Nutritive value of Tanzania grass for dairy cows under rotational grazing
Texto
Imagem
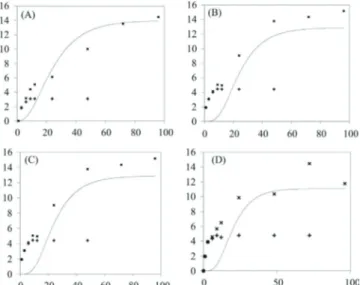
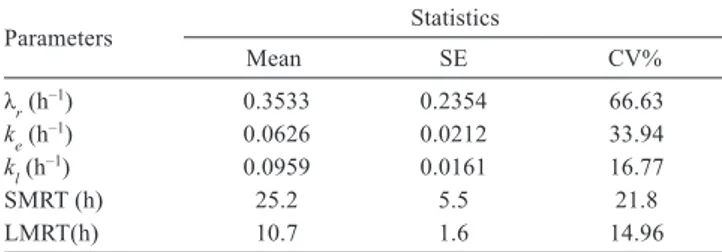
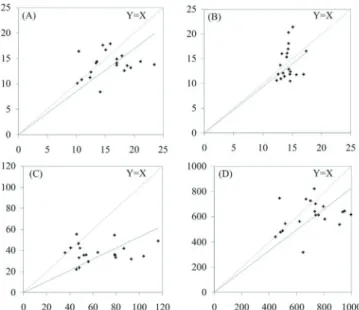
Documentos relacionados
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
While static studies suggested that nails/claws could not disrupt bone, specific tiger enrichment activities documented that bones were susceptible to damage from the kinetic
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
didático e resolva as listas de exercícios (disponíveis no Classroom) referentes às obras de Carlos Drummond de Andrade, João Guimarães Rosa, Machado de Assis,
Este modelo permite avaliar a empresa como um todo e não apenas o seu valor para o acionista, considerando todos os cash flows gerados pela empresa e procedendo ao seu desconto,
i) A condutividade da matriz vítrea diminui com o aumento do tempo de tratamento térmico (Fig.. 241 pequena quantidade de cristais existentes na amostra já provoca um efeito
Na hepatite B, as enzimas hepáticas têm valores menores tanto para quem toma quanto para os que não tomam café comparados ao vírus C, porém os dados foram estatisticamente
Para uma melhor abordagem do recorte histórico proposto será necessário contextualizar as condições de possibilidade da inserção da irmandade AA na cidade, partindo
