Transcriptome sequencing reveals differences between primary and secondary hair follicle-derived dermal papilla cells of the Cashmere goat (Capra hircus).
Texto
Imagem
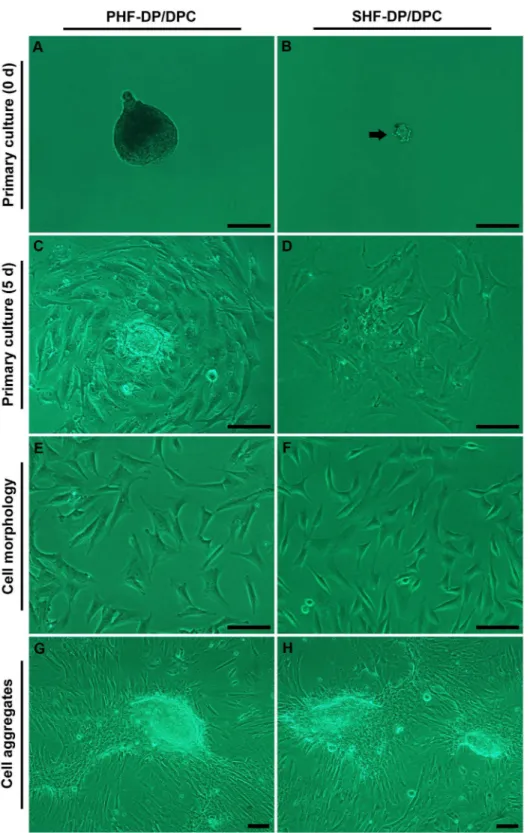
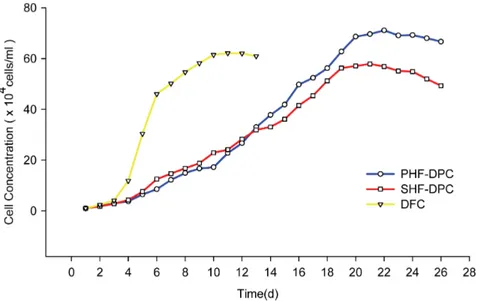
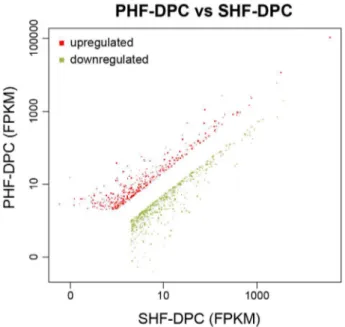
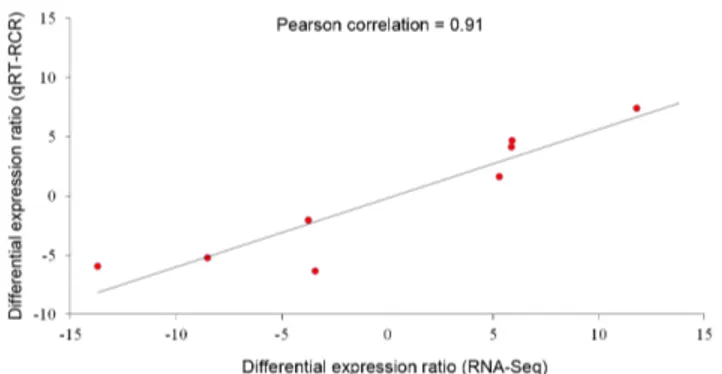
Documentos relacionados
After the two burns, the comparative statistical ana1ysis showed that there were no differences between the three groups regarding induction of vascularization (p
In the present study, the results of the analyzed behavioral tests were considered reliable as no differences were found in the comparison between the test and retest
The differences between the predicted spirometry values derived for White adults in Brazil in 2007 and those derived for Black adults in the present study were plotted against
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
the sialyltransferase-transduced cells revealed that groups of genes involved in specific functions were regulated in parallel in the two cell lines, regardless
Significant differences were also observed between diploid and triploid for frequencies of differential cell counts, suggesting that triploidy affected the number of cells as well
While no differences in the tumor cells were detected between the mice from the Ehrlich cell (E) group and those from the isolated treatment (CY and MI) groups,
The integration of functional gene annotations in the analysis confirms the detected differences in gene expression across tissues and confirms the expression of porcine genes being
