THE CEREBRAL CORRELATES OF SET-SHIFTING
An fM RI st udy of t he t rail making t est
Jorge M oll
1, Ricardo de Oliveira-Souza
1,2, Fernanda Tovar M oll
1,
Ivanei Edson Bramat i
1, Pedro Angelo Andreiuolo
1ABSTRACT - The t rail making t est (TM T) pert ains t o a family of t est s t hat t ap t he abilit y t o alt ernat e bet w een cognit ive cat egories. How ever, t he value of t he TM T as a localizing inst rument remains elusive. Here w e report t he result s of a funct ional magnet ic resonance imaging (fM RI) st udy of a verbal adapt at ion of t he TM T (vTM T). The vTM T t akes advant age of t he set -shift ing propert ies of t he TM T and, at t he same t ime, minimizes t he visuospat ial and visuomot or component s of t he w rit t en TM T. Whole brain BOLD fM RI w as performed during t he alt ernat ing execut ion of vTM TA and vTM TB in seven normal adult s w it h more t han 12 years of formal educat ion. Brain act ivat ion relat ed t o t he set -shift ing component of vTM TB w as invest igat ed by comparing performance on vTM TB w it h vTM TA, a simple count ing t ask. There w as a marked asymmet ry of act ivat ion in favor of t he left hemisphere, most not ably in dorsolat eral prefront al cort ex (BA 6 lat eral, 44 and 46) and supplement ary mot or area/cingulat e sulcus (BA 6 medial and 32). The int rapariet al sulcus (BA 7 and 39) w as bilat erally act ivat ed. These findings are in line w it h clinico-anat omic and funct ional neuroimaging dat a t hat point t o a crit ical role of t he dorsolat eral and medial prefront al cort ices as w ell as t he int rapariet al sulci in t he regulation of cognitive flexibility, intention, and the covert execution of saccades/anti-saccades. M any commonly used neuropsychological paradigms, such as t he St roop, Wisconsin Card Sort ing, and go - no go t asks, share some pat t erns of cerebral act ivat ion w it h t he TM T.
KEY WORDS: t rail making, funct ional M RI, execut ive funct ion, prefront al cort ex, int rapariet al sulcus, cognit ive sw it ching.
Correlatos cerebrais da flexibilidade cognitiva: RM funcional do teste das trilhas
RESUM O - O Test e de t rilhas (TT) pert ence a uma família de t est es que aferem a capacidade de alt ernar ent re cat egorias cognit ivas. Não obst ant e, o valor de localização do TT permanece obscuro. Nest e est udo, relat amos a análise de um est udo de uma adapt ação verbal do TT (TTv) com ressonância magnét ica funcional (RM f). O TTv realça a propriedade de mudança de cat egorias do TT convencional (escrit o) ao mesmo t empo em que minimiza seus component es visuoespaciais e visuomot ores. O cérebro int eiro foi est udado pela t écnica BOLD durant e a execução alt ernant e do TTvA e TTvB em set e adult os normais com mais de 12 anos de escolaridade. A ativação cerebral relacionada ao componente de alternância cognitiva da Parte B foi investigada comparando-se o decomparando-sempenho no TTvB e no TTvA. Obcomparando-servamos acent uada assimet ria de at ivação a favor do hemisfério esquerdo, especificament e no córt ex front al dorsolat eral (áreas 6 lat eral, 44 e 46) e área mot ora suplement ar/ sulco do cíngulo (área 6 medial e 32). O sulco int rapariet al (áreas 7 e 39) foi at ivado bilat eralment e. Esses achados est ão de acordo com dados clínico-anat ômicos e de imagem funcional que apont am para o papel crítico do córtex prefrontal dorsolateral e medial, bem como do sulco intraparietal, na regulação da flexibilidade cognit iva, dos mecanismos de int enção e execução de saques e ant i-saques oculares. M uit os paradigmas neuropsicológicos de amplo emprego na prát ica clínica, como os t est es de St roop e de Wisconsin, e t arefas do t ipo vai / não vai, compart ilham padrões de at ivação cerebral com o TT.
PALAVRAS-CHAVE: t est e de t rilhas , ressonância magnét ica funcional, córt ex prefront al, sulco int rapariet al, função execut iva, alt ernância cognit iva.
St udy conduct ed at t he Barra D’Or Hospit al, RJ Rio de Janeiro, Brazil: 1LABS–Rede D’Or, Grupo de Neuroimagem e Neurologia do Comport ament o (GNNC), RJ; 2Hospit al Universit ário Gaffrée e Guinle e Inst it ut o Philippe Pihel.
Received 12 April 2002. Accept ed 24 June 2002.
Dr. Jorge M oll - Avenida Ayrt on Senna 2541 - 22793-010 Rio de Janeiro, RJ - Brasil. E-mail: [email protected] The abilit y t o sw it ch bet w een cognit ive cat
ego-ries, or “ sets” , is a classical indicator of normal neuro-behavioral funct ioning1. The t rail marking t est (TM T)
is w idely employed as a diagnost ic t ool for elicit ing
shifts betw een cognitive sets2. It consists of tw o parts
paper3. In Part A (TM TA) t he circles are numbered
from 1 t o 25, w hereas in Part B (TM TB) t hey cont ain numbers from 1 t o 13 and let t ers from A t o M (t he Port uguese alphabet does not include t he let t er K), w hich must be connect ed in alt ernat ing progression, from 1-A t o M -13. Tot al score is t he t ime in seconds spent t o complet e each part . In view of t he low lo-calizing and lat eralizing value of t he TM T4,5, t he
ce-rebral areas t hat are crit ical for it s execut ion have been difficult t o define on t he basis of clinico-ana-t omic daclinico-ana-t a alone6. A verbal adapt at ion of t he TM T
(vTM T) has been increasingly employed in normal individuals as w ell as in mixed clinical samples7,8. The
vTM T is more suit ed for probing t he abilit y t o sw it ch bet w een cognit ive set s because it reduces t o a mini-mum t he visuospat ial and visuomot or fact ors of t he w rit t en format9.
In t he present st udy w e used funct ional magnet ic resonance imaging (fM RI) t o invest igat e t he cerebral correlat es of t he vTM T, w hile at t empt ing t o preserve t he basic cognit ive st ruct ure of vTM T as a neuropsy-chologic inst rument .
M ETHOD
Neuropsychological paradigm
Seven right-handed normal volunteers (three men) w ith ages ranging from 19 t o 43 years (mean = 24± 9 years) part icipat ed in t he st udy aft er providing w rit t en informed consent . Subject s had m ore t han 12 years of f orm al educat ion and a mean Edinburgh Invent ory score10 of 82 ± 3011. Bef ore scanning, subject s w ere administ ered t he w rit t en version of t he TM T and rehearsed t he verbal adap-t aadap-t ion of adap-t he adap-t esadap-t . To maadap-t ch adap-t he scanning procedure as closely as possible, a few t rials of 25 seconds each w ere administ ered during t he t raining session. Subject s w ere asked t o st art count ing covert ly at 1 (vTM TA) and t o alt er-nat e bet w een numbers and let t ers (vTM TB) as quickly as possible w hen t hey heard t he commands “ count ” and “ alt ernat e” , respect ively. To prevent unsolvable anagram effect s, subject s w ere t old t o st art over if t hey slipped off t he t rack. During scanning, t he procedure w as repeat ed. Besides, t hey w ere asked t o keep t heir eyes shut and t o avoid making art iculat ory movement s. Af t er t he scanning session, subject s rat ed t he at t ent ional effort allocat ed t o each part of t he t est on 100 mm visual analogue scales (VAS) and report ed how far t hey had progressed, on ave-rage, on each part of t he t est12. Tot al scores for vTM TA and vTM TB w ere, respect ively, t he highest number and let t er reached in t heir best t rials. For t he purposes of dat a analysis, t he numerical order of t hat let t er in t he alphabet w as doubled and used as t he final vTM TB score.
St at ist ical analysis of cognit ive rat ings
We obt ained six cognit ive measures from each part ici-pant , namely, TM T (A and B), vTM T (A and B), and t he VAS
for at t ent ional effort for each part of vTM T. Performance indexes on Part s A and B of each t ask w ere compared w it h St udent ’s t-t est for paired samples, t w o-t ailed11. To know w het her any possible differences bet w een vTM T A and B w ould survive t he confounding effect s of at t en-t ional efforen-t and en-t he explicien-t visuospaen-t ial/visuomoen-t or fac-t ors of fac-t he w rifac-t fac-t en formafac-t , w e compared vTM T A and B performances w it h analysis of covariance ent ering TM T A and B, and t he VAS scores as covariat es. M eans differences w ere assessed posthoc w it h Scheffé’s t est . A 0.05 level of significance, t w o-t ailed, w as set for all st at ist ical t est s.
f M RI procedures
Functional data covering the w hole brain w ere acquired on a Siemens Vision 1.5T scanner. One hundred funct ional volumes w ere collect ed during t he alt ernat ion of vTM TA and vTM TB in a blocked design, each block last ing 25 se-conds. There w ere 10 blocks for part A and t en blocks for part B of t he t est . Sequence paramet ers w ere: gradient -echo EPI, TE = 66 ms, TR = 5 s, flip angle = 90°, s lice t hickness 5.0 mm, int er-slice gap 0.25 mm, FOV = 250 mm, 128x128 mat rix. Dat a w ere analyzed using BrainVo-yager v. 3.9 (Brain Innovat ion, Germany). Funct ional dat a w ere mot ion correct ed in t hree-dimensional space, and spat ial (4mm FWHM ) and t emporal (3-32Hz) smoot hing w ere performed in the frequency domain. Intersubject ana-lysis w as performed using t he general linear model ap-proach. Act ivat ion maps w ere creat ed using cross-corre-lat ion analysis, t ransformed int o t he Talairach space, and coregist ered t o t he anat omic M RI. P-values < 0.05, cor-rect ed for mult iple comparisons, w it h a 3D clust er t hre-shold of 200 voxels in volumet ric int erpolat ed dat a w ere considered significant13. Lat eralizat ion indexes for regions of int erest (ROI) w ere comput ed follow ing an adapt at ion of Oldfield’s formula for handedness10, w hich t ook int o account t he number of voxels (NOV) per ROI:
NOV Right ROI – NOV Left ROI x 100 NOV Right ROI + NOV Left ROI
RESULTS
Table 1. Neuropsychological result s (mean ± sd).
TM T (seconds)* VTM T* * At t ent ional effort * * *
(it ems per block of 25 seconds) (visual analogue scales, mm)
A B A B A B
21 ± 5 45 ± 23 60 ± 14 29 ± 10 17 ± 17 57 ± 18
*t = 3.26, p < 0.02; * * t = 6.7, p < 0.002; * * * t = 5.3, p < 0.004.
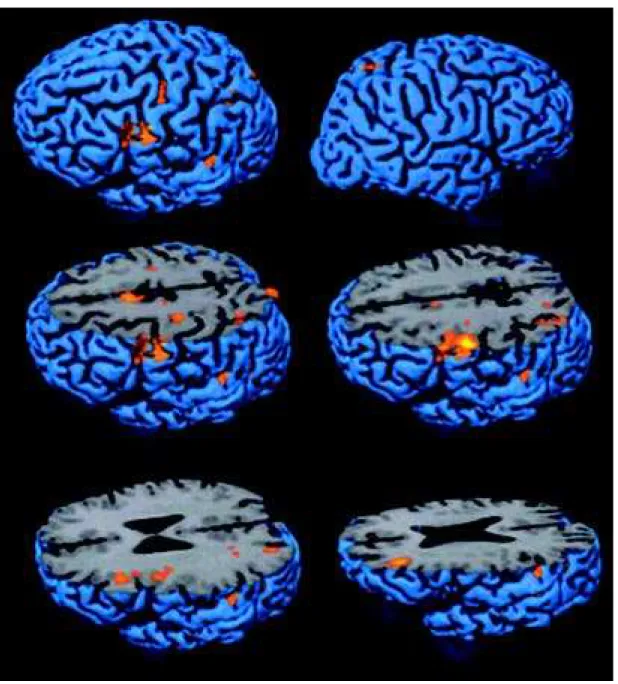
Table 2. Hemispheric side and regions act ivat ed by vTM TB minus vTM TA.
Talairach
Side Put at ive coordinat es
Brodmann Area x y z
low er precent ral gyrus L 6 lat eral -44 -2 38
inferior front al sulcus L 44, 46 ¯ 40 23 29
middle front al gyrus L 44, 46 -36 38 22
dorsal premot or cort ex L 6 lat eral -31 -16 50
int rapariet al sulcus L 7, 39 -35 -55 34
rost ral SM A and cingulat e sulcus L 6 medial, 32 -06 3 49
int rapariet al sulcus R 7, 39 26 -55 34
cut s on Figure 2 show s t hat most act ivat ions fall w it hin cort ical sulci.
DISCUSSION
The TM T pert ains t o a family of neuropsycholo-gical t est s w hich t ap t he abilit y t o inhibit prepot ent responses in favor of less habitual, yet more adaptive, ones. Ot her members of t he family include t he go-no go and t he St roop t asks14. The TM T differs from
bot h, crit ically, because it requires verbal t hinking t o follow a predet ermined concept ual rout e, w hich is given by t he ordered series of numbers (part A), or numbers and let t ers (part B). Not w it hst anding t his fact, the similarities among these tasks may outw eigh t he differences. Behaviorally, all seem t o share a com-mon ground of cognit ive int erference, response con-flict , and cognit ive set shift ing. M oreover, funct ional neuroim aging st udies have show n t hat t he areas act ivat ed by bot h t he St roop and t he go-no go t asks overlap t o a great ext ent , most not ably in t he middle dorsolat eral and medial prefront al cort ex as w ell as in t he post erior pariet al cort ex15,16. This pat t ern is
remarkably similar t o t he one w e found for t he TM T.
The ant erior hemispheric asymmet ry seen in our subject s concurs w it h evidence implicat ing t he left hem isphere in sym bolic t asks. It m ay also ref lect som e st ruct ural peculiarit ies of vTM T proper. In
support of t his view, t he concept ual handling of numbers w as associat ed w it h act ivat ion of t he left inferior front al gyrus and sulcus, premot or cort ex, and inferior pariet al lobule17. The left inferior
pre-front al cort ex w as likew ise found t o be more act ive in let t er t han in semant ic fluency t asks18. The left
pref ront al sect ors act ivat ed in our st udy largely overlap w it h t he inf erior pref ront al area of ot her investigations w hich implicate the inferior frontal gyri and sulci in t he mediat ion of cognit ive sw it ching. Thus, w hile t he right prefront al cort ex seems t o be crit ical for response suppression in go-no go t asks, t he left is more direct ly involved in biasing responses t ow ards novel principles at play15,16,19,20. The role of
the inferior dorsolateral prefrontal cortex in providing flexibilit y t o behavior is st rengt hened by t he obser-vat ion t hat cognit ive inflexibilit y, or “ perseverat ion” , is a core sympt om of damage t o t hese areas21. The
left premot or cort ex act ivat ion seen in our subject s may be relat ed t o t he t ask sw it ching demands of vTM TB. This area w as also act ivat ed in a st udy of t he neural correlat es of t ool-use w hich required rapid manual shift s of t ool-relat ed act ions22. The left
of the left hemisphere is critically involved in tasks that depend on rapid action and cognitive shifts.
An alternative, or complementary, explanation for t he inferior dorsolat eral act ivat ion involving w orking memory mechanisms might be raised. That t his w as not t he case, how ever, is suggest ed by t he fact t hat bot h vTM T A and B required comparable inst ruct ions (i.e. “ count ” and “ alt ernat e” ), t hus equalizing t he w orking memory load across blocks. This conclusion is support ed by t he absence of act ivat ion in BA 9, w hich is relat ed t o t he holding of inst ruct ions in w or-king memory during t ask execut ion23.
The cont ribut ion of t he medial front al cort ex t o normal TM TB performance had already been
sug-gest ed by case report s of pat ient s w it h damage t o this region24 or to its subcortical connections25,26.
The-se individuals easily loThe-se t rack of t he alt ernat ing res-ponse pattern. The supracingular cluster of activation in our subject s w as locat ed in an area slight ly post e-rior t o t he plane of t he ant ee-rior commissure, corres-ponding t o t he supplement ary mot or area27. This
area exert s a f acilit at ory inf luence on m ovem ent preparat ion and init iat ion or, more simply, on “ in-t enin-t ion”28. The fact t hat bilat eral damage t o t he
me-dial front al lobe leads t o loss of spont aneit y and be-havioral slow ness29, w hile unilat eral damage gives
rise t o t he clinical syndrome of “ mot or neglect ”30
dial act ivat ion observed in t he present st udy reflect s t he call int o play of processes w hich lie at t he int en-t ion-acen-t ion inen-t erface. The absence of anen-t erior cingu-lat e act ivat ion is in agreement w it h recent st udies show ing t hat cognit ive int erference is primarily me-diat ed by t he cort ex of t he left inferior front al sulcus, w hile t he ant erior cingulat e is more direct ly relat ed t o response preparat ion28,31. Since our st udy prot ocol
reduced overt mot or responses t o a minimum, it is not surprising t hat t he ant erior cingulat e w as not significant ly act ivat ed in our subject s.
The cort ex w it hin and around t he IPS is a complex somat osensory and visual convergence zone t hat has been funct ionally relat ed t o object and spat ial w or-king memory, visual search, tool-use praxis, reaching, and reallocat ion of at t ent ional resources22,32.
Ho-w ever, only a feHo-w st udies have addressed t he me-aning of IPS act ivat ion in paradigms employing cog-nit ive sw it ching31. A pat t ern of bilat eral IPS act
iva-t ion similar iva-t o iva-t he one found in iva-t he preseniva-t invesiva-t i-gat ion has been observed in associat ion w it h t he
generat ion of ant i-saccades, covert visual orient ing and suppression of oculomot or responses33. Alt
ho-ugh w e did not monit or t he eye movement s of our subject s, t he possibilit y t hat bilat eral IPS act ivat ion during vTM TB performance is relat ed t o t he covert int erplay of saccades and ant i-saccades w hile t he individual ment ally t racks his progression along t he concept ual let t er-number t rail deserves furt her expe-riment al t est ing34.
The observat ion t hat act ivat ions t ended t o fall w it hin cort ical sulci adds t o t he evidence in favor of t he “ fundal cognit ion hypot hesis”35. According t o
t his hypot hesis, t he neural comput at ions underlying complex cognit ive operat ions t ake place preferen-t ially w ipreferen-t hin corpreferen-t ical sulci, in conpreferen-t raspreferen-t preferen-t o preferen-t he gyral surfaces. The fundal cognit ion hypot hesis is in kee-ping w it h t he complexit y of t he operat ions involved in t rail making.
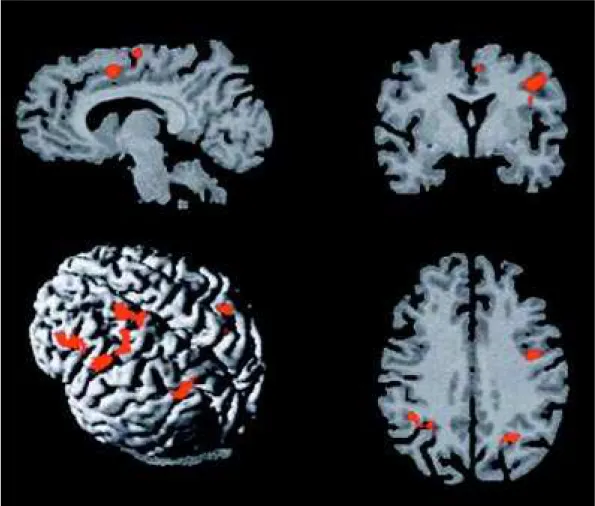
St udies as t he present one m ay provide new insights into the brain circuits underlying the execution Fig 2. A: M edial w all of t he lef t f ront al lobe show ing act ivat ion in t he cingulat e sulcus (CS) and supplement ary
of com m on neuropsychological t est s by norm al individuals and shed light on t he m echanism s of impairment and recovery of function in patients w ith brain disease. They m ay also im prove neuropsy-chological test design by refining cognitive dimensions of interest on the basis of brain activation patterns.
Acknowledgements - The authors are indebted to Prof. Omar da Rosa Santos, Hospital Universitário Gaffrée e Guinle.
REFERENCES
1. Freeman T, Gathercole CE. Perseveration-the clinical symptoms-in chro nic schizo p hrenia and o rganic d ementia. Bri J Psychiatry 1966;112:27-32.
2. Butler M, Retzlaff P, Vanderploeg R. Neuropsychological test usage. Professional Psychology: Research and Practice 1991;22:510-512. 3. Reitan RM. Validity of the Trail Making Test as an indicator of organic
brain damage. Perceptual and Motor Skills 1958;8:271-276. 4. Lezak MD. Neuropsychological assessment. 3.Ed. New York: Oxford
Univ Press, 1995:381-384.
5. Reitan RM, Tarshes EL. Differential effects of lateralized brain lesions on the Trail Making Test. J Nervous Mental Dis 1959;129:257-262. 6. Spreen O, Strauss E. A Compendium of neuropsychological tests:
administration, norms, and commentary. 2.Ed. New York: Oxford Univ Press, 1998.
7. A braham E, A xelrod BN, Ricker JH. A pplication of the Oral Trail Making Test to a mixed clinical sample. Archi Clini Neuropsychol 1996;11:697-701.
8. Oliveira-Souza R, Moll J, Passman LJ, et al. Trail making and cognitive set-shifting. Arq Neuropsiquiatr 2000;58:826-829.
9. Crowe SF. The differential contribution of mental tracking, cognitive flexibility, visual search, and motor speed to performance on Parts A and B of the Trail Making Test. J Clin Psychol 1998;54:585-591.
10. Old field RC. The assessment and analysis o f hand ed ness: The Edinburgh Inventory. Neuropsychologia 1971;9:97-113.
11. Kachigan SK. Statistical analysis. an interdisciplinary introduction to univariate & multivariate methods. New York: Radius Press, 1986:589. 12. McCormack HM, Horne DJ, Sheather S. Clinical applications of visual
analogue scales: a critical review. Psychol Med 1988;18:1007-1019. 13. Forman SD, Cohen JD, Fitzgerald M, Eddy WF, Mintun MA, Noll DC.
Improved assessment of significant activation in functional magnetic resonance imaging (fMRI): use of a cluster size threshold. Magnetic Resonance Med 1995;33:636-647.
14. Weintraub S. Neuropsychological assessment of mental state. In Mesulan MM (Ed.) Principles of behavioral and cognitive neurology. 2.Ed. New York: Oxford Univ Press, 2000:121-173.
15. Garavan H, Ross TJ, Stein EA . Right hemispheric dominance for inhibitory control: an event-related functional MRI study. Proc Natl Acad Sci USA 1999;96:8301-8306.
16. Kawashima R, Satoh K, Itoh H, et al. Functional anatomy of GO/ NO-GO discrimination and response selection: a PET study in man. Brain Res 1996;728:79-89.
17. Rickard TC, Romero SG, Basso G, Wharton C, Flitman S, Grafman J. The calculating brain: an fMRI study. Neuropsychologia 2000;38:325-335. 18. Paulesu E, Goldacre B, Scifo P, et al. Functional heterogeneity of left inferior
frontal cortex as revealed by fMRI. NeuroReport 1997;8:2011-2016. 19. Do ve A , Po llmann S, Schubert T, Wiggins CJ, vo n Cramo n DY.
Prefrontal cortex activation in task switching: an event-related fMRI study. Cogn Brain Res 2000;9:103-109.
20. Konishi S, Kawazu M, Uchida I, Kikyo H, Asakura I, Miyashita Y. Contribution of w orking memory to transient activation in human inferior prefrontal cortex during performance of the Wisconsin Card Sorting Test. Cer Cortex 1999;9:745-753.
21. Stuss DT, Levine B, Alexander MP, et al. Wisconsin Card Sorting Test performance in patients with focal frontal and posterior brain damage: effects of lesion location and test structure on separable cognitive processes. Neuropsychologia 2000;38:388-402.
22. Moll J, Oliveira-Souza R, Passman LJ, Cunha FC, Lima FS, Andreiuolo PA. Functional magnetic resonance imaging of pantomimes of tool use. Neurology 2000;54:1331-1336.
23. MacDonald AW, Cohen JD, Stenger VA, Carter CS. Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Science 2000;288:1835-1838.
24. Leimkuhler ME, Mesulam MM. Reversible go-no go deficits in a case frontal lobe tumor. Ann of Neurol 1985;18:617-619.
25. Sandson TA, Daffner KR, Carvalho PA, Mesulam MM. Frontal lobe dysfunction following infarction of the left-sided medial thalamus. Arch of Neurol 1991;48:1300-1303.
26. Jahanshahi M, Ardouin CMA, Brown RG, et al. The impact of deep brain stimulation on executive function in Parkinson’s disease. Brain 2000;123;1142-1154.
27. Humbersto ne M, Saw le GV, Clare S, et al. Functio nal magnetic resonance imaging of single motor events reveals human presupple-mentary motor area. Ann Neurol 1997;42:632-637.
28. Heilman KM, Valenstein E, Watson RT. Intentional motor disorders. In Levin HS, Eisenberg HM, Benton AL (Eds.). Frontal lobe function and dysfunction. New York: Oxford Univ Press, 1991:199-213. 29. Fisher CM. A bulia mino r vs agitated behavio r. Clin Neuro surg
1984;31:9-31.
30. Laplane D, Degos JD. Motor neglect. J Neurol Neurosurg Psychiatry 1983;46:152-158.
31. Zysset S, Müller K, Lohmann G, von Cramon DY. Color-word matching Stroop task: separating interference and response conflict. Neuroimage 2001;13:29-36.
32. Belger A, Puce A, Krystal JH, Gore JC, Goldman-Rakic P, McCarthy G. Dissociation of mnemonic and perceptual processes during spatial and nonspatial working memory using fMRI. Hum Brain Mapp 1998;6:14-32. 33. Co nno lly JD, Go o d ale MA , Deso uza JFX, Meno n RS, Vilis T. A
comparison of frontoparietal fMRI activation during anti-saccades and anti-pointing. J Neurophysiol 2000;84:1645-1655.