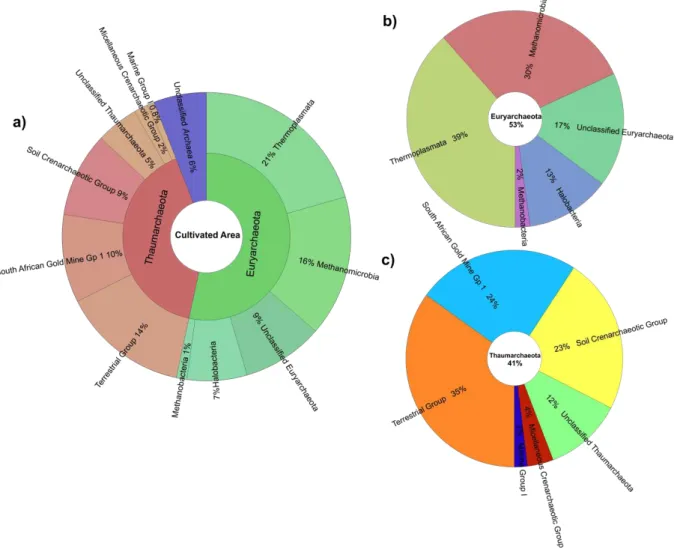
Figura 10 –Percentuais de participação das OTUs classificadas a 97% de similaridade em até o nível de classe: (a) na amostra de solo de mata nativa; (b) no filo Euryarchaeota e, (c) no filo Thaumarchaeota, por Krona. ... 53
Figura 11 – Percentuais de participação das OTUs classificadas a 97% de similaridade em até o nível de classe: (a) na amostra de solo da área cultivada com dendezeiros; (b) no filo Euryarchaeota e, (c) no filo Thaumarchaeota, por Krona. ... 54
Figura 12 – Árvore filogenética construída a partir de sequências de 16S rRNA classificadas como unclassifiedThaumarchaeota das amostras de mata nativa (NF) e área cultivada com dendezeiros (CA), por MEGA6. ... 56
Figura 13 – Perfil taxonômico comparativo das OTUs classificadas em até o nível de classe das amostras de mata nativa (em laranja) e área cultivada (em azul) computadas pelo MOTHUR. ... 57
Figura 14 – Percentuais de OTUs classificadas em até o nível de gênero nas amostras: (a) de mata nativa e (b) área cultivada com dendezeiros, por Krona. ... 57
LISTA DE TABELAS
LISTA DE ABREVIATURAS
AF – Amarelecimento Fatal
AOA – arqueia oxidadora de amônia
Ca. - Candidatus
DCA – dendezeiros com AF
DGGE - eletroforese em gel com gradiente desnaturante
DNA - ácido desoxirribonucleico
DSA – dendezeiros sem sintomas de AF
EDTA – ácido etilenodiamino tetra-acético
g – constante de gravidade
g – grama
ITS – Intergenic Transcribred Spacer
kb – quilobase
kDa – quilo Dalton
kg – quilograma
LB – Luria Bertani
m/v – massa por volume
mg – miligrama
mL – mililitro
ORF – open reading frame
OTU – operational taxonomic unit
pb – pares de bases
PCA – Análise de Componentes Principais
PCR – Reação em cadeia da polimerase
RISA – Ribosomal Intergenic Spacer Analysis
RNA – ácido ribonucleico
SDS – dodecil sulfato de sódio
TE – tampão Tris-EDTA
v/v – volume por volume
β – beta
µg – micrograma
SUMÁRIO
AGRADECIMENTOS ... 5
RESUMO ... 7
ABSTRACT ... 8
LISTA DE ILUSTRAÇÕES ... 9
LISTA DE TABELAS ... 12
LISTA DE ABREVIATURAS ... 13
SUMÁRIO ... 14
1. Introdução ... 15
2. Revisão Bibliográfica ... 17
2.1. Região Amazônica ... 17
2.2. Biodiesel, Dendê e Amarelecimento Fatal ... 20
2.3. Biodiversidade e a Biologia Molecular ... 28
2.4. Arqueias ... 30
3. Objetivos ... 38
3.1. Objetivos Gerais ... 38
3.2. Objetivos Específicos ... 38
4. Materiais e Métodos ... 39
4.1. Coleta de Solo ... 39
4.2. Análises Físico-Químicas do Solo ... 40
4.3. Extração do DNA Total do Solo ... 41
4.4. PCR do gene 16S rRNA e Pirosequenciamento ... 42
4.5. Bioinformática ... 43
5. Resultados ... 47
5.1. Análise das comunidades de arqueias de solos de mata nativa versus área cultivada com dendezeiros ... 47
5.1.1. Análises Químicas do Solo ... 47
5.1.2. PCR do gene 16S rRNA de arqueias ... 47
5.1.3. Curvas de Rarefação ... 48
5.1.4. Índices de Riqueza e Diversidade ... 49
5.2. Análise das Comunidades de Arqueias de Solos de Dendezeiros com e sem
Sintomas de AF ... 60
5.2.1. Análises Químicas dos Solos ... 60
5.2.2. Análise da Correlação Entre os Dados Químicos e Filogenéticos ... 60
5.2.3. Curvas de Rarefação ... 61
5.2.4. Índices de Riqueza e Diversidade ... 62
5.2.5. Análises Filogenéticas ... 63
6. Discussão e Conclusões ... 76
6.1. Análise das Comunidades de Arqueias de Solos de Mata Nativa versus Área Cultivada com Dendezeiros ... 76
6.2. Análise das Comunidades de Arqueias de Solos Cultivados com Dendezeiros com e sem Sintomas de AF ... 85
REFERÊNCIAS BIBLIOGRÁFICAS ... 91
ANEXOS ... 109
Anexo A – PCA dos dados físico-químicos dos solos de dendezeiros com e sem sintomas de AF. ... 109
Anexo B – Gráfico das análises físicas das 18 amostras de solos de dendezeiros. ... 110
Anexo C – Resultados das análises químicas para solos associados à dendezeiros com e sem sintomas de AF e mata nativa. ... 111
Anexo D – Amostras sequenciadas, MIDs utilizadas e nomenclatura das amostras para as análises ... 112
Anexo E – Número de sequências observadas, cobertura de Good, índices de riqueza e diversidade para as amostras de mata nativa, área cultivada com dendezeiros e grupos 0, 5 e 8113 Anexo F - Perfis taxonômicos comparativos das OTUs não normalizadas classificadas em nível de classe dos grupos 0 , 5 e 8. ... 114
1. INTRODUÇÃO
A diversidade microbiana se refere à quantidade de indivíduos de determinada espécie e à riqueza de espécies de fungos, bactérias e arqueias existentes em determinado habitat (ALTIERI, 1999; NANNIPIERI et al., 2003). Os microrganismos são responsáveis pela ciclagem de nutrientes no solo, o que os caracteriza como essenciais para a homeostase de um ecossistema (MOREIRA; SIQUEIRA, 2006). Práticas de cultivo, como aragem e adubação de solos, podem modificar a estrutura das comunidades de microrganismos, desequilibrando essa microbiota (BISSETT et al., 2011). Além disso, a conversão de áreas nativas em solos cultivados, especialmente com monoculturas, pode levar à diminuição dos teores de nitrogênio e oxigênio nos solos (DUPONT et al., 2010). Devido ao papel determinante dos microrganismos na qualidade dos solos, o estudo dos mesmos se torna importante; entretanto, apenas uma pequena parcela desses microrganismos é hoje cultivada em laboratórios, o que torna os estudos independentes de cultivo uma ferramenta poderosa no estudo dessas microbiotas.
O crescimento da demanda energética mundial e preocupação com as mudanças climáticas estão levando diversos países a buscarem na bioenergia uma alternativa para suas demandas energéticas. Segundo o Ministério do Meio Ambiente, o uso de biocombustíveis em substituição aos combustíveis fósseis deverá evitar o lançamento de cerca de 570 milhões de toneladas de CO2 na atmosfera entre 2008/2017, no Brasil (BRASIL, 2015). O biodiesel é uma fonte de energia renovável, disponível e sustentável.
O biodiesel é um biocombustível líquido derivado de biomassa, um éster de óleos ou gorduras de diversas origens, produto de uma reação de transesterificação entre um triacilglicerol e um álcool, na presença de um catalisador, gerando glicerol como subproduto; já o diesel é um subproduto do petróleo. O biodiesel pode usar como matéria-prima óleos vegetais, óleos e gorduras residuais (restos de fritura) ou ainda, gorduras animais, além de etanol ou metanol e catalisadores. No Brasil, o dendê apresenta grande potencial na produção de biodiesel, tanto por sua alta produtividade quanto pela qualidade superior de seu óleo.
Entretanto existe um problema para a implementação do dendê como fonte principal de óleo para a produção de biodiesel: o Amarelecimento Fatal (AF).
principais características dessa doença são: amarelecimento dos folíolos basais das folhas intermediárias (3, 4, 5 e 6), seguido por aparecimento de necroses nas extremidades dos folíolos, com evolução para a seca total dessas folhas, especialmente da folha flecha. A doença vem crescendo de maneira exponencial, exterminando plantações industriais no Pará, levando a centenas de demissões.
Até os dias de hoje ainda não há um fator etiológico conhecido do Amarelecimento Fatal; mesmo após a realização de diversos estudos, nas mais diversas áreas, a doença segue sem uma causa definida. Como um dos sintomas do AF é o apodrecimento das raízes (DE SOUZA, VEIGA; RAMOS, 2000; DE FRANQUEVILLE, 2003), seria possível a presença do patógeno, se o AF tiver origem biótica, na rizosfera.
Existem poucas informações sobre a diversidade de Archaea em solos; especialmente em solos Amazônicos. Tampouco sobre os papéis dessas arqueias, seja nos solos ou na doença. Neste sentido, a caracterização das comunidades de arqueias dos solos de dendezeiros afetados com a doença e sua comparação com a microbiota “saudável” pode fornecer pistas de um possível agente etiológico biótico.
Esta tese caracterizou e comparou as microbiotas de arqueias associadas a solos amazônicos de mata nativa e cultivados com dendezeiros, além da caracterização da microbiota de arqueias de solos cultivados com dendezeiros com e sem sintomas de AF, complementando um estudo inédito. Este trabalho faz parte de um projeto maior, que visa a caracterização das microbiotas fúngica, bacteriana e de arqueias em solos e folhas de dendezeiros com e sem sintomas do Amarelecimento Fatal.
A caracterização da diversidade bacteriana foi realizada por Jéssica C. Bergman, em sua tese de doutorado, onde foi encontrada uma maior riqueza no grupo 5; além disso, os gêneros Erwinia, Pseudomonas e Bacillus, já isolados de folhas de dendezeiros com AF e considerados supostos agentes etiológicos do AF, foram encontrados em todos os grupos estudados, mas em menor abundância relativa no grupo 0 (BERGMANN, 2013). Todavia, o agente etiológico do AF não pode ser identificado, mas a caracterização inicial da microbiota bacteriana desses solos agrega informações relevantes que serão de grande utilidade em estudos posteriores.
2. REVISÃO BIBLIOGRÁFICA
2.1. Região Amazônica
O Brasil possui cerca de 3,6 milhões de km2 de florestas, estando em primeiro lugar no mundo em área de florestas tropicais, sendo, de acordo com o IBGE, a grande maioria dessas florestas concentradas na região Amazônica, com a Amazônia legal perfazendo cerca de 60% de todo o território brasileiro (IBGE, 2014).
É igualmente impressionante a biodiversidade de macroespécies – de 10 a 20% de toda a riqueza animal e vegetal conhecida até o momento (VERÍSSIMO, CAPOBIANCO; GROSS, 2004).
Segundo a classificação climática de Köppen-Geiger, a região amazônica apresenta três regiões climáticas distintas: (1) clima tropical chuvoso, sem estação seca definida (Af); (2) clima tropical chuvoso, com um período de seca (Am), e (3) clima tropical chuvoso, com uma estação seca bem definida (Aw).
O solo é formado por materiais orgânicos e minerais, sendo o primeiro encontrado em diversos estados de decomposição, provenientes de restos vegetais e animais, substâncias húmicas e outros materiais orgânicos, além de biomassa microbiana. Estes se encontram associados a materiais minerais em proporções variáveis; os quais são formados por compostos inorgânicos diversos, encontrados em diversos estágios de intemperismo (SANTOS et al., 2006). A região amazônica apresenta predominantemente oxisols e ultisols distróficos, segundo a classificação norteamericana (SILVA, 1996). A Embrapa desenvolveu o sistema brasileiro de classificação de solos, que classifica os solos amazônicos como latossolos amarelos na sua maioria, com presença de argissolos distróficos (SANTOS et al., 2006); sendo o primeiro caracterizado por solos envelhecidos, ácidos a fortemente ácidos, de boa drenagem e baixa fertilidade natural; enquanto que os últimos são pobres e ácidos. Eventualmente pode-se encontrar solos eutróficos, do tipo Terra Roxa.
levam a um empobrecimento ainda maior destes solos, que não tardam a serem abandonados, gerando as chamadas “capoeiras”, um tipo de vegetação secundária.
A deposição de matéria orgânica no solo, proveniente da cobertura vegetal na região Amazônica, é parte importante da ciclagem de nutrientes e manutenção dos nutrientes dos solos, em especial de solos pobres, como é o caso dos solos amazônicos (LUIZÃO; SCHUBART, 1987).
Segundo o Projeto de Monitoramento do Desmatamento na Amazônia Legal por Satélite (PRODES), realizado pelo Instituto Nacional de Pesquisas Espaciais (INPE), foram desmatados 4.848 km2 da Amazônia Legal no período de agosto de 2013 a julho de 2014, uma redução de 18% em relação ao período anterior (INPE, 2015). O projeto PRODES conta com o apoio do IBAMA, Ministérios da Ciência, Tecnologia e Inovação (MCTI) e Ministério do Meio Ambiente (MMA). Entretanto, apesar dos esforços governamentais, ainda existe desmatamento da região Amazônica, colocando em risco a sua biodiversidade.
Apesar da floresta Amazônica ser conhecida por sua biodiversidade de macroespécies, ainda não existem muitos estudos dedicados à sua diversidade microbiana, em especial de arqueias. Os impactos da conversão da floresta amazônica em pasto foram analisados, como a queda da biomassa microbiana após a prática de desmatamento seguida de queima da cobertura vegetal (LUIZAO, BONDE; ROSSWALL, 1992), assim como o aumento da liberação de CO2, um dos gases responsáveis pelo efeito estufa (FEARNSIDE; IMBROZIO BARBOSA, 1998).
Em relação aos impactos sobre as comunidades microbianas, a influência dos fatores físico-químicos dos solos e uso da terra sobre as comunidades bacterianas nos mais diversos tipos de solos está bem estabelecida, tanto pela microbiologia clássica, quanto por abordagens moleculares, como técnicas independentes de cultivo e sequenciamento de alto desempenho (YAN, MCBRATNEY; COPELAND, 2000; ZHONG; CAI, 2007; SHENG et al., 2013; PAN et al., 2014).
a estes solos antropogênicos. Foi observada, ainda, homogeneização biótica das comunidades de bactérias dos solos amazônicos após o cultivo, com aumento da diversidade taxonômica e filogenética local, mas as comunidades se apresentam mais similares entre as diversas áreas adjacentes comparadas (RODRIGUES et al., 2012). Um estudo recente sobre o impacto do uso da terra sobre as comunidades microbianas de solos amazônicos encontrou alterações não apenas na composição e abundância dessas comunidades, mas também na diversidade de genes funcionais na conversão de floresta em pasto (PAULA et al., 2014).
Já as comunidades fúngicas não são tão bem estudadas quanto as bacterianas; mas, devido ao seu papel importante como decompositores, vários estudos focam em suas enzimas para uso biotecnológico, seja para a produção de biocombustíveis (OKAMOTO et al., 2012), revisado por (MACDONALD, SUZUKI; MASTER, 2012); ou, ainda, em outros usos biotecnológicos, como na indústria química e de alimentos (BAFFI et al., 2012).
A região amazônica, por seu clima quente e úmido, favorece a proliferação de fungos. Os solos amazônicos também possuem grande diversidade de fungos de interesse biotecnológico, como para a produção de pigmentos (CELESTINO et al., 2014).
As micorrizas arbusculares também são bem estudadas por serem de interesse agronômico. Esse tipo de fungo cria associações simbióticas mutualistas com as raízes das plantas e outros órgãos subterrâneos; onde as plantas recebem água e nutrientes minerais diretamente dos fungos e estes recebem açúcares provenientes da fotossíntese. Entretanto, este relacionamento pode nem sempre ser benéfico para as duas partes envolvidas, sendo ainda alvo de estudos (BENDER, DA SILVA VALADARES; TAUDIERE, 2014). Estes fungos também são estudados em solos amazônicos (LEAL, STURMER; SIQUEIRA, 2009).
As ações antrópicas também afetam as comunidades fúngicas dos solos. O estudo desses impactos é importante, já que os fungos possuem papéis importantes na ciclagem de nutrientes e decomposição de matéria orgânica nos solos. Em um estudo das comunidades fúngicas de solos de florestas primárias e secundárias, plantações e pasto usando PCR-DGGE, além de sequenciamento de clones de uma biblioteca de 18S rRNA, pode-se observar mudanças nas comunidades fúngicas com as ações antrópicas (FRACETTO et al., 2013).
Entre os domínios da vida, Archaea é o menos estudado, sendo assim o mais interessante a ser explorado. Até o momento, apenas um trabalho de ecologia microbiana focou a diversidade de arqueias em solos amazônicos, e o estudo usou um tipo bem específico de solos – a Terra Preta de Índio (TAKETANI; TSAI, 2010).
primeiro estudo independente de cultivo de comunidades microbianas de solos amazônicos do Pará. Todavia, foram usando primers universais para 16S rRNA (530F/1494R) para amplificar o DNA de solos de floresta e pasto, com apenas um representante do domínio
Archaea, do filo Crenarchaeota, encontrado em ambas amostras (BORNEMAN; TRIPLETT,
1997). As comunidades de arqueias associadas a solos antropogênicos do estado de Manaus foram estudadas usando DGGE, comparando-as àquelas de solos adjacentes (GROSSMAN et al., 2010). As diferenças entre os perfis das amostras foram extremas, chegando à 90%. Porém, poucas OTUs foram encontradas, todas do filo Crenarchaeota; mas ao serem reanalisadas com o banco de dados SILVA 119, essas OTUs foram classificadas como AOAs do filo Thaumarchaeota.
É importante ressaltar que existem poucos estudos usando sequenciamento de alto desempenho da Amazônia, como, por exemplo, análises genômicas de bactérias, fungos e vegetação amazônicos (RODRIGUES, SERRES; TIEDJE, 2011; MUELLER et al., 2014).
2.2. Biodiesel, Dendê e Amarelecimento Fatal
O Brasil apresenta grande potencialidade para a produção de biodiesel, com produção de diversas oleaginosas pelo país (Figura 1) (BERGMANN et al., 2013). Na atualidade, o estado do Pará é o principal produtor de dendê do Brasil.
Segundo a Organização das Nações Unidas para a Alimentação e Agricultura - FAO (2012), houve um aumento nas plantações destinadas à produção de biocombustíveis no mundo de 68,3 milhões de toneladas em 2006 para 130 milhões de toneladas em 2011, ocupando cerca de 45 milhões de hectares de terras cultiváveis. Um olhar mais detalhado a respeito do setor agropecuário leva à conclusão de que a participação do óleo diesel no setor deverá aumentar até 2030: de 57%, em 2004, para aproximadamente 70%, em 2030 (BRASIL, 2007). Em 2013, a oferta interna de energia apresentou uma taxa de crescimento de 4,5%, com o setor dos transportes liderando essa demanda (EPE, 2014).
sentido de aumentar a produtividade das matérias-primas disponíveis para baratear os custos, sem aumento das áreas cultiváveis.
Figura 1 – Locais de produção das principais oleaginosas com potencial para produção de biodiesel no Brasil (BERGMANN et al., 2013).
Tabela 1 - Informações sobre diversas oleaginosas com potencial agroenergético.
Espécie % de óleo Ciclo de vida
Rendimento (kg óleo/ha)
Referências
Dendê 22 Perene 2.000 a 8.000 (MAPA, 2013)
Pinhão manso 38 Perene 1.200 a 1.500 (DURÃES et al., 2009)
Coco 58 a 65 Perene 1.300 a 1.500 (CGEE, 2004)
Babaçu 60 Perene 120 (DE SÁ PARENTE, 2003)
Mamona 39,6 a 59,5 Anual 470 a 810 (MAPA, 2013)
Amendoim 40 a 60 Anual 788 (MAPA, 2013)
Girassol 40 a 47 Anual 774 (SEBRAE, 2009)
Soja 18 a 21 Anual 560 (MAPA, 2013)
Algodão 18 a 20 Anual 361 (MAPA, 2013)
O documento “Diretrizes de Política de Agroenergia 2006 - 2011” (BRASIL, 2005) destaca as seguintes diretrizes:
“(1) desenvolvimento da agroenergia...pela implantação da cadeia produtiva do biodiesel; (2) agroenergia e produção de alimentos...não afetará a produção de alimentos para o consumo interno; (3) desenvolvimento tecnológico...agregação de valor aos produtos e redução de impactos ambientais;
(4) ...deverá contribuir para a inserção econômica e social; (5) autonomia energética comunitária e geração de emprego e renda, e otimização do
aproveitamento de áreas antropizadas...recuperação de áreas degradadas.”
A cultura do dendê está totalmente alinhada com essas diretrizes, sendo uma fonte limpa de biodiesel, de maior produtividade, quando comparada com outras espécies oleaginosas, além de ajudar na fixação do homem no campo, trazendo renda para a agricultura familiar.
O Brasil é um país de grande extensão territorial, em grande parte formado por áreas cultiváveis, além de possuir um clima favorável para o cultivo de diversas oleaginosas com potencial para a produção de biodiesel.
Em meados de 2004 foi criado no Brasil um programa intitulado “Programa Nacional de Produção e Uso do Biodiesel” – PNPB, tendo por objetivos a inclusão social, desenvolvimento regional e geração de renda e empregos por meio do cultivo de diversas oleaginosas com potencial para a produção do biodiesel (BRASIL). Outras medidas de incentivo foram tomadas no âmbito do programa, como a obrigatoriedade de adição de um percentual de biodiesel ao diesel.
A Lei nº 11.097, de 13 de setembro de 2005, foi responsável pela introdução do biodiesel na matriz energética brasileira e assim o conceitua:
“Biocombustível derivado de biomassa renovável para uso em motores a combustão interna com ignição por compressão ou, conforme regulamento, para
geração de outro tipo de energia, que possa substituir parcial ou totalmente
combustíveis de origem fóssil.” (NR)
Além disso, esta Lei estabelece percentuais mínimos de mistura de biodiesel ao diesel. Em 2008 foi instituído o B3 – adição de 3% de biodiesel no diesel. Em janeiro de 2010, este percentual foi ampliado para 5%. Recentemente, em setembro de 2014, o percentual foi ampliado para 7%.
A cultura do dendê atende às diretrizes do documento “Diretrizes de Política de Agroenergia 2006 - 2011” (BRASIL, 2005), sendo uma fonte limpa de biodiesel, de maior produtividade quando comparada com outras espécies oleaginosas, além de ajudar na fixação do homem no campo, trazendo renda para a agricultura familiar (CGEE, 2010).
nutricional, fonte de vitamina E e betacaroteno e de baixo custo de extração, com diversas aplicações nas indústrias alimentícia e de cosméticos, e também pode ser usado na produção de biodiesel (BERGMANN et al., 2013). O dendê é capaz de gerar renda a partir de uma produção sustentada e ecologicamente limpa, necessitando de mão de obra intensiva, fixando agricultores no campo, além de proteger o solo, prevenindo a degradação das terras e erosão e ainda, oferecendo alta taxa de sequestro de carbono (VENTURIERI et al., 2009).
Figura 2 - Dendezeiro adulto.
Fonte: Jessica C. Bergmann.
De acordo com o Anuário Estatístico da Agroenergia 2012, no ano de 2010 foram cultivados no Brasil cerca de 112.500 ha com dendezeiros (MAPA, 2013).
Entretanto, o dendê ainda não se estabeleceu no Brasil como principal fonte de óleo para a produção de biodiesel. O Amarelecimento Fatal, doença sem fator etiológico conhecido, vem crescendo de maneira exponencial, exterminando plantações industriais no Brasil e em diversos países sul americanos. Mais de 5.000 hectares de dendezais foram dizimados somente no estado do Pará (VENTURIERI et al., 2009). Souza e colaboradores (2000) elaboraram uma cartilha para a identificação do AF no campo, e criaram uma escala de severidade dos sintomas da doença, de 1 a 10.
sintomatologia, como o apodrecimento fétido do meristema das palmeiras que ocorre na Pudrición del Cogollo mas não no Amarelecimento Fatal; a velocidade da progressão da doença e casos de remissão (DE FRANQUEVILLE, 2003).
As principais características dessa doença são: amarelecimento dos folíolos basais das folhas intermediárias (3, 4, 5 e 6), seguido por aparecimento de necroses nas extremidades dos folíolos, com evolução para a seca total dessas folhas, especialmente da folha flecha (Figura 3). O sistema radicular é fortemente afetado, apresentando necrose, juntamente com o início dos sintomas foliares (FREIRE, 1988; BOARI, 2008).
Figura 3 - Dendezeiro acometido por AF em estágio 8 e folhas de dendezeiro com sintomas de AF.
Fonte: Jéssica C. Bergmann.
Diversos estudos foram realizados desde a década de 80 no sentido de identificar as causas do AF. Ramos e colaboradores (2006) estudaram o comportamento de dendezeiros ao AF em áreas de replantio sob diferentes formas de manejo na Denpasa, empresa do Pará, onde foi primeiramente identificada a doença no Brasil.
Viégas e colaboradores estudaram as concentrações de diversos nutrientes nas folhas de dendezeiros sadios, com AF e de plantas resistentes (híbrido E. oleifera x E. guineensis), onde encontraram uma relação entre a concentração de ferro, cobre, zinco e manganês e o aparecimento de AF (VIÉGAS et al., 2000). Com base nesse trabalho, outro estudo foi realizado, para comparar os efeitos da adubação foliar com e sem omissão de nutrientes sobre a evolução dos sintomas de AF em dendezeiros. Os resultados obtidos levaram a uma recomendação de adubação foliar com formulação completa, com exceção de nitrogênio e fósforo, para a área estudada, onde se pode inferir que o AF não está associado à toxicidade por ferro ou outros micronutrientes. Para testar a hipótese de que o AF seja causado por baixa disponibilidade de ferro à planta devido à má absorção, características do solo, irrigação excessiva, baixa drenagem ou interação com outros micronutrientes, os pesquisadores realizaram outro experimento, expondo as raízes primárias das plantas à soluções com sulfato ferroso, sem nenhuma variação na evolução dos sintomas da doença (SILVEIRA et al., 2000).
Foi investigada a ocorrência de nematóides associados a tecidos de dendezeiros com AF, não sendo encontradas formas fitoparasitas de nematóides que pudessem ser relacionadas com a doença, sendo descartada a hipótese de que nematóides sejam causadores da doença (SILVA et al., 1996).
Vírus e viróides foram estudados por meio de transmissões mecânicas e uso de extratos de tecidos infectados, sem sucesso aparente (TRINDADE, POLTRONIERI; FURLAN JÚNIOR, 2005).
Celestino Filho e colaboradores (1993) não encontraram uma relação entre a fauna de insetos com áreas afetadas com AF no Pará.
Fungos e bactérias foram isolados de diversos tecidos de dendezeiros acometidos por AF e inoculados em plantas adultas e jovens. As inoculações foram realizadas em plantas submetidas a estresses hídrico e físico, usando suspensão de tecido de flechas e zona meristemática de plantas acometidas por AF, através de enxertos de pedaços de ráquis de plantas doentes em sadias e por meio do uso de ferramentas de despalma, cortando plantas doentes e sadias, respectivamente (SILVA, 1989). Após um ano de testes, não foram obtidos resultados conclusivos.
Deve-se observar que os microrganismos usados nesses estudos foram isolados e cultivados in vitro, ou seja, perfazem cerca de 1% da microbiota associada ao dendezeiro. Uma abordagem independente de cultivo ainda não foi utilizada e pode ter sucesso em correlacionar microrganismos da microbiota associada a dendezeiros com AF ao aparecimento dessa doença.
Uma prática não convencional de manejo da doença foi utilizada na Denpasa em 1992, onde o solo, após a remoção de plantas doentes, foi queimado antes do replantio, o que retardou o reaparecimento da doença em mais de um ano, em comparação à áreas não queimadas, o que pode sugerir que o solo foi descontaminado. Sendo assim, é possível imaginar que o patógeno responsável possa estar presente no solo contaminado (DE FRANQUEVILLE, 2003).
Um estudo de 2008, realizado na Colômbia, isolou de folhas de dendezeiros doentes e identificou o fungo Phytophthora palmivora como responsável pela Pudrición del Cogollo (SARRIA et al., 2008). Existem relatos de sintomas diferentes entre dendezeiros doentes de diversas regiões da América Latina (AYALA, 2001), de maneira que é possível que o Amarelecimento Fatal e a Pudrición del Cogollo não sejam a mesma doença. Além disso, neste trabalho se extraiu DNA de fungos isolados das folhas doentes e foram utilizados frutos-armadilha, sem o uso de sequenciamento de alto desempenho, nem DNA total extraído das folhas, sendo possível que algum patógeno não cultivável nesses frutos-armadilha não tenha sido identificado (MARTÍNEZ et al., 2010).
de Phythophtora seja encontrada nos dendezeiros brasileiros. Para tal, o DNA total dos solos associados a dendezeiros deve ser investigado em busca de representantes desse gênero.
2.3. Biodiversidade e a Biologia Molecular
Os seres vivos foram divididos em dois grandes grupos: os procariontes e os eucariontes, tendo por base a organização celular. Com o desenvolvimento da análise de sequências de DNA e proteínas, essa divisão, puramente fenotípica, mostrou-se inexata. As arqueias são procariontes, mas, no nível molecular, não mostram mais similaridade com outros procariontes do que com os eucariontes (WOESE, KANDLER; WHEELIS, 1990).
A capacidade de isolar bactérias do ambiente e cultivá-las em meios controlados ampliou o conhecimento desses seres e, então, veio uma visão reducionista da biologia, onde uma entidade biológica ou sistema devia ser separado em suas partes constituintes para melhor entendimento (WOESE, 2004). Entretanto, os organismos vivos vivem em biomas, interagindo entre si, dentro de um equilíbrio dinâmico.
Assim como as bactérias, os fungos também desempenham importantes funções ecológicas (CHRISTENSEN, 1989). Contudo, o entendimento destes é menor comparado ao das comunidades bacterianas, já que, até pouco tempo, a maioria dos micologistas baseavam seus estudos em métodos de cultura (ANDERSON; CAIRNEY, 2004). O mesmo ocorre com as arqueias, que apresentavam poucos estudos, na sua grande maioria de extremófilas cultiváveis, gerando, assim, poucos dados sobre a diversidade e papéis biológicos desses microrganismos. Entretanto, com o avanço dos estudos, as arqueias tem sido encontradas em diversos ambientes diferentes, se mostrando ubíquas, assim como fungos e bactérias.
Whitman e colaboradores, em 1998, estimaram o número de procariontes na Terra em 4 – 6.1030 células, distribuídas especialmente na água e solos, onde são essenciais nos processos geoquímicos (WHITMAN, COLEMAN; WIEBE, 1998). Hoje já é sabido que apenas cerca de 1% dos microrganismos encontrados na biota podem ser cultivados in vitro (AMANN, LUDWIG; SCHLEIFER, 1995). A metagenômica pode ser uma grande aliada para contornar essa barreira.
fundo do mar, corpo humano, solo, além de ambientes extremos, como minas ácidas (ALLEN; BANFIELD, 2005; JIAO et al., 2006; DEMANECHE et al., 2009; HUANG et al., 2009; PETROSINO et al., 2009).
A diversidade microbiana tem sido exaustivamente estudada, explorando diferentes ambientes. Em meados da década de 80, Peace e colaboradores (1986) propuseram a ideia de clonar DNA ambiental em hospedeiros e em 1991 publicaram seu primeiro estudo em que o DNA de uma amostra ambiental foi diretamente clonado em vetores fagos (SCHMIDT, DELONG; PACE, 1991). Uma biblioteca metagenômica de água do mar foi criada, onde foi identificada uma sequência de 16S rRNA proveniente de uma arqueia jamais cultivada anteriormente (STEIN et al., 1996). Tamames e colaboradores (2010) mostraram que vários táxons podem ser encontrados em diferentes ambientes. Este estudo também mostra que o solo é um ambiente pouco restritivo, englobando elevado número de táxons procarióticos.
Diversas técnicas foram desenvolvidas (PCR, clonagem molecular, sondas de DNA, sequenciamento de alto desempenho), dando novo impulso aos estudos filogenéticos. Entretanto, para cada nova técnica, vieses metodológicos são adicionados em cada passo físico, químico ou biológico (VON WINTZINGERODE, GOBEL; STACKEBRANDT, 1997). Como, por exemplo, na coleta de amostras, armazenamento e transporte, onde pode ocorrer enriquecimento das amostras (ROCHELLE et al., 1994) ou até mesmo contaminação do DNA. Na extração de DNA pode ocorrer extração preferencial ou, ainda, degradação, na tentativa de extração de todo o material presente na amostra (VON WINTZINGERODE, GOBEL; STACKEBRANDT, 1997). A Reação em Cadeira da Polimerase (PCR) possui diversos erros e vieses potenciais, gerando artefatos ou, ainda, amplificação preferencial (ACINAS et al., 2005). A escolha de primers também é crítica para as análises, já que sua especificidade é comum, podendo ocorrer amplificações preferenciais, com perda de representatividade de grupos de raros, por exemplo. A Eletroforese em Gel com Gradiente Desnaturante (DGGE) tem sido muito utilizada na geração de perfis moleculares (molecular
fingerprinting), tendo como vantagens a possibilidade de analisar diversas amostras
KANG, 2014). A própria escolha do método de análise a ser empregado é foco de divergências nos resultados (CASAMAYOR et al., 2002).
2.4. Arqueias
Em meados da década de 70, Woese e colaboradores propuseram uma nova divisão do mundo vivo em três grandes reinos – Archaebacteria, Urkaryonte e Eubacteria, tendo como base as diferenças moleculares entre esses grupos (WOESE; FOX, 1977). Mas foi no início da década de 90 que Woese e colaboradores sugeriram modificações de nomenclatura, especialmente a troca de Archaebacteria para Archaea, para evitar confusão entre arqueias e bactérias, tendo em vista diversos estudos que mostraram que as arqueias formam um novo domínio, tendo, por vezes, mais similaridades em nível molecular com os eucariontes do que com as bactérias (KELMAN, 2000). Sendo assim os domínios passaram a chamar-se
Archaea, Eukarya e Bacteria (WOESE, KANDLER; WHEELIS, 1990) (Figura 4). Nesta
árvore pode-se observar que as arqueias possuem um ancestral comum mais próximo com o Domínio Eukarya do que com o Bacteria.
Figura 4 – Árvore filogenética universal. (Fonte:(WOESE, 2000)).
alta salinidade ou pressão (ROTHSCHILD; MANCINELLI, 2001). Seus papéis biológicos ainda são pouco conhecidos, assim como sua diversidade não está bem estabelecida, já que até pouco tempo alguns poucos exemplares haviam sido cultivados em laboratório, sendo extremófilos em sua maioria, o que nos deu poucas pistas a respeito de seus papéis em ambientes como o solo (SCHLEPER, JURGENS; JONUSCHEIT, 2005). Entretanto, estudos independentes de cultura começam a revelar informações relevantes sobre estes microrganismos (BINTRIM et al., 1997; SIMON, DODSWORTH; GOODMAN, 2000).
Em 1996 foi sequenciado o primeiro genoma completo de arqueia:
Methanocaldococcus jannaschii (BULT et al., 1996). Hoje, em bancos de dados como o
UCSC Archaeal Genome Browser (CHAN et al., 2012) são disponibilizados 120 genomas
completos (acesso em 12/2014). O Global Genome Biodiversity Network (GGBN), um banco de DNA, conta hoje com 319 espécies de arqueias e 503 amostras de DNA (disponível online
em http://www.dnabank-network.org;acesso em 12/2014).
As arqueias são procariontes, mas, no nível molecular, não mostram mais similaridade com outros procariontes do que com os eucariontes (WOESE, KANDLER; WHEELIS, 1990). As arqueias possuem, no caso de sua maquinaria de replicação, um tipo de RNA polimerase semelhante às enzimas eucarióticas (HUET et al., 1983); além de histonas, presentes em alguns grupos, que compartilham um ancestral comum eucariótico (REEVE et al., 2004). Apresentam uma composição de parede celular variável, diferenciada das membranas celulares eucarióticas, como, por exemplo, no caso da arqueia Sulfolobus acidocaldarius, que apresenta uma parede celular com composição nunca antes encontrada (GUAN et al., 2011). Entretanto, os genes das proteínas ribossomais das arqueias estão organizados em operons, de maneira semelhante às bactérias (YANG et al., 1999).
A classificação taxonômica das arqueias vem sofrendo constantes adições, especialmente com o aumento dos estudos filogenéticos. Inicialmente, o domínio Archaea foi descrito por Woese e colaboradores, na década de 70 (WOESE; FOX, 1977). Posteriormente, o mesmo grupo sugeriu a separação do domínio em dois filos, Crenarchaeota e Euryarchaeota (WOESE, KANDLER; WHEELIS, 1990). Entretanto, como era de se esperar, novos grupos estão sendo propostos, como os filos Thaumarchaeota (BROCHIER-ARMANET et al., 2008) e Korarchaeota (BARNS et al., 1996).
O filo Crenarchaeota, sendo um dos primeiros a serem propostos, possuía representantes em vários ambientes, como o solo e mares (OCHSENREITER et al., 2003), além dos termófilos extremos, que suportam temperaturas superiores a 80oC. Seu nome veio do Grego “crenos”, origem. Entretanto, com o avanço dos estudos filogenéticos, seus representantes estão sendo alocados em novos filos, como no caso de representantes mesofílicos como o Cenarchaeum symbiosum (HALLAM et al., 2006), inicialmente reconhecido como representante do filo Crenarchaeota e, posteriormente, do filo Thaumarchaeota (BROCHIER-ARMANET et al., 2008).
Os representantes do Filo Thaumarchaeota são um grupo de arqueias predominantemente mesofílicas de difícil cultivo, compreendendo todas as arqueias oxidadoras de amônia (AOA), além de indivíduos de metabolismo ainda desconhecido (PESTER, SCHLEPER; WAGNER, 2011). “Thaumas”, do grego “maravilha”, foi o nome escolhido para esse novo filo por Brochier-Armanet e colaboradores em 2008, quando, usando não apenas análises de 16S rRNA, mas também marcadores genéticos, mostraram, por meio de árvores filogenéticas, que Cenarchaeum symbiosum, uma arqueia mesofílica que havia sido recentemente descoberta (HALLAM et al., 2006), era, na verdade, o primeiro membro deste grupo a ter seu genoma publicado (BROCHIER-ARMANET et al., 2008). Este filo está divido em três grupos: I.1a, formado por arqueias mesofílicas encontradas em diversos habitats (DELONG, 1998); I.1b: formado por AOAs do solo, águas termais, água doce e seus sedimentos (ZHALNINA et al., 2014); e I.1c: arqueias mesofílicas do solo, amplamente encontradas em florestas (JURGENS, LINDSTROM; SAANO, 1997b; OCHSENREITER et al., 2003; HU et al., 2013). O grupo I.1a possui um representante cultivado, Nitrosopumilus maritimus (WALKER et al., 2010).
O filo Nanoarchaeota foi descoberto em 2002, quando uma arqueia de genoma diminuto (~490 kb) foi cultivada juntamente com seu simbionte do gênero Igniccocus e denominada Nanoarchaeum equitans (HUBER et al., 2002). Esta arqueia é termofílica (90oC), vivendo na presença de enxofre, em uma atmosfera de H2 e CO2 (HOHN, HEDLUND; HUBER, 2002). Entretanto, sua classificação filogenética ainda é incerta, com novos estudos mostrando que artefatos podem surgir de classificações filogenéticas dentro de um domínio ainda “em construção” (BROCHIER et al., 2005).
Hoje já se sabe que as arqueias possuem papéis importantes nos ciclos geobioquímicos (BERG et al., 2010; MEYER-DOMBARD, AMEND; OSBURN, 2013), revisado por (OFFRE, SPANG; SCHLEPER, 2013). Venter e colaboradores, estudando o metagenoma do mar do Sargaço, observaram que altas concentrações de nitritos podem ser resultado da oxidação da amônia por parte de arqueias (VENTER et al., 2004). O que veio a ser comprovado em 2005, com o isolamento de uma arqueia marinha pertencente ao filo Crenarchaeota, com atividade nitrificante (KONNEKE et al., 2005). Com cerca de 20% de todos os procariotos nos mares pertencendo ao domínio Archaea, fica clara sua importância (KARNER, DELONG; KARL, 2001).
Treusch e colaboradores, no mesmo ano, encontraram não apenas arqueias marinhas, mas também componentes terrestres mesofílicos do filo Crenarchaeota capazes de oxidar amônia; não apenas em condições aeróbias, mas também, potencialmente, em condições anaeróbias (TREUSCH et al., 2005). A participação das arqueias entre os procariontes do solo é de 1 a 5% em suas camadas superficiais (BATES et al., 2011). Posteriormente, essas arqueias foram enquadradas em um novo filo, Thaumarchaeota, onde estão classificados todos os oxidadores de amônia nos grupos I.1a e I.1b (STAHL; DE LA TORRE, 2012).
Estudos com base em estudos filogenéticos e comparativos usando o gene amoA, que codifica a subunidade alfa da enzima amônia monooxigenase, envolvida na oxidação da amônia, mostram novas possíveis classificações para essas AOAs (PESTER et al., 2012); revisado por (STAHL; DE LA TORRE, 2012).
Fica claro que muito ainda há por ser descoberto em relação a este domínio; o uso de técnicas como a genômica unicelular associadas à metagenômica na análise de habitats ainda pouco conhecidos pode nos ajudar a elucidar os relacionamentos filogenéticos e evolutivos microbianos (RINKE et al., 2013)
tendo em vista sua distribuição universal, funcionalidade constante, além de serem conservados e apresentarem comprimento adequado para prover uma visão profunda dos relacionamentos evolutivos dos organismos em estudo. Além disso, existem extensos bancos de dados de 16S rRNA, como o Ribosomal Database Project (RDP), disponível online em
http://rdp.cme.msu.edu ou, ainda, o SILVA Database com base de dados de 16S/18S rRNA,
disponível online em http://arb-silva.de (COLE, 2007; QUAST et al., 2013). Entretanto, apesar desses genes serem úteis neste tipo de análise, o nível de variação entre estes genes é normalmente insuficiente para uma classificação em níveis mais específicos, como espécie.
Hoje é fato que os microrganismos presentes nos mais diversos habitats ainda são, em grande parte, desconhecidos. O uso de técnicas como o sequenciamento de alto desempenho ou, ainda, a metagenômica, nos revela novos genomas, muitas vezes de microrganismos ainda não cultivados, agregando novos grupos aos já conhecidos.
Na rizosfera, as plantas apresentam um série de interações com raízes de outras plantas e com a microbiota do solo. Estas comunidades microbianas são sustentadas pelas grandes quantidades de carbono orgânico produzidas pelas raízes (BUEE et al., 2009). Elas são influenciadas por exsudatos liberados pelas raízes, em um fenômeno chamado “rizodeposição”, onde, além da liberação de exsudatos, ocorre a produção de mucilagens e deposição de células das raízes. Inclusive, as próprias características físico-químicas destes solos podem ser modificadas por esses exsudatos (BAIS et al., 2006). Essa influência das plantas sobre a microbiota do solo pode ser tanta que estudos falam em “solos supressores de doenças” (disease-suppressive soils), onde a incidência de diversas doenças é menor, se comparada a solos adjacentes, mesmo com a presença do patógeno nesses solos (WELLER et al., 2002). Um estudo sobre os mecanismos moleculares de defesa das rizobactérias contra patógenos das raízes cita alguns mecanismos: desde a produção de antibióticos pela microbiota residente contra patógenos até a estimulação e suporte seletivo de populações antagonistas destes patógenos pelas plantas (COOK et al., 1995).
Existem diversos atores na rizosfera, como vírus, fungos, bactérias e arqueias (SUGIYAMA et al., 2014).
Em um estudo usando DGGE, a liberação de exsudatos das raízes foi relacionada com a composição das comunidades microbianas da rizosfera (YANG; CROWLEY, 2000).
microbiota do solo e são modificados por ação dos exsudatos vegetais (BENDER, DA SILVA VALADARES; TAUDIERE, 2014).
Raízes de milho produzem exsudatos capazes de atrair uma rizobactéria colonizadora benéfica, Pseudomonas putida, que compete com bactérias patogênicas (NEAL et al., 2012).
Em um trabalho sobre as arqueias aderidas às raízes de tomateiros usando DGGE, observou-se que cerca de 40% da variação entre as comunidades associadas às raízes pode ter sido ocasionada por mudanças na composição dos exsudados em virtude do status nutricional de ferro das plantas (SIMON, DODSWORTH; GOODMAN, 2000).
Por outro lado, também existem patógenos nos solos, como fungos, bactérias, nematóides e vírus. O pirosequenciamento tem sido utilizado com sucesso no estudo da presença de possíveis patógenos em plantas, como, por exemplo, no caso da identificação de vírus ou, ainda, no estudo de bactérias simbiontes de insetos vetores de doenças de plantas (AL RWAHNIH et al., 2009; HIRSCH et al., 2012; ROY et al., 2013).
Até o momento, não existe confirmação da existência de arqueias patogênicas; entretanto, em face do pouco conhecimento deste domínio, é possível que possam existir arqueias patogênicas ainda não identificadas em plantas.
Cavicchioli e colaboradores (2003), em um trabalho de revisão, analisaram diversos fatores que levariam a uma patogenicidade em arqueias:
a) exemplos de associação de arqueias com doenças; b) abundância das arqueias na natureza;
c) existência de interações entre arqueias e eucariontes; d) evidências de respostas imunes a arqueias;
e) arqueias habitando hospedeiros em número significativo, e
f) arqueias com estruturas moleculares semelhantes àquelas encontradas em patógenos.
Estes itens foram reavaliados à luz do conhecimento atual e com os dados deste trabalho.
principalmente por bactérias, além de serem predominantes em ambientes extremos como fontes termais (ROBERTSON et al., 2005).
Apesar de ainda não terem sido encontrados indícios de arqueias patogênicas em plantas, vários trabalhos associam arqueias com doenças em humanos.
Arqueias já foram encontradas no intestino, vagina e gengivas humanas. No estudo das fezes de indivíduos, Methanobrevibacter smithii foi o microrganismo metanogênico mais abundante encontrado (MILLER; WOLIN, 1983). Na vagina, as amostras positivas para arqueias do gênero Methanobrevibacter foram obtidas de pacientes com vaginose bacteriana, um desequilíbrio da microbiota responsável por corrimento vaginal (BELAY et al., 1990). Diversos estudos associaram arqueias metanogênicas com periodontite, uma infecção anaeróbica causada por complexos microbianos: as chamadas placas dentais (LEPP et al., 2004; VIANNA et al., 2008). Methanobrevibacter oralis, uma arqueia pertencente ao filo Euryarchaeota, foi identificada em placas dentais e sua abundância foi relacionada com o grau de severidade da periodontite em humanos (BRINGUIER et al., 2013). Membros deste gênero (M. smithii e M. oralis) também foram identificados em gengivas humanas e de macacos
(Macaca mulatta) (KEMP et al., 1983; KULIK et al., 2001). Pérez-Chaparro e colaboradores,
em um estudo de revisão atual, analisaram dados de 41 estudos sobre periodontite, mostrando que o domínio Archaea, assim como diversas espécies bacterianas, estão envolvidos com essa doença (PEREZ-CHAPARRO et al., 2014). Em um estudo comparativo das microbiotas de raspados de língua de indivíduos sadios e com periodontite, abundâncias moderadas de arqueias foram encontradas em indivíduos sadios, enquanto que um aumento na abundância foi encontrado em indivíduos doentes (GOEHLER et al., 2014).
Essa relação entre o aumento da abundância de arqueias na microbiota bucal e incidência de doenças periodontais foi observada em inúmeros trabalhos. Existem indícios de que arqueias metanogênicas que habitam o intestino grosso de indivíduos com doenças severas de cólon poderiam contribuir para as doenças a partir de produção de metano, produzindo um ambiente adequado para o crescimento de microrganismos patogênicos, em um papel indireto no surgimento dessas doenças (DE MACARIO; MACARIO, 2009).
Todavia, recentemente, foi observada a indução de resposta inflamatória pela arqueia metanogênica M. stadtmanae em intestino humano, um importante fator indicativo de patogenicidade (BANG et al., 2014).
relações simbióticas conhecidas envolvendo arqueias: a interação entre Ignicoccushospitalis, hospedeira da Nanoarchaeum equitans, uma nanoarqueia até o presente momento nunca cultivada sozinha (FORTERRE, GRIBALDO; BROCHIER-ARMANET, 2009). Também já foram registrados casos de simbiose entre arqueias e bactérias, como no caso da interação entre arqueias oxidadoras de metano e bactérias redutoras de sulfato (NAUHAUS et al., 2007).
Arqueias também são endossimbiontes de eucariontes, como no caso de ciliados marinhos vivendo em regiões anóxicas (EDGCOMB et al., 2011). Além disso, arqueias metanogênicas são encontradas no intestino de ruminantes (revisado por (JANSSEN; KIRS, 2008)).
O último fator a ser analisado em relação a uma possível patogenicidade de arqueias é a presença de assinaturas moleculares comuns a patógenos, como a produção de toxinas. As arqueias halófilas são conhecidas pela produção de toxinas, as halocinas, proteínas de tamanhos diversos capazes de inibir o crescimento de outras arqueias halófilas (O'CONNOR; SHAND, 2002). No entanto, ainda não foi encontrada toxicidade contra membros dos demais domínios (HASELTINE et al., 2001).
3. OBJETIVOS
3.1. Objetivos Gerais
Estudo dos impactos da implementação de uma monocultura (dendê) sobre a microbiota de arqueias de solo de mata nativa amazônica desmatada pra esse fim; além do estudo do impacto do Amarelecimento Fatal sobre as comunidades de arqueias de solos associados à dendezeiros cultivados em solos amazônicos.
3.2. Objetivos Específicos
• Caracterização e comparação das comunidades de arqueias do solo de mata nativa e cultivados com dendezeiros por meio da tecnologia do pirosequenciamento do gene que codifica o 16S rRNA.
4. MATERIAIS E MÉTODOS
4.1. Coleta de Solo
A cidade de Moju, no Pará, está localizada a 117 km de Belém, é coberta por árvores de 25 a 35 m de altura em uma floresta tropical densa (RIBEIRO, SILVA; CARVALHO, 2003). O clima é equatorial, quente e úmido (Am), de acordo com a classificação de Köppen-Geiger, com temperaturas anuais variando de 25 a 27oC e precipitações entre 2.000 mm e 3.000 mm por ano, distribuídas irregularmente (COSTA et al., 1998). O solo é predominantemente latossolo amarelo (oxisol) (ALMEIDA et al., 2001).
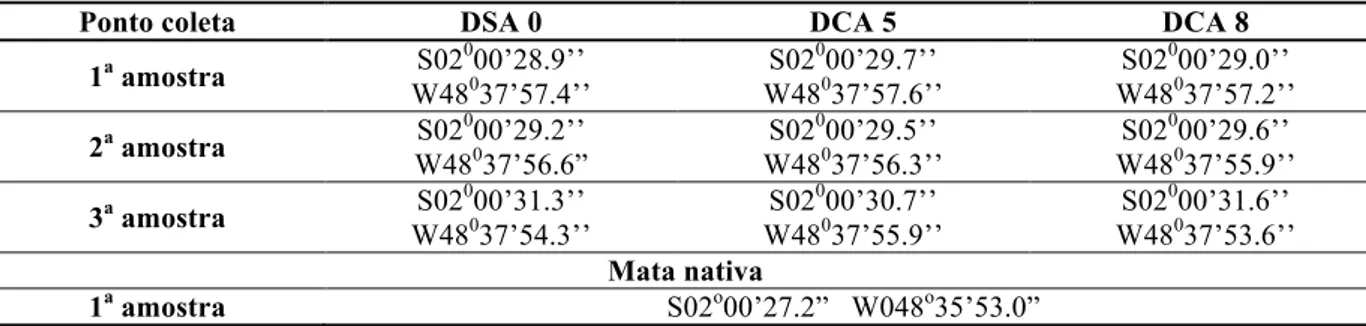
As amostras de solos de dendezeiros foram coletadas em outubro de 2010 na época chuvosa na Agroindústria Marborges, em Moju, Pará, e a amostra de solo de mata nativa em uma região de floresta amazônica adjacente às áreas cultivadas (BERGMANN, 2013). A coleta foi realizada na Empresa Marborges em época de chuvas; no local ocorrem alagamentos, sendo muitas vezes necessárias drenagens dos solos nas áreas de plantio (Tabela 2).
Tabela 2 – Pontos de coleta na Agroindústria Marborges georreferenciados.
Ponto coleta DSA 0 DCA 5 DCA 8
1a amostra S02
000’28.9’’
W48037’57.4’’
S02000’29.7’’ W48037’57.6’’
S02000’29.0’’ W48037’57.2’’
2a amostra S02
0
00’29.2’’ W48037’56.6”
S02000’29.5’’ W48037’56.3’’
S02000’29.6’’ W48037’55.9’’
3a amostra S02
000’31.3’’
W48037’54.3’’
S02000’30.7’’ W48037’55.9’’
S02000’31.6’’ W48037’53.6’’ Mata nativa
1a amostra S02o00’27.2” W048o35’53.0” DSA: dendezeiros sem AF; DCA: dendezeiros com AF.
nomear as amostras que leva em consideração três fatores: o estágio da doença (DCA – dendezeiros com AF: 5 ou 8 e DSA – dendezeiros sem AF: 0), o grupo de palmeiras escolhido (1, 2 ou 3) e o lado da planta em que foi feita a coleta (1 ou 2) (Figura 5). As amostras foram armazenadas em sacos plásticos em gelo seco até serem armazenadas em freezer -80oC até o momento das análises.
Figura 5 - Desenho esquemático da amostragem dos solos. Cada grupo contém três dendezeiros em três estágios da doença. Os pontos 1 e 2 representam o lado da planta onde ocorreu a coleta.
4.2. Análises Físico-Químicas do Solo
4.3. Extração do DNA Total do Solo
O DNA total das amostras de solo de dendezeiros foi extraído anteriormente (BERGMANN, 2013), de acordo com o protocolo de Smalla e colaboradores (1993), com modificações.
O DNA total do solo de mata nativa foi extraído utilizando-se o mesmo protocolo. Numa tentativa de diminuir o viés relativo à extração de DNA total do solo e extrair o DNA total dos microrganismos, inclusive dos raros, foram feitas 4 extrações por amostra composta de solo.
temperatura ambiente. Os precipitados foram secos ao ar e ressuspensos em 50 µL de tampão TE 1x (1 mM de Tris e 1 mM de EDTA em pH 8,0). Um gel de agarose 0,8% corado com brometo de etídio (2 µg/mL) e exposição à luz UV (312 nm) foi usado para verificação da qualidade das extrações.
4.4. PCR do gene 16S rRNA e Pirosequenciamento
Uma grande limitação na caracterização de comunidades microbianas ambientais usando métodos independentes de cultura é a escolha dos primers, especialmente para
Archaea. A maioria dos primers específicos para Archaea foram desenhados quando os
bancos de dados eram limitados e não cobriam a diversidade existente de arqueias (BAKER, SMITH; COWAN, 2003). Os primers escolhidos para serem usados neste trabalho, 340f/1000r (GANTNER et al., 2011), foram recentemente desenhados baseando-se no alinhamento de 8.500 sequências de 16S rRNA do banco de dados SILVA (QUAST et al., 2013). Devido ao grande número de sequências de arqueias utilizadas, além do fato de serem utilizadas Archaea de diversos ambientes, é esperado que estes primers sejam capazes de amplificar um grande diversidade de arqueias.
As 10 amostras escolhidas foram utilizadas para realização de PCR usando os primers 340f (5’ – CCCTAYGGGGYGCACAG – 3’) e 1000r (5’ – GGCCATGCACYWCYTCTC – 3’). Foram ainda utilizados adaptadores A e B para pirosequenciamento em 454 e 10 diferentes MIDs (Multiplex Identifiers), já que as 10 amostras formaram um pool que foi sequenciado em uma só placa, sendo possível a identificação posterior das sequências de cada uma das 10 amostras (ROCHE, 2011). Os MIDs utilizados para identificar as 10 amostras e o agrupamento das mesmas para as análises posteriores se encontram na Tabela 12 – Anexo D.
Estão esquematicamente representadas a seguir as estruturas dos primers direto e reverso: Primer direto 340f:
5’-CCATCTCATCCCTGCGTGTCTCCGACTCAG-[MID]-[340f]-[template]-3’
Primer reverso 1000r:
5’-CCTATCCCCTGTGTGCCTTGGCAGTCTCAG-[CGAGAGATAC]-[1000r