Bulletin of Insectology 69 (1): 117-126, 2016
ISSN 1721-8861
Distribution and genetic variability of Staphylinidae across a
gradient of anthropogenically influenced insular landscapes
José A. P. MARCELINO1,2, Rosanna GIORDANO3, Paulo A. V. BORGES1, Patricia V. GARCIA1,2, Felipe N.
SOTO-ADAMES3,4, António O. SOARES1,2
1CE
3C, Centre for Ecology, Evolution and Environmental Changes, Azorean Biodiversity Group, Department of
Biology, University of the Azores, Portugal
2CIRN, Department of Biology, University of the Azores, Portugal 3Department of Biology, University of Puerto Rico, San Juan, USA 4Department of Entomology, University of Illinois, Urbana, USA
Abstract
This paper describes the distribution and genetic variability of rove beetles (Coleoptera Staphylinidae) in anthropogenically influ-enced insular landscapes. The study was conducted in the Azores archipelago, characterized by high anthropogenic influence and landscape fragmentation. Collections were made in five islands, from eight habitats, along a gradient of anthropogenic influence. The species of Staphylinidae from the Azores collected for this study were widely distributed and showed low habitat fidelity. Rove beetle richness was associated with anthropogenic influence and habitat type, increasing from less to more anthropogenic impacted habitats. However, genetic diversity of profiled species (i.e. with three or more specimens per species/habitat) does not seem affected by anthropogenic influence in the different habitat types, isolation or landscape fragmentation. COI haplotypes were, as a rule, not exclusive to a given island or habitat. High level of genetic divergence and nucleotide saturation was found in closely related morphological designated species, demonstrating possible disparities between currently defined taxonomic units based on morphology and molecular phylogenies of Staphylinidae. This study found evidence of cryptic speciation in the Atheta
fungi (Gravenhorst) species complex which had thus far remained undetected. Similar trends were found for Oligota parva
Kraatz, Oxytelus sculptus Gravenhorst, Oligota pumilio Kiesenwetter. Previous studies with lower taxonomical resolution may have underestimated the biotic diversity reported in the Azores in comparison to other Macaronesian archipelagos.
Key words: habitat type, landscape fragmentation, geographic isolation, genetic variability, Staphylinidae, rove beetles. Introduction
Remote volcanic oceanic archipelagos are usually char-acterized by high endemism and represent a significant portion of terrestrial biodiversity (Holland and Hadfield, 2002; Whittaker and Fernández-Palacios, 2007; Martín
et al., 2008). The archipelago of the Azores possesses a
nutrient rich volcanic soil that supports an abundant flora. As a result, the Azores are recognized worldwide as important conservation hotspots (Myers, 2000; Dias, 2007), despite the high level of landscape fragmentation and anthropogenic impact experienced in its islands (Cardoso et al., 2009; 2010; Meijer et al., 2011).
In comparison to other Macaronesian archipelagos, the Azores have comparatively lower endemic diversity (Borges et al., 2008; 2010a; 2010b; Arechavaleta et al., 2010). They are characterized by the ubiquitous pres-ence of shared endemics across islands and few single island endemics (Borges et al., 2010b). Recent studies have reported a paucity of species radiation for the flora (Carine and Schaefer, 2010) and fauna (Amorim et al., 2012) of the Azores. Reasons for this phenomenon have been debated, and they range from: 1) Lack of past cli-matic fluctuations (Carine and Schaefer, 2010); 2) Low taxonomical species resolution and species underesti-mates (Schaefer et al., 2011a); 3) Relatively recent geo-logical origin and thus in the first stages of their bio-logical evolution (Borges and Hortal, 2009; Triantis et
al., 2012) and 4) Low habitat heterogeneity (Triantis et al., 2012). The interplay between these factors may
have dictated the comparative lower level of taxonomic
diversity in the Azores, and the lower level of endemics may have facilitated dispersal within the archipelago.
Landscape genetic studies could help to further clarify the patterns observed in the distribution, speciation and radiation of species in the Azores. Landscape genetics has been used as a tool in strategies to manage biodiver-sity, with the aim of discerning the impact of ecological, anthropogenic and geographic constrains on the distribu-tion of genetic variability, as well as evoludistribu-tionary proc-esses in complex landscape matrices (Manel et al., 2010; Segelbacher et al., 2010; Sommer et al., 2013). Molecu-lar data have only recently been incorporated into biodi-versity studies in the Azores (Carine and Schaefer, 2010; Schaefer et al., 2011a; 2011b; Amorim et al., 2012), and they have been focused mainly on Azorean plants (Ru-meu et al., 2011; Schaefer et al., 2011a; 2011b; Silva et
al., 2011; Martins et al., 2013) and more recently on Tarphius beetles (Amorim et al., 2012), gastropods
(Jor-daens et al., 2009, Dayrat et al., 2011) and birds (Rodri-gues et al., 2013). A common conclusion from these works is that to better understand patterns of distribution of genetic variation in the Azores, studies with more ro-bust genetic data (e.g., number and length of genes pro-filed) are necessary. More than half of the endemic spe-cies found in the Azores are arthropods (Borges et al., 2010b), rendering this group an optimal choice for the study of the distribution of genetic variability in popula-tions isolated on oceanic islands and fragmented habi-tats. Arthropods have been used in landscape conserva-tion studies, to test for the correlaconserva-tion between genetic diversity and anthropogenic influence (Heidinger et al.,
2013). One of the reasons species on oceanic islands are of particular interest is due to their isolation from mainland species pools (Gillespie and Roderick, 2002; Whittaker and Fernández-Palacios, 2007). The Azores have been inhabited by humans for the past 600 years. Anthropogenic effects on the archipelago have given rise to a variable landscape consisting of mainly pastures interspaced with commercial production forests of
Cryp-tomeria japonica (L.f.) D.Don and few remnants of
pris-tine forests. The cultural and natural history of the Azores makes these islands an ideal environment to study how landscape fragmentation, and anthropogenic influence across a large range of habitats, influences the genetic distribution of arthropod diversity.
In order to study the distribution and genetic variabil-ity of the arthropod fauna of the Azores, we have cho-sen rove beetles (Coleoptera Staphylinidae) as a model group, since they occupy a wide diversity of habitats in the Azores. Staphylinids represent one of the most di-verse lineages of arthropods, inhabiting practically all terrestrial niches (Thayer, 2005). To date 47,744 species of Staphylinidae have been reported worldwide (Thayer, 2005) with 117 described from the Azores (Borges et
al., 2010a). Rove beetles are a common and
ecologi-cally important component of soil fauna and known as potential bioindicators of environmental quality (Bohac, 1999; Rainio and Niemela 2003; Basset et al., 2004; Hodkinson and Jackson 2005; Pohl et al., 2007). De-spite their abundance and widespread occurrence, in-formation as to their ecological role is scarce (Chatzi-manolis, 2004) and limited to few species (Dennis et al., 1997; Krooss and Schaefer, 1998; Andersen and Eltun, 1999; Byers et al., 2000; Caballero et al., 2009). Studies of staphylinids on insular ecosystems are limited to spe-cies lists (Assing, 2002; Klimaszewski et al., 2002; Borges et al., 2010a; Théry and Leschen, 2013). More-over, the taxonomic treatment of members of this group has been difficult due to the paucity of diagnostic mor-phological characters. As a result, the taxonomic and phylogenetic relationship among tribes has been ex-tremely challenging (Anderson and Ashe, 2000; Ashe, 2005; Beutel and Molenda, 1997; Caterino and Vogler, 2005; Chatzimanolis et al., 2010; Elven et al., 2010; 2012; Gutiérrez-Chacón et al., 2009; Hansen, 1997; Thayer, 2005; Thomas, 2009; Osswald et al., 2013).
This study aims to determine whether anthropogeni-cally fragmented insular landscape matrices influence the spatial distribution and genetic variability of staphylinids on the Azores, as well as to determine the taxonomic relationship of Azorean taxa to the four in-formal groups of Staphylinidae sensu Lawrence and Newton (1995), i.e. Omaliine, Tachyporine, Oxyteline and Staphylinine), vis-à-vis published COI data from other species published in Genbank. We hypothesize that the distribution, species richness, abundance and genetic diversity of staphylinids will change with increasing an-thropogenic influence. We stress that the genetic analysis in this work aims to determine the taxonomic relation-ship of the species collected on the Azores in relation to other species for which genetic information is available, not to establish a definitive molecular phylogeny of the family. Such a task is beyond the scope of this work.
Materials and methods
Habitat selection and sampling
Habitats were selected to represent the land-use diver-sity of the Azorean archipelago, located across the Mid-Atlantic ridge (36°35'-39°43'N 24°45'-31°17'W). The five islands sampled were selected on the basis of the proportion of land present in a pristine and fragmented state along a gradient of anthropogenic influence, i.e., São Miguel (SMG) with a large proportion of land allo-cated to agriculture and low/medium proportion of natu-ral habitats; Terceira (TER) with a large proportion of land dedicated to pastureland and a medium/high portion of natural habitats; Pico (PIC) with a high pro-portion of land dedicated to pastureland and me-dium/high proportion of natural habitats, classified as UNESCO World Heritage cultural landscapes; Flores (FLO) with a small proportion of land dedicated to pas-tureland and a high proportion of natural communities, classified as UNESCO Biosphere Reserve; Santa Maria (SMR) with a high proportion of land allocated to agri-culture, although not all presently used, and a small proportion of natural habitats, albeit of unique conserva-tion value. The habitats sampled for this study, depict-ing gradients of increasdepict-ing anthropogenically influenced habitats were as follows; Herbaceous gradient: mead-ows, semi-natural pastures, intensive pasture land and corn fields. Arborescent gradient: pristine forest of
Lau-rus azorica (Seub.) Franco, invasive forests of Pitto-sporum undulatum Vent., production forests of C. ja-ponica, and orchards of Citrus sinensis L. (see
Marcelino et al., 2013 for further description of the habitats and supplemental material (figure S1, supple-ment 1) for further description of the islands and distri-bution of biodiversity).
To obtain a representative sampling of Staphylinidae species in each habitat, pitfall traps (PF) and Berlese-Tullgren traps (BT) were used. A total of sixteen sites/island were sampled for Staphylinidae diversity, these spanned eight habitats (2 replicates per habitat type and island sampled, with two transects of 100 m each). Habitats were ca. 5 km-20 km apart. PF consisted of plastic cylinder cups, 78 mm deep and 42 mm in di-ameter, filled with ca. 80 ml of 96% ethanol and 0.04% commercial liquid dishwashing detergent to break the surface tension of the ethanol. PF were placed in the soil with the rim flushed with the surface and covered with a plastic plate at ca. 3 cm above the trap to avoid desicca-tion, flooding or insectivore predation. Traps remained in the soil for 7 days prior to removal. Thirteen PF were set up in each transect. The number and distance of BT traps was equal to PF traps. Soil and litter samples, as well as sweeping and vacuum samples, were collected at each PF trap area. Soil and litter samples were stored at 10 °C prior to being placed in BT traps. Sweeping and vacuum samples were also stored at 10 °C and sub-sequently sorted manually. BT consisted of two plastic darkened containers assembled together to provide an upper vented area (14 cm diameter × 11.5 cm high) with four openings (1 cm diameter covered with a 0.3 × 0.3 mm diameter mesh), coupled to a 15 W lamp on top, and a collecting area at the bottom (13 cm diameter × 10
cm high) partially filled with ca. 80 ml of the same liq-uid mixture used in PF. Litter samples were processed in the same device as described above but with a 1.8 × 1.8 mm mesh attached to a plastic funnel in the center of the device that filtered small invertebrates. The number and distance of BT traps was equal to the PF traps. Lit-ter samples remained in BT traps for 72 h before re-moval, storage and taxonomic identifications. All sam-pling was conducted in June 2009 to ensure similar cli-matic and vegetation conditions. Species inventory completeness was determined by plotting species rare-faction curves using EstimateS (Colwell, 2011). Sam-pling representativeness was determined as:
SObserved species/SJackknife1 species estimator
We selected the Jackknife estimator since it is one of the most robust when using incidence data (Hortal et al., 2006).
Taxonomic identifications
Specimens were sorted under a stereomicroscope and assigned to morpho-species using reference image gal-leries generated for the study (www.eden-azores.com). Reference voucher specimen collections were generated for each habitat type and islands surveyed and were stored at the Entomology Laboratory, Department of Biology of the University of the Azores. They are part of the Arthropoteca of the University of the Azores (ARZ), at Ponta Delgada, Portugal. All specimens were preserved in 96% ethanol and stored in a chamber at 4 °C, prior to higher taxonomical identifications (genus and species).
All specimens of staphylinids were identified to spe-cies based on morphological characters by a member in the group (PAVB), using as a reference the Entomoteca Dalberto Teixeira Pombo, at University of the Azores, Terceira, Portugal. Subsequently these were recon-firmed by a Staphylinidae systematics expert (Volker Assing), using dissecting techniques. A list of species can be found in table S1 (supplement 1).
DNA extraction
Single specimens preserved in ethanol were allowed to air dry prior to DNA extraction using the QIAamp® DNA microkit (QIAGEN Inc., Chatsworth, CA). The nearly complete Cytochrome Oxidase (COI) gene (1265 bp) was amplified using primers indicated in table S2 (supplement 1) and Illustra PuReTaq™ Ready-To-Go PCR™ beads (GE, Fairfield, CT) using the following protocol: 95 °C 2 min followed by (95 °C 30 sec, 50 °C 30 sec, 72 °C 2 min) for 40 cycles. PCR amplifications were verified by gel electrophoresis using 1% agarose gel with 1 µl GelGreen™ Nucleic Acid Gel Stain, 10,000X (Biotium Inc., Hayward, CA). If multiple non-specific bands were present, PCR products were cleaned by excising the target length band from the gel using a sterile scalpel and cleaned with the Zymoclean™ Gel DNA recovery kit (Zymo Research, Irvine, CA). If only a single band was present, PCR reactions were cleaned using QIAquick PCR purification Kit (QIAGEN, Va-lencia, CA) according to the manufacturer’s protocol. PCR reactions were carried out in a DNA Engine Peltier
thermal cycler (Bio-Rad, Hercules, CA). PCR products were purified using the QIAquick PCR purification kit (QIAGEN, Valencia, CA) according to the manufac-turer’s protocol. PCR products were measured using a Nanodrop (Thermo Scientific, Hudson, NH). Five ng of DNA per 100 bp was used in each 20 µl sequencing re-action, as well as 1 µl of primer and a mixture of 2:1:1 BigDye Terminator v3.1; dGTP BigDye Terminator v3.0; Big Dye Terminator v1.1, and v3.1 5X sequencing buffer (Life Technologies, Carlsbad, CA). The follow-ing sequencfollow-ing protocol was used: 95º C 2 min followed by (98º C for 10 s, 50º C for 5 s, 60ºC for 4 min) for 30 cycles. Sequencing products were cleaned using PER-FORMA Ultra 96-well Plate (Edge Bio, Gaithersburg, MD) and run on a ABI 3730 automated capillary Se-quencer (Life Technologies, Carlsbad, CA) at the Keck Center for Comparative and Functional Genomics, Uni-versity of Illinois at Urbana-Champaign, USA. Raw se-quence data were edited using Sequencher™ 4.7 (Gene Codes Corporation, Ann Arbor, MI).
Sequences were aligned using a combination of algo-rithmic (Clustal W software) and manual alignment, and subsequently analyzed using PAUP 4.0b10 (Swofford, 2002). Start codon positions were determined using the mitochondrial genome of Drosophila yakuba Burla (Genbank accession number NC001322) as a reference (Clary and Wolstenholme, 1985). Differences in the genotype of individuals were determined with pairwise distances in PAUP (table S3, supplement 1). Haplotypes were determined for each species between islands and between habitats within island (table S4, supplement 1). Genetic analysis
The COI gene was sequenced for a total of 142 indi-viduals in subfamilies and tribes of the four informal groups of Staphylinidae sensu Lawrence and Newton (1995), found in the Azores. The COI sequences ob-tained from Azorean specimens were aligned with those of other staphylinid species in Genbank. The final data set of 275 individuals is comprised of: 54 species in 13 tribes of the Tachyporine group; 57 species in 7 tribes of the Staphylinine group; 23 species in 2 tribes of the Omaliine group; and 14 species in 2 tribes of the Oxyte-line group. The complete list of the 275 individuals used in the genetic analysis is given in table S5 (supplement 1).
We evaluated the saturation level of the nucleotide se-quences for the four Staphylinidae groups individually using the Akaike information criterion implemented in MrBayes. Graphic representations were obtained by plotting nucleotide corrected pairwise distances, ob-tained in PAUP (Swofford, 2003), using the best substi-tution model identified in jModeltest v0.1.1 (Posada 2008; Guindon and Gascuel, 2003). Maximum Likeli-hood was then used to measure pairwise genetic dis-tances between nucleotide sequences and plotted against uncorrected pairwise distances. Saturation was assessed as deviation from a linear unsaturated data set as estab-lished by Jeffroy et al. (2006). In addition, the nucleotide data sets for each Staphylinidae group sensu Lawrence and Newton (1995) was partitioned by codon position and saturation estimated independently for each codon position during the analyses. Four Markov chains with a
temperature of 0.5 °C were run for 30 million genera-tions. Samples were taken every 1000 generagenera-tions. The first 15000 trees produced were discarded during burn in.
We inferred the taxonomic relationship of taxa in the four groups of Staphylinidae found in the Azores, based on their COI genetic profile, and published data from species found across the globe, using a Neighbor Join-ing (NJ) analysis as implemented in PAUP version 4.0a147. The outgroup taxa was selected based on the taxonomical proximity to all tribes included in the analysis. We chose a species in the superfamily Staphylinoidea (Coleoptera, infraorder Staphylinifor-mia), in which the family Staphylinidae is included, (see table S5, supplement 1). The output tree was visualized using FigTree v1.3.1 (Rambaut, 2009).
Results
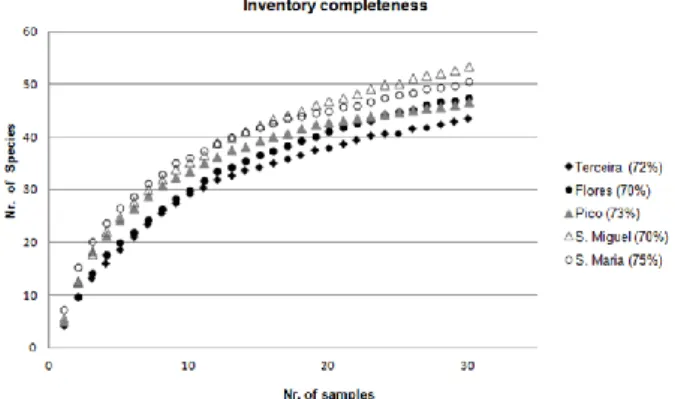
Inventory completeness for Staphylinidae collected dur-ing this project in the Azores archipelago reached 70-75%, depending on the island sampled (figure 1). A to-tal of 38 species were found (table S1, supplement 1). The distribution of species richness across islands, and within habitat types sampled, reflect anthropogenic in-fluence, ranging from three species in the less anthropo-genic impacted habitats to 18 species in the more influ-enced habitats (figure S1, supplement 1). Disparities were more evident in the degree of abundance, ranging from eight specimens in less anthropogenic influenced habitats, to 260 in the most anthropogenic influenced habitats (figure S1, supplement 1). Species turnover was more evident in highly anthropogenically influenced habitats such as corn is all the islands sampled except S. Miguel and invasive forest in Flores Island. With few exceptions, habitat types with high anthropogenic influ-ence, i.e. corn, intensive pasture and orchards, presented higher abundance when compared with less anthropo-genically influenced habitats such as pristine forests, semi natural pastures and meadows. Santa Maria and Pico had the highest abundance of staphylinids (figure S1, supplement 1). Geologically, these are the oldest and youngest islands of the archipelago, respectively.
In order to discriminate species diversity at a finer resolution as well as to test the level of genetic diversity at the population level we obtained COI mitochondrial DNA sequences for all morphospecies represented by more than three specimens per locality. In total we se-quenced the COI gene for 142 staphylinids. Specimens sequenced belonged to the four informal Staphylinidae groups sensu Lawrence and Newton (1995) i.e., Tachy-porine, Staphylinine, Omaliine and Oxyteline groups. The nucleotide dataset showed high levels of saturation (figures S2 and S3, supplement 1), therefore, to avoid biased inferences, all analyses were conducted using the corresponding amino acid sequences. Within the taxo-nomically assigned putative species, amino acid se-quence divergence (table S3, supplement 1) for Tachy-porine, ranged between 0% [Atheta fungi (Gravenhorst)] and 10.3% (Oligota pumilio Kiesenwetter) while across species it was 17.8%; for Staphylinine we observed 0-0.25% within species [Quedius simplicifrons Fairmaire,
Figure 1. Species rarefaction curves and inventory
completeness (%) for each island based on the number of species collected (Jackknife richness estimator).
Rugilus orbiculatus orbiculatus (Paykull)] and 13.06%
across species; Oxyteline species divergences varied from 0.8% [Anotylus nitidifrons (Wollaston)] and 15.57% (Oxytelus sculptus Gravenhorst) within species, and 17.9% across species; Omaliine divergence within species was 3.6% (Phloeonomus punctipennis Thom-son) and 14.2% across species. The invasive A. fungi was the most widespread and abundant species across all islands (figure S4, supplement 1). Analysis of COI sequences for this species shows no geographic differ-entiation, as none of the haplotypes were found to be exclusive to a given island (figure 2 and table S4, sup-plement 1). The majority of haplotypes were dissemi-nated across habitats and islands sampled, showing amino acid level pair-wise divergences ranging from 0.2%-5.7%. The second most widespread species, the invasive A. nitidifrons, was found in five islands and six of the eight habitats, and had four haplotypes differing from 0.16%-0.87% at the amino acid level. This species was followed by the native R. orbiculatus orbiculatus (five islands, four habitats, two haplotypes, 0.24% di-vergence) and the native O. pumilio (four islands, three habitats, two haplotypes, 10.3% divergence).
The NJ analysis of the COI gene, for the complete Staphylinidae dataset, shows one large core group of genera for three informal Staphylinidae groups sensu Lawrence and Newton (1995), i.e., Oxyteline, Staphylininae and Tachyporinae (figure 2) (figure S5, supplement 2 for high resolution imaging). Clades of these major core groups of genera seldom included gen-era from other groups. The Omaliine group does not show a large core group of taxa, but several smaller groups dispersed across the middle and base of the tree.
Discussion
Staphylinidae in the Azores presented an evenly distrib-uted pattern of species richness across islands, indica-tive of low habitat fidelity and strong dispersal capabili-ties. This was not the case for the different habitats in the gradients of anthropogenic influence, where species richness and abundance increased as human impact rose. This pattern could be a consequence of sampling
F ig ure 2. Neig hb or J oin in g tr ee f or Stap hy lin id ae s pec ies in s ub fam ilies an d tr ib es o f th e fo ur in fo rm al gr ou ps o f Stap hy lin id ae s en su L awr en ce an d N ewto n (1 99 5) , f ou nd in th e Azo res an d wo rld wid e, f or wh ich C OI s eq ue nce s wer e gen er ated ( 12 65 b p) o r av aila ble in Gen ba nk , resp ec tiv ely . Stap hy lin id ae g ro up s co lo ur ed as fo llo ws: T ac hy-po rin e (g ree n) , Om aliin e (r ed ), Ox ytelin e (b lac k) an d Sta ph ylin in e (b ro wn ). Ou tg ro up wa s co lo ur ed in b ro wn . T ax a ar e fo llo wed b y th eir Gen ban k ac ce ss io n nu m ber , tr ib e or s ub fam ily ( *) wh en tr ib e was no t av ailab le in Gen ban k. C OI s eq uen ce s fo r tax a in b ol d wer e gen er ated in th is s tu dy . I slan d of o rig in f or ea ch s pe cies is s pec if ied as fo l-lo ws: SMG - Sao Mig uel ; T E R T er ce ir a; SMR San ta Ma ria ; FLO - Flo res ; PIC - Pico .
A high-resolution figure is available in the supplemental material (figure S5 supplement 2). Oxytelus sculptus JX626139 SMG (Oxytelini)
Oxytelus sculptus JX626138 SMG (Oxytelini)
Platystethus arenarius DQ155733 (Oxytelini) Anotylus tetracarinatus DQ155804(Oxytelini) Oxytelus sculptus JX626142 PIC (Oxytelini) Anotylus rugosus DQ155729 (Oxytelini)
Anotylus anustus DQ156068 (Oxytelini) Anotylus sculpturatus DQ155747 (Oxytelini) Anotylus nitidrifons JX626131 PIC (Oxytelini) Anotylus nitidrifons JX626123 TER/FLO/SMR/PIC/SMG (Oxytelini)
Anotylus nitidrifons JX626132 SMG/FLO (Oxytelini) Carpelimus corticinus JX626122 FLO (Oxytelini) Carpelimus corticinus DQ155778 (Oxytelini)
Carpelimus elongatulus DQ156050 (Oxytelini) Carpelimus similis DQ155978 (Oxytelini) Carpelimus bilineatus DQ155950 (Oxytelini)
Bledius femoralis DQ222015 (Oxytelini) Quedius antipodus GU377393 (Quediini)
Gyrohypnus fracticornis JX626082 SMG (Xantholinini) Gyrohypnus fracticornis JX626083 PIC (Xantholinini)
Atanygnathus acuminatus GU377363 (Staphylinini) Atanygnathus sp. GU377364 (Staphylinini)
Holisus sp. GU377375 (Staphylinini) Gabrius nigritulus JX626105 SMG (Philonthini) Xanthopygus sp. GU377403 (Staphylinini)
Nausicotus spectabilis GU377378 (Staphylinini) Platydracus sp. GU377390 (Staphylinini)
Naddia sp. GU377377 (Staphylinini) Ontholestes murinus GU377384 (Staphylinini)
Platydracus cinnamopterus GU377389 (Staphylinini) Hesperus sp. GU377374 (Staphylinini)
Ocypus olens JX626093 SMG/PIC/SMR/FLO (Staphylinini) Ocypus aethiops JX626093 TER (Staphylinini) Ocypus olens GU377382 (Staphylinini) Tasgius morsitans GU377398 (Staphylinini) Tasgius predator GU377399 (Staphylinini)
Dinothenarus saphyrinus GU377369 (Staphylinini) Belonuchus mimeticus GU377366 (Staphylinini)
Tolmerinus sp. GU377360 (Staphylinini)
Tympanophorus sp. GU377401 (Staphylinini) Elecatopselaphus sp. GU377371 (Staphylinini) Xenopygus analis GU377404 (Staphylinini) Nordus fungicola GU377380 (Staphylinini)
Oligotergus sp. GU377383 (Staphylinini) Gastrisus sp. GU377372 (Staphylinini)
Hesperopalpus sp. GU377388 (Staphylinini) Hadropinus fossor GU380341 (Staphylinini)
Hadrotes crassus GU377373 (Staphylinini) Quedius simpliciformis JX626084 SMG (Quediini) Quedius curtipennis JX626087 SMG/PIC (Quediini) Quedius simpliciformis JX626108 SMG (Quediini) Quedius simpliciformis JX626086 SMR (Quediini) Quedius simpliciformis JX626109 PIC/TER (Quediini) Quedius picipes GU377395 (Quediini) Quedius suturalis GU377397 (Quediini)
Quedius molochinus GU377396 (Quediini) Thinopinus pictus GU377400 (Staphylinini) Veileius dilatatus GU377402 (Staphylinini)
Cafius seminitens GU377368 (Staphylinini) Philonthus splendens GU377387 (Philonthini) Neobisnius occidentoides GU377379 (Staphylinini)
Gymnusa variegata GQ980860 (Gymnusini) Anaquedius vernix GU377358 (Staphylinini) Quedius cruentus GU377394(Quediini)
Othius punctulatus GU377385 (Othiini) Quediocafus insolitus GU377392 (Quediini) Nudobius pugetanus GU377381(Xantholinini)
Myllaena audax GQ980881 (Myllaenini) Algon sp. GU377356 (Staphylinini) Indoquedius sp. GU377376 (Staphylinini) Arrowinus relictus GU377361 (Arrowinini)
Diochus sp. GU377370 (Diochini) Atrecus punctiventris GU377365 (Othiini)
Bolitogyrus sp. GU377367 (Staphylinini) Lesteva heeri DQ155752 (Omaliini)
Platyprosopus sp. GU377391 (Platyprosopini) Tachynus proximus GQ980859 (Tachyporini) Olophrum piceum DQ155842 (Omaliini) Dropephylla ioptera DQ155920 (Omaliini)
Lesteva pubescens DQ156049 (Omaliini) Lesteva longoelytrata DQ156029 (Omaliini) Dropephylla vilis DQ155756 (Omaliini)
Glypholoma sp. DQ155756 (Glypholomatinae*) Anthobium atrocephalum DQ155885 (Omaliini)
Anthobium unicolor DQ155841 (Omaliini)
Coproporus pulchellus JX626143 SMG (Tachyporini)
Habrocerus capillaricornis DQ155843 (Habrocerinae*) Philonthus sp.1 JX626107 SMG (Philonthini) Tachyporus nitidulus JX626001 SMG/SMR (Tachyporini) Sepedophilus lusitanicus JX626008 SMR (Tachyporini)
Tachyporus nitidulus JX626000 SMG (Tachyporini) Tachyporus dispar JX626004 FLO (Tachyporini)
Tachyporus formosus AJ293031 (Tachyporini)
Phloeonomus punctipennis JX626112 SMG/FLO (Omaliini) Paraphloeostiba gayndahensis JX626098 PIC/FLO/SMR (Omaliini)
Omalium rugatum DQ155962 (Omaliini) Omalium caesum DQ155696 (Omaliini)
Omalium rivulare DQ155713 (Omaliini)
Phloeonomus sp. JX626120 SMR (Omaliini) Phloeonomus punctipennis JX626121 SMR (Omaliini)
Aleochara clavicornis JX626137 PIC (Aleocharini)
Aleochara laticornis AJ293048 (Aleocharini) Aleochara moerens GQ980861 (Aleocharini) Aleochara clavicornis AJ293040 (Aleocharini) Cordalia obscura GQ980864 (Faligriini)
Scaphidium quadrimaculatum DQ221993 (Scaphidiinae) Placusa sp. prope tachyporoides GQ980883 (Placusini)
Haplandria lateralis GQ980872 (Hoplandriini) Haplandriini gen. sp. AJ293039 (Hoplandriini) Haploglossa villosula AJ293037 (Oxypodini)
Atheta fungi JX626053 PIC/FLO/SMR/SMG (Athetini) Atheta fungi JX626068 SMR (Athetini)
Atheta (Mycetota) pasadenae GQ980921(Athetini)
Dadobia immersa GQ980953(Athetini) Amischa analis GQ980895(Athetini)
Acronata sp. prope austiniana GQ980886 (Athetini) Atheta (vaga-group) vaga GQ980930 (Athetini) Atheta (Ceritaxa) pervagata GQ980906 (Athetini)
Atheta vestita AJ293032 (Athetini) Liogluta nigropolita GQ980938 (Athetini)
Ecitophya gracillima GQ980863 (Ecitocharini)
Atheta atramentaria JX626015 SMG (Athetini)
Atheta (s. str.) graminicola GQ980928 (Athetini) Boreophilia hyperborea GQ980933 (Athetini)
Zyras collaris AJ293033 (Lomechusini) Zyras haworthi AJ293034 (Lomechusini)
Atheta fungi sp. 1 JX626051 PIC (Athetini)
Acronata sp. prope assecla GQ980884 (Athetini)
Atheta fungi JX626069 SMG (Athetini) Atheta fungi JX626071 TER (Athetini) Atheta fungi JX626070 SMG (Athetini)
Drusilla canaliculata AJ293035 (Lomechusini) Drusilla canaliculata GQ980873 (Lomechusini)
Phihygra debilis GQ980942 (Athetini) Pella caliginosa GQ980878 (Lomechusini) Atheta (Alaobia) gagatina GQ980901(Athetini) Stethusa spuriella GQ980951(Athetini)
Lypoglossa lateralis GQ980887 (Athetini) Atheta (Alaobia) membranata GQ980903(Athetini) Atheta (Microdota) subtilis GQ980919 (Athetini) Atheta (Datomicra) celata GQ980910 (Athetini)
Atheta fungi JX626027 FLO/SMR/PIC/TER/SMG (Athetini) Atheta fungi JX626049 SMG (Athetini)
Gyrophaena congrua GQ980867 (Homalotini) Nehemitropia lividipennis GQ980892 (Athetini) Schistoglossa gemina GQ980947 (Athetini)
Atheta fungi JX626052 PIC (Athetini)
Trichiusa ursina GQ980964 (Athetini) Strigota ambigua GQ980893 (Athetini) Dinaraea aequata GQ980935 (Athetini) Atheta (ravilla-group) ravilla GQ980925 (Athetini)
Geostiba circellaris GQ980954 (Athetini) Acyclophorus sp. GU377355 (Staphilinini) Atheta (Dimetrota)hampshirensis GQ980916 (Athetini)
Silusida marginella GQ980870 (Homalotini) Hydrosmecta sp. GQ980955 (Athetini) Amidobia talpa GQ980898 (Athetini)
Atheta (Alaobia) pallidicornis GQ980904 (Athetini) Atheta fungi JX626097 FLO (Staphilinini) Atheta fungi JX626021 FLO/PIC/SMR (Staphilinini) Atheta fungi JX626024 PIC/SMR (Staphilinini)
Mocyta fungi GQ980888 (Staphilinini) Mocyta scopula GQ980890 (Staphilinini) Oxypoda longipes AJ293036 (Oxypodini) Oxypoda praecox GQ980882 (Oxypodini)
Bolitochara pulchra GQ980865 (Homalotini) Tomoglossa ornatella GQ980952 (Athetini)
Euplectus infirmus JX626111 TER (Euplecetini)
Plectophloeus nitidus DQ155919 (Pselaphinae*) Euplectus nanus DQ155918 (Euplecetini)
Oligota pumilio JX626013 SMG/FLO/SMR/TER (Hypocyphtini)
Neophonus sp. DQ222007 (Omaliini)
Rugilus orbiculatus orbiculatus JX626072 SMG/FLO/SMR/TER/PIC (Paederinae*) Rugilus orbiculatus orbiculatus JX626801 PIC (Paederinae*) Astenus sp. GU377362 (Paederinae*)
Oligota pumilio JX626014 (Hypocyphtini)
Astenus sp. GU377362 (Paederinae*) Metopsia clypeata DQ155840 (Proteininae*)
Brachynopus scutellaris JQ267152 (Scaphidiinae) Megarthrus depressus DQ156067 (Proteininae*)
Notosaprinus sp. KF801964 (Staphyliniformia: Histeridae) 0.05 substitutions/site Oxyteline Staphylinine Omaliine Tachyporine Omaliine Tachyporine Tachyporine Staphylinine Staphylinine Omaliine
in artificially nutrient enriched habitats composed of depauperate communities (i.e., low β diversity and high species turnover) such as intensive pasture land, corn fields and orchards which were dominated by the most tolerant and competitive introduced arthropod species (data not shown). Although natural and anthropogeni-cally impacted habitats seemed close in species rich-ness, they were in fact very distinct since introduced cosmopolitan species did not contribute to the unique species pool characteristic of a given community as-semblage, such as the one in pristine forests, meadows and semi natural pastures. These findings concur with previous reports that also found a broad dissemination of generalist species across the Azorean islands and the existence of few single island endemics (Borges et al., 2010a). Similar results have also been shown for popu-lations of highly vagile insects in central Europe (Černá
et al., 2013). Landscape fragmentation may be of
cru-cial importance for the success of some species and of less consequence to highly vagile insects that can easily move across a fragmented matrix. Different habitats may act as stepping stones for dispersal across a highly fragmented landscape (Finn et al., 2007; Louy et al., 2007; Prevedello and Vieira, 2010; Bossart and Antwi, 2013). This may be the case for staphylinids in the Azores, which do not seem to have specific macrohabi-tat requirements and are ubiquitously distributed across the fragmented matrices, particularly in more anthropo-genic impacted habitats (both in abundance and number of species). Moreover, the strong influence of human activity in the region suggests that passive dispersal may be a common bridge for inter-island colonization in an-thropophilic staphylinids. This has been the case for the Japanese beetle Popillia japonica Newman in the Azores (Martins and Simões, 1985). The low habitat fidelity and strong dispersal capabilities of Staphylini-dae in the Azores was also supported by genetic data. Human induced landscape fragmentation, as well as geographic distance between habitat patches and is-lands, was not correlated with genetic differentiation and isolation. Genetic diversity among Staphylinidae populations, subjected to geographic and anthropogeni-cally influenced barriers, does not appear affected by isolation (in opposition to species richness and abun-dance), as haplotypes represented with multiple speci-mens were, as a rule, not exclusive to a given island. This contradicts the expected decrease in genetic con-nectivity with increasing geographic distance (Wright, 1943; Slatkin, 1993). The absence of an effect of land-scape fragmentation on genetic isolation has been re-ported in other animals (Sumner et al., 2004; Banks et
al., 2005; Richmond et al., 2009; Dixo et al., 2009).
We compared the Staphylinidae taxa obtained from the Azores archipelago to taxa present in Genbank based on a large dataset of complete and partial mtDNA COI (1265 bp) sequences, using Neighbor Joining analysis. As previous studies have attested with analysis at the group or tribe level (Maus et al., 2001; Thomas, 2009; Chatzimanolis et al., 2010; Elven et al., 2010; 2012; McKenna et al., 2015), our results suggest that the relationships for species and tribes of Staphylinidae are unresolved. The Tachyporine and Oxyteline are the
Staphylinidae groups sensu Lawrence and Newton (1995) in which taxonomical affinities seem more ro-bust. Only seldom we found genera interspersed among other groups. Conversely, the Omaliine and to a smaller extent the Staphylinine, showed minor groups and iso-lated taxa distributed among other Staphylinidae groups. This is not surprising since taxonomic classifications of rove beetles are largely based on poorly understood ana-tomical and morphological characters (Maus et al., 2001). The taxonomic and phylogenetic relationships among tribes have been reported as extremely challeng-ing (Anderson and Ashe, 2000; Ashe, 2005; Beutel and Molenda, 1997; Caterino and Vogler, 2005; Chatzi-manolis et al., 2010; Elven et al., 2010; Gutiérrez-Chacón et al., 2009; Hansen, 1997; Thayer, 2005). The recognition of non-monophyletic tribes and phyloge-netic arrangements has been extensively reported in mo-lecular analyses focusing on particular subfamilies or tribes of Staphylinidae, namely the Tachyporine and Staphylinine groups (Ashe, 2005; Elven et al., 2010; 2012; Osswald et al., 2013; Maus et al., 2001; Thomas, 2009; Chatzimanolis et al., 2010), as well as the Staphyliniformia (Caterino et al., 2005). Moreover, dif-ficulties arising from the high level of genetic diver-gence and nucleotide saturation in closely related spe-cies of this study are documented here. These latter findings corroborate the known disparities between cur-rently defined taxonomic units and the existing molecu-lar phylogenies of Staphylinidae.
The A. fungi complex of species suggests that cryptic speciation in Staphylinidae is extensive in the Azores, and probably beyond this region (Volker Assing per-sonal communication), and has remained undetected due to the lack of taxonomic studies. Cryptic speciation has been documented in many biological groups (Bickford et al., 2006). Low species identification reso-lution and underestimates of species number in the Azores have been previously described as a probable cause for the presumably lower biota diversity found in the Azores when compared with other Macaronesian archipelagos (Borges et al., 2008a; 2010a; Arechavaleta
et al., 2010; Schaefer et al., 2011a; Amorim et al.,
2012), Lack of detailed taxonomic work also can ex-plain the disparities between current assigned taxonomic units and the genetic profiles of Staphylinidae taxa. A work integrating morphological and molecular data, across the four Staphylinidae lineages, would help to further resolve the intricate taxonomic relationships of the family, most probably rearranging tribes and species into a more cohesive classification.
In this study we also determined the effect of anthro-pogenic influence, landscape fragmentation and isola-tion, in the distribution of species richness and the ge-netic variability of rove beetles, collected from an iso-lated archipelago, the Azores, which has lost more than 95% of its native vegetation in less than 600 years. Us-ing a traditional measure of biodiversity, i.e. number of species, coupled with genetic diversity, a measure not commonly used in community ecology studies, we ob-served that Staphylinidae in the Azores present low habi-tat fidelity and strong dissemination capabilities, inde-pendent of landscape fragmentation. It is likely that the
unresolved taxonomic status of a significant proportion of staphylinid taxa has contributed to underestimating the diversity of members of this group in the Azores. This study gives evidence for the existence of cryptic speciation among some taxa indicating the need for fur-ther morphological and molecular studies to be able to elucidate the biodiversity of rove beetles in the Azores.
Our study indicates that anthropogenic influence in different habitats can contribute to the increase in cies richness and abundance of a few opportunistic spe-cies, with limited genetic variability, possibly compro-mising the viability of less competitive and unique spe-cies. Retaining genetic variability at the population level is crucial for species resilience in habitats that are con-tinually and drastically altered due to anthropogenic and/or climatic impacts. This research underscores that more attention should be given to the long-term effects of these impacts on species richness. Oceanic islands continue, since Darwin and Wallace, to provide insights into the mechanisms that promote biodiversity, as well as the effects of anthropogenic action.
Acknowledgements
We would like to thank Roberto Resendes and João Moniz for field assistance, and Betty Ujhelyi, Massimo Pessino and Therese Anne Catanach for technical sup-port. We also thank Volker Assing for taxonomic identi-fication of some specimens. This work was supported by a grant from the Fundo Regional Ciência e Tecnolo-gia, Regional Government of the Azores and PROEM-PREGO and the research project “PROBIO, Profiling Reliable Organisms as Bioindicators: an integrated ap-proach for island systems” financed by FLAD (Funda-ção Luso-Americana para o Desenvolvimento). This study was also financed by Portuguese National Funds, through FCT – Fundação para a Ciência e a Tecnologia, within the project UID/BIA/00329/2013.
References
AMORIM I.R.,EMERSON B.C.,BORGES P.A.V.,WAYNE R.K., 2012.- Phylogeography and molecular phylogeny of Macaronesian Tarphius (Coleoptera: Zopheridae): why so few species in the Azores?.- Journal of Biogeography, 39: 1583-1595.
ANDERSON R.S.,ASHE J.S.,2000.- Leaf litter inhabiting
bee-tles as surrogates for establishing priorities for conservation of selected tropical montane cloud forests in Honduras, Cen-tral America (Coleoptera; Staphylinidae, Curculionidae).-
Biodiversity and Conservation, 9: 617-653.
ANDERSEN A.,ELTUN R., 1999.- Long-term developments in
the carabid and staphylinid (Col., Carabidae and Staphylini-dae) fauna during conversion from conventional to biologi-cal farming.- Journal of Applied Entomology, 124: 51-56. ARECHAVALETA M., RODRÍGUEZ S., ZURITA N., GARCIA A.,
2010.- Lista de especies silvestres de Canarias. Hongos,
plantas y animales terrestres.- Gobierno de Canarias, Spain.
ASHE J.S., 2005.- Phylogeny of the tachyporine group
sub-families and 'basal' lineages of the Aleocharinae (Coleop-tera: Staphylinidae) based on larval and adult characteris-tics.- Systematic Entomology, 30: 3-37.
ASSING V., 2002.- On the Staphylinidae of the Canary Islands.
IX. New synonyms and records, and asystematic rearrange-ment of some endogean and cavernicolous Aleocharinae (Coleoptera).- Vieraea, 30: 45-66.
BANKS S.C.,LINDENMAYER D.B.,WARD S.J.,TAYLOR A.C.,
2005.- The effects of habitat fragmentation via forestry plan-tation establishment on spatial genotypic structure in the small marsupial carnivore, Antechinus agilis.- Molecular
Ecology, 14: 1667-1680.
BASSET Y., MAVOUNGOU J. F., MIKISSA J. B., MISSA O.,
MILLER S.E.,KITCHING R.L.,ALONSO A., 2004.-
Discrimi-natory power of different arthropod data sets for the biologi-cal monitoring of anthropogenic disturbance in tropibiologi-cal for-ests.- Biodiversity and Conservation,13: 709-732.
BEUTEL R.G.,MOLENDA R., 1997.- Comparative morphology
of larvae of Staphylinoidea (Coleoptera, Polyphaga) with phylogenetic implications.- Zoologischer Anzeiger, 236: 37-67.
BICKFORD D.,LOHMAN D.J.,SODHI N.S.,NG P.K.,MEIER R.,
WINKER K.,INGRAM K.K.,DAS I., 2006.- Cryptic species as
a window on diversity and conservation.- Trends in Ecology
and Evolution, 22: 148-155.
BOHAC J., 1999.- Staphylinid beetles as bioindicators.- Agri-culture, Ecosystems and Environment, 74: 357-372.
BORGES P.A.V.,HORTAL J., 2009.- Time, area and isolation:
Factors driving the diversification of Azorean arthropods.-
Journal of Biogeography, 36: 178-191.
BORGES P.A.V.,ABREU C.,AGUIAR A.M.F.,CARVALHO P.,
JARDIM R., MELO I., OLIVEIRA P., SÉRGIO C., VIEIRA P.,
SERRANO A.R.M., 2008.- A list of the terrestrial fungi, flora and fauna of Madeira and Selvagens archipelagos.-
Di-recção Regional do Ambiente da Madeira and Universidade dos Açores, Funchal and Angra do Heroísmo, Portugal. BORGES P. A.V., COSTA A., CUNHA R.,GABRIEL R., G
ON-ÇALVES V.,MARTINS A.F., MELO I., PARENTE M., R APO-SEIRO P.,RODRIGUES P.,SANTOS R.S.,SILVA L.,VIEIRA V.,
2010a.- A list of the terrestrial and marine biota from the
Azores.- Princípia, Cascais, Portugal.
BORGES P. A.V., COSTA A., CUNHA R.,GABRIEL R., G ON-ÇALVES V.,MARTINS A.F., MELO I., PARENTE M., R APO-SEIRO P.,RODRIGUES P.,SANTOS R.S.,SILVA L.,VIEIRA V.,
VIEIRA P.,VIEIRA V., MENDONÇA E., BOEIRO M., 2010b.-
Description of the terrestrial and marine biodiversity of the Azores, pp. 9-33. In: A list of the terrestrial and marine
bi-ota from the Azores (BORGES P.A.V.,COSTA A.,CUNHA R.,
GABRIEL R.,GONÇALVES V.,MARTINS A.F.,MELO I.,P AR-ENTE M.,RAPOSEIRO P.,RODRIGUES P.,SANTOS R.S.,SILVA
L.,VIEIRA V., Eds).- Princípia, Cascais, Portugal.
BOSSART J.L.,ANTWI J.B., 2013.- Species-specific traits pre-dict genetic structure but not genetic diversity of three frag-mented Afrotropical forest butterfly species.- Conservation
Genetics, 14: 511-528.
BROOK B.W.,SODHI N.S.,BRADSHAW C.J., 2008.- Synergies among extinction drivers under global change.- Trends in
Ecology and Evolution, 23: 453-60.
BUSH J.W.M.,PRAKASH M.,HU D.L., 2007.- The integument of water- walking arthropods: form and function.- Advances
in Insect Physiology, 34: 117-192.
BYERS R.A.,BARKER G.M.,DAVIDSON R.L.,HOEBEKE E.R., SANDERSON M. A., 2000.- Richness and abundance of
Carabidae and Staphylinidae (Coleoptera) in northeastern dairy pastures under intensive grazing.- Great Lakes
Ento-mologist, 33: 81-105.
CABALLERO U.,LEÓN-CORTÉS J.L.,MORÓN-RÍOS A., 2009.- Response of rove beetles (Staphylinidae) to various habitat types and change in Southern Mexico.- Journal of Insect
Conservation, 13: 67-75.
CARDOSO P.,LOBO J.M.,ARANDA S.C.,DINIS F.,GASPAR C., BORGES P.A.V., 2009.- A spatial scale assessment of
habi-tat effects on arthropod communities of an oceanic island.-
CARDOSO P.,BORGES P.A.V.,DINIS F., GASPAR C., 2010.- Patterns of alpha and beta diversity of epigean arthropods at contrasting land-uses of an oceanic island (Terceira, Azores), pp.73-88. In: Terrestrial arthropods of
Macarone-sia - biodiversity, ecology and evolution (SERRANO A.R.M.,
BORGES P.A.V.,BOIEIRO M.,OROMÍ P., Eds).- Sociedade
Portuguesa de Entomologia, Lisboa, Portugal.
CARINE M., SCHAEFER H., 2010.- The Azorean diversity
enigma: why are there so few Azorean endemic flowering plants and why are they so widespread?- Journal of
Bio-geography, 37: 77-89.
CARLSON C.S.,THOMAS D.J.,EBERLE M.A.,SWANSON J.E.,
LIVINGSTON R.J.,RIEDER M.J., 2005.- Genomic regions
ex-hibiting positive selection identified from dense genotype data.- Genome Research, 15: 1553-1565.
CATERINO M.S.,HUNT T.,VOGLER A.P., 2005.- On the
consti-tution and phylogeny of Staphyliniformia.- Molecular
Phy-logenetics and Evolution, 34: 655-672.
ČERNÁ K., STRAKA J., MUNCLINGER P., 2013.- Population
structure of pioneer specialist solitary bee Andrena vaga (Hymenoptera: Andrenidae) in central Europe: the effect of habitat fragmentation or evolutionary history?.-
Conserva-tion Genetics, 14: 875-883.
CHATZIMANOLIS S., COHEN I. M.,SCHOMANN A., S OLODOV-NIKOV A., 2010.- Molecular phylogeny of the mega-diverse
rove beetle tribe Staphylinini (Insecta, Coleoptera, Staphylinidae).- Zoologica Scripta, 39: 436-449.
CLARY D.O.,WOLSTENHOLME D. R., 1985.- The mitochon-drial DNA molecule of Drosophila yakuba: nucleotide se-quence, gene organization, and genetic code.- Journal of
Molecular Evolution, 22: 252-271.
CLEMENT M.,POSADA D., CRANDALL K.A., 2000.- TCS: a
computer program to estimate gene genealogies.- Molecular
Ecology, 9: 1657-1660.
COLWELL R. K., 2011.- EstimateS, Version 8.2: statistical es-timation of species richness and shared species from sam-ples.- Software and user's guide.
DAYRAT B., CONRAD M., BALAYAN S., WHITE T. R.,
ALBRECHT C.,GOLDING R.,GOMES S.R.,HARASEWYCH M. G.,MARTINS A.M.F.,2011.- Phylogenetic relationships and
evolution of pulmonate gastropods (Mollusca): new insights from increased taxon sampling.- Molecular Phylogenetics
and Evolution, 59: 425-437.
DENNIS P.,YOUNG M.R.,HOWARD C.L.,GORDON I.J., 1997.-
The response of epigeal beetles (Col.: Carabidae, Staphylini-dae) to varied grazing regimes on upland Nardus stricta grasslands.- Journal of Applied Ecology, 34: 433-443. DIAS E.,ELIAS R.B.,MELO C.,MENDES C., 2007.- Biology
and ecology of the forests of the islands. Azores. Trees and forests of Portugal, Azores and Madeira, pp. 51-80. In: A
floresta das ilhas (SILVA L., Ed.).- Público, Comunicação
Social, S.A. and Fundação Luso-Americana, Lisbon, Portu-gal. (In Portuguese).
DIXO M.,METZGER J.P.,MORGANTE J.S., ZAMUDIO K.R., 2009.- Habitat fragmentation reduces genetic diversity and connectivity among toad populations in the Brazilian Atlan-tic Coastal Forest.- Biological Conservation, 142: 1560-1569.
ELVEN H.,BACHMANN L.,GUSAROV V.I., 2010.- Phylogeny of
the tribe Athetini (Coleoptera: Staphylinidae) inferred from mitochondrial and nuclear sequence data.- Molecular
Phy-logenetics and Evolution, 57: 84-100.
ELVEN H.,BACHMANN L.,GUSAROV V.I., 2012.- Molecular
phylogeny of the Athetini-Lomechusini-Ecitocharini clade of Aleocharine rove beetles (Insecta).- Zoologica Scripta, 41: 617-636.
FINN D.S.,BLOUIN M.S.,LYTLE D.A., 2007.- Population
ge-netic structure reveals terrestrial affinities for a headwater stream insect.- Freshwater Biology, 52: 1881-1897.
GILLESPIE R.G., RODERICK G.K., 2002.- Arthropods on is-lands: colonization, speciation, and conservation.- Annual
Review of Entomology, 47: 595-632.
GRIMALDI D.,ENGEL M.S., 2005.- Evolution of the insects.-
Cambridge University Press, Cambridge, UK.
GUINDON S.,GASCUEL O., 2003.- A simple, fast, and accurate
algorith to estimate large phylogenies by maximum likeli-hood.- Systematic Biology, 52: 696-704.
GUTIÉRREZ-CHACÓN C.,ZÚÑIGA M.C.,VAN BODEGOM P.M.,
CHARÁ J., GIRALDO L., 2009.- Rove beetles (Coleoptera:
Staphylinidae) in Neotropical riverine landscapes: character-ising their distribution.- Insect Conservation and Diversity, 2: 106-115.
HANSEN M., 1997.- Phylogeny and classification of the staphyliniform beetle families. (Coleoptera).- Biologiske
Skrifter 48, Kongelige Danske Videnskabernes Selskab, Co-penhagen, Denmark.
HEIDINGER I.M. M.,HEIN S., FELDHAAR H., POETHKE H.-J., 2013.- The genetic structure of populations of Metrioptera
bicolor in a spatially structured landscape: effects of
disper-sal barriers and geographic distance.- Conservation
Genet-ics, 14: 299-311.
HILLIS D.M.,DIXON M.T., 1991.- Ribosomal DNA:
molecu-lar evolution and phylogenetic inference.- The Quaterly
Re-view of Biology, 66: 411-453.
HODKINSON I.D.,JACKSON J.K.,2005.- Terrestrial and aquatic invertebrates as bioindicators for environmental monitoring, with particular preference to mountain ecosystems.-
Envi-ronmental Management, 35: 649-666.
HOLLAND B.S.,HADFIELD M.G., 2002.- Islands within an
is-land: phylogeography and conservation genetics of the en-dangered Hawaiian tree snail Achatinella mustelina.-
Mo-lecular Ecology, 11: 365-375.
HORTAL J.,BORGES P.A.V.,GASPAR C., 2006.- Evaluating the
performance of species richness estimators: Sensitivity to sample grain size.- Journal of Animal Ecology, 75: 274-287. HOWLAND D.E.,HEWITT G.M., 1995.- Phylogeny of the Col-eoptera based on mitochondrial cytochrome oxidase I se-quence data.- Insect Molecular Biology, 3: 203-215. JEFFROY O.,BRINKMANN H.,DELSUC F.,PHILIPPE H., 2006.-
Phylogenomics: the beginning of incongruence?.- Trends in
Genetics, 22: 225-231.
JORDAENS K., VAN RIEL P.,MARTINS A.M.F.,BACKELJAU T., 2009.- Speciation on the Azores islands: congruent patterns in shell morphology, genital anatomy, and molecular mark-ers in endemic land snails (Gastropoda, Leptaxinae).-
Bio-logical Journal of the Linnean Society, 97: 166-176.
KLIMASZEWSKI J., WINCHESTER N. N., BOLTE K. B., 2002.-
Aleocharine Rove Beetles (Coleoptera, Staphylinidae) of the Ancient Sitka Spruce Forest on Vancouver Island, British Columbia, Canada.- Canadian Entomologist, 135: 867-868. KROOSS S.,SCHAEFER M., 1998.- The effect of different
farm-ing systems on epigeic arthropods: a five-year study on the rove beetle fauna (Coleoptera: Staphylinidae) of winter wheat.- Agriculture, Ecosystems and Environment, 69: 121-133.
LALIBERTÉ E., WELLS J.A., DECLERCK F., METCALFE D. J.,
CATTERALL C.P.,QUEIROZ C.,AUBIN I.,BONSER S.P.,DING
Y.,FRATERRIGO J.M.,MCNAMARA S.,MORGAN J.W.,M ER-LOS D.S.,VESK P.A.,MAYFIELD M.M., 2010.- Land-use in-tensification reduces functional redundancy and response di-versity in plant communities.- Ecological Letters, 13: 76-86. LAWRENCE J.F.,NEWTON A.F., 1995.- Families and
subfami-lies of Coleoptera (with selected genera, notes, references and data on family-group names), pp 779-1006. In: Biology,
phylogeny and classification of Coleoptera: Papers
cele-brating the 80th birthday of Roy A. Crowson (PAKALUK J.,
SLIPINSKI S. A., Eds).- Muzeum i Instytut Zoologii PAN,
LIBRADO P.,ROZAS J., 2009.- DnaSP v. 5: a software for com-prehensive analysis of DNA polymorphism data.-
Bioinfor-matics, 25: 1451-1452.
LOUY D.,HABEL J.C.,SCHMITT T.,ASSMANN T.,MEYER M.,
MULLER P., 2007.- Strongly diverging population genetic patterns of three skipper species: the role of habitat fragmen-tation and dispersal ability.- Conservation Genetics, 8: 671-681.
MANEL S.,JOOST S.,EPPERSON B.K.,HOLDEREGGER R.,S TOR-FER A.,ROSENBERG M.S.,SCRIBNER K.T.,BONIN A.,FORTIN
M.J., 2010.- Perspectives on the use of landscape genetics
to detect genetic adaptive variation in the field.- Molecular
Ecology, 19: 3760-3772.
MARCELINO J. A. P. , SILVA L., GARCIA P. V., WEBER E.,
SOARES A.O., 2013.- Using species spectra to evaluate plant
community conservation value along a gradient of anthro-pogenic disturbance.- Environmental Monitoring and
As-sessment, 185: 6221-6233.
MARTÍN J.L.,BORGES P.A.V.,ARECHAVALETA M.,FARIA B.,
2008.- The top 100 list, pp. 421-449. In: The 100 threatened
species prioritary for management in Macaronesia (MARTÍN
J.L.,ARECHAVALETA M.,BORGES P.A.V.,FARIA B., Eds).- Consejería de Medio Ambiente y Ordenación Territorial, Canary Islands, Spain.
MARTINS A., SIMÕES N., 1985.- Population dynamics of the
Japanese beetle (Coleoptera - Scarabaeidae) in Terceira is-land - Azores.- Arquipélago, 6: 57-62.
MARTINS J.M.,MOREIRA O.C.B.,RAINHA N.F.P.,BAPTISTA
J.A.B.,SILVA L.,MOURA M.M.T., 2013.- Population
ge-netics and conservation of the Azorean tree Picconia
azo-rica.- Biochemical Systematics and Ecology, 49: 135-143.
MAUS C.,PESCHKE K., DOBLER S., 2001.- Phylogeny of the
genus Aleochara inferred from mitochondrial cytochrome oxidase sequences (Coleoptera: Staphylinidae).- Molecular
Phylogenetics and Evolution, 18: 202-216.
MCKENNA D.D.,FARRELL B.D.,CATERINO M.S.,FARNUM C.
W.,HAWKS D.C.,MADDISON D.R.,SEAGO A.E.,SHORT A. E.Z.,NEWTON A.F.,THAYER M.K.,2015.- Phylogeny and
evolution of the Staphyliniformia and Scarabaeiformia: For-est litter as a stepping-stone for diversification of non-phytophagous beetles.- Systematic Entomology, 40: 35-60. MEIJER S.S.,WHITTAKER R.J.,BORGES P.A.V., 2011.- The
effects of land-use change on arthropod richness and abun-dance on Santa Maria Island (Azores): unmanaged planta-tions favour endemic beetles.- Journal of Insect
Conserva-tion, 15: 505-522.
MYERS N., MITTERMEIER R. A., MITTERMEIER C. G., DA
FONSECA G.A.B.,KENT J.,2000.- Biodiversity hotspots for
conservation priorities.- Nature, 403: 853-858.
OSSWALD J.,BACHMANN L.,GUSAROV V.I., 2013.- Molecular
phylogeny of the beetle tribe Oxypodini (Coleoptera: Staphylinidae: Aleocharinae).- Systematic Entomology, 38: 507-522.
POHL G.R.,LANGOR D.W.,SPENCE J.R., 2007.- Rove beetles and ground beetles (Coleoptera: Staphylinidae, Carabidae) as indicators of harvest and regeneration practices in western Canadian foothills forests.- Biological Conservation, 137: 294-307.
POSADA D., 2008.- jModelTest: phylogenetic model
averag-ing.- Molecular Biology and Evolution, 25: 1253-1256. PREVEDELLO J.A.,VIEIRA M.V., 2010.- Does the type of
ma-trix matter? A quantitative review of the evidence.-
Biodi-versity and Conservation, 19: 1205-1223.
RAINIO J., NIEMELA J., 2003.- Ground beetles (Coleoptera: Carabidae) as bioindicators.- Biodiversity and Conservation, 12: 487-506.
RAMBAUT A., 2009.- FigTree.- version 1.3.1 [online] URL:
http://tree.bio.ed.ac.uk/software/figtree/ (Accessed 16 No-vember 2014).
RICHMOND J.Q.,REID D.T.,ASHTON K.G.,ZAMUDIO K.R., 2009.- Delayed genetic effects of habitat fragmentation on the ecologically specialized Florida Sand Skink (Plestiodon reynoldsi).- Conservation Genetics, 10: 1281-1297.
RODRIGUES P.,LOPES R.J.,DROVETSKI S.V.,REIS S.,RAMOS J. A.,CUNHA R.T., 2013.- Phylogeography and genetic
diver-sity of the robin (Erithacus rubecula) in the Azores islands: evidence of a recent colonisation.- Journal of Ornithology, 154: 889-900.
ROSENBERG M.S.,ANDERSON C.D., 2011.- PASSaGE: Pattern
analysis, spatial statistics, and geographic exegesis, version 2.- Methods in Ecology and Evolution, 2: 229-232.
RUMEU B.,CAUJAPÉ-CASTELLS J.,BLANCO-PASTOR J.L.,JAÉN
-MOLINA R.,NOGALES M.,ELIAS R. B.,VARGAS P., 2011.-
The colonization history of Juniperus brevifolia (Cupres-saceae) in the Azores Islands.- PLoS ONE, 6: e27697. SCHAEFER H.,MOURA M.,MACIEL M.G.B.,SILVA L.,RUMSEY
F.J.,CARINE M., 2011a.- The Linnean shortfall in oceanic island biogeography: a case study in the Azores.- Journal of
Biogeography, 38: 1345-1355.
SCHAEFER H., HARDY O. J., SILVA L., BARRACLOUG T. G.,
SAVOLAINEN V., 2011b.- Testing Darwin´s naturalization hypothesis in the Azores.- Ecological Letters, 14: 389-396. SEGELBACHER G., CUSHMAN S.A., EPPERSON B. K., FORTIN
M.-J.,FRANCOIS O.,HARDY O.J.,HOLDEREGGER R.,MANEL
S., 2010.- Applications of landscape genetics in conserva-tion biology: concepts and challenges.- Conservaconserva-tion
Genet-ics, 11: 375-385.
SILVA L., ELIAS R.B., MOURA M.,MEIMBERG H.,DIAS E.,
2011.- Genetic variability and differentiation among popula-tions of the Azorean endemic gymnosperm Juniperus
brevi-folia: baseline information for a conservation and restoration
perspective.- Biochemical Genetics, 49: 715-734.
SIMON C.,FRATI F.,BECKENBACH A.,CRESPI B.,LIU H.,FLOOK
P.,1994.- Evolution, weighting, and phylogenetic utility of
mitochondrial gene sequences and a compilation of con-served polymerase chain reaction primers.- Annals of the
Entomological Society of America, 87: 651-701.
SLATKIN M., 1993.- Isolation by distance in equilibrium and non-equilibrium populations.- Evolution, 47: 264-279. SOMMER S.,MCDEVITT A.D.,BALKENHOL N., 2013.-
Land-scape genetic approaches in conservation biology and man-agement.- Conservation Genetics, 14: 249-251.
SUMNER J.,JESSOP T.,PAETKAU D.,MORITZ C., 2004.- Limited
effect of anthropogenic habitat fragmentation on molecular diversity in a rain forest skink, Gnypetoscincus
queenslan-diae.- Molecular Ecology, 13: 254-269.
SWOFFORD D.L., 2003.- PAUP*. Phylogenetic analysis using parsimony (*and other methods) version 4.- Sinauer
Associ-ates, Sunderland, Massachusets, USA.
TAJIMA F., 1989.- The effect of change in population size on
DNA polymorphism.- Genetics, 123: 597-601.
TAMURA K.,PETERSON D.,PETERSON N.,STECHER G.,NEI M.,
KUMAR S., 2011.- MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods.- Molecular Biology and
Evolution, 28: 2731-2739.
TAUTZ D.,HANCOCK J.M.,WEBB D.A.,TAUTZ C.,DOVER G.A., 1988.- Complete sequences of the rRNA genes of Drosophila
melanogaster.- Molecular Biology and Evolution, 5: 366-376.
THAYER M.K., 2005.- Staphylinoidea, Staphylinidae Latreille,
1802. Coleoptera, Vol I. Morphology and Systematics (Ar-chostemata, Adephaga, Myxophaga, Polyphaga partim), pp. 296-344. In: Handbook of zoology, arthropoda: insecta, (BEUTEL R.G.,LESCHEN R.A.B.,KRISTENSEN N. P., Eds).-
De Gruyter, Berlin, Germany.
THÉRY T.,LESCHEN R.A.B., 2013.- Pselaphinae (Coleoptera:
Staphylinidae) of the Three Kings Islands.- New Zealand
THOMAS J.C., 2009.- A preliminary molecular investigation of Aleocharine phylogeny (Coleoptera: Staphylinidae).- Annals
of the Entomological Society of America, 102: 189-195.
TRIANTIS K.A.,HORTAL J.,AMORIM I.,CARDOSO P.,SANTOS
A.M.C.,GABRIEL R.,BORGES P.A.V., 2012.- Resolving the Azorean knot: a response to Carine and Schaefer 2010.-
Journal of Biogeography, 39: 1179-1184.
VOGLER A.P.,PEARSON D.L., 1996.- A molecular phylogeny
of the tiger beetles: congruence of mitochondrial and nuclear rDNA data sets.- Molecular Phylogenetics and Evolution, 6: 321-338.
WESTERN D., 2001.- Human-modified ecosystems and future
evolution.- Proceedings of the National Academy of
Sci-ences of the United States of America, 98: 5458-5465.
WHITTAKER R.J.,FERNÁNDEZ-PALACIOS J.M., 2007.- Island biogeography - ecology, evolution and conservation.-
Ox-ford University Press, OxOx-ford, UK.
Authors’ addresses: José A. P. MARCELINO (corresponding
author, [email protected]), Paulo A. V. BORGES, Patricia V.
GARCIA, António O. SOARES, CE3C, Azorean Biodiversity
Group, Department of Biology, University of the Azores, rua da Mae de Deus 13-A, 9501-801 Ponta Delgada, Azores,
Por-tugal; Rosanna GIORDANO, Department of Biology, PO Box
23360, University of Puerto Rico, San Juan, PR 00931, USA;
Felipe N. SOTO-ADAMES, Department of Entomology,
Univer-sity of Illinois, Urbana, IL, USA